
Abstract
Research in attention and action control produced substantial evidence suggesting the presence of feature binding. This study explores the binding of task-irrelevant context features in cued task switching. We predicted that repeating a context feature in trial n retrieves the trial n − 1 episode. Consequently, performance should improve when the retrieved features match the features of the current trial. Two experiments (N = 124; N = 96) employing different tasks and materials showed that repeating the task-irrelevant context improved performance when the task and the response repeated. Furthermore, repeating the task-irrelevant context increased task repetition benefits only when the context feature appeared synchronously with cue onset, but not when the context feature appeared with a 300-ms delay (Experiment 1). Similarly, repeating the task-irrelevant context improved performance when the task and the response repeated only when the context feature was part of the cue, and not when it was part of the target (Experiment 2). Taken together, binding and retrieval processes seem to play a crucial role in task switching, alongside response inhibition processes. In turn, our study provided a better understanding of binding and retrieval of task-irrelevant features in general, and specifically on how they modulate response repetition benefits in task repetitions.
Humans need to effectively interact with a continuous flow of stimuli by responding with appropriate actions given their goals. This requires focusing the attention on the currently relevant stimulus and selecting the appropriate response. Attention switching and action selection involve cognitive control, although automatic processes are also involved (Koch, Poljac, et al., 2018). In the present study, we investigated feature binding in cued task-switching. Specifically, we examined whether a task-irrelevant visual feature that repeats in the subsequent trial can reactivate (i.e., retrieve) the task and the response of the previous trial. Furthermore, we studied the timing of such binding and retrieval processes by manipulating the asynchrony between the beginning of a trial and the onset of the task-irrelevant feature.
In task switching, participants work through a (usually unpredictable) sequence of tasks, in which two or more tasks alternate (for reviews, see Kiesel et al., 2010; Koch, Poljac, et al., 2018; Monsell, 2003; Vandierendonck et al., 2010). When the task in the current trial is the same as in the previous trial, participants are generally faster and more accurate than when the task switches from the previous trial (i.e., task repetition benefits). Task switching was extensively used to study cognitive control mechanisms underlying attention-switching and flexible action control (e.g., inhibitory control; see Kiesel et al., 2010). However, other processes are also at play in task switching, namely, bottom-up, stimuli-based (re-)activation (retrieval or priming).
The latter processes emerge when a stimulus retrieves a previously formed association: a cue may automatically retrieve the stimulus-response mapping it was associated with; a target stimulus may retrieve a previously given response. A great number of studies uncovered diverse retrieval processes such as the exogenous cueing of task-set (Rogers & Monsell, 1995), the priming of stimulus-task association (Koch & Allport, 2006; Waszak et al., 2003), or the nonmediated route of response selection (e.g., Frings et al., 2015; Li et al., 2019; Schneider & Logan, 2014). However, delving into each of these processes is beyond the scope of this study. Central for the present study is the assumption they underlie. In fact, for these processes to take place, stimulus features must become associated with the responses and, when they repeat, stimulus features must be able to reactivate such associations, impacting performance.
The response-repetition effect in task switching
The response-repetition (RR) effect is a robust finding in the task-switching literature (Altmann, 2011; Druey, 2014; Grzyb & Hübner, 2013; Kiesel et al., 2010; Kleinsorge, 1999; Koch et al., 2011; Rogers & Monsell, 1995; Schuch & Koch, 2004, 2010). Repeating a response usually yields a benefit in task repetitions, which disappears or becomes a cost in task switches. The RR effect is indicated statistically by an interaction between task relation (repetition vs. switch) and response relation (repetition vs. switch).
Different accounts propose explanations for the RR effect, which can be attributed to two families: accounts assuming episodic-retrieval or association-learning mechanisms and response-inhibition accounts. According to the latter accounts, when participants have to alternate between different tasks and responses, responses are inhibited right after execution, possibly to avoid unintentional re-execution (i.e., perseveration) of the response in the next trial (see Druey, 2014; Hübner & Druey, 2006; Koch et al., 2011; Rogers & Monsell, 1995). However, the task-relevant stimulus categories that are mapped to a certain response can prime the response. As a consequence, when the task-relevant stimulus categories repeat (i.e., in task repetitions), the previously inhibited response is primed, counteracting the inhibition. In task switches, the task-relevant stimulus categories switch as well, so that they cannot prime the response. Thus, an RR cost is observed due to inhibition of the response (Hübner & Druey, 2006; Koch et al., 2011).
According to the episodic-retrieval or association-learning accounts (Altmann, 2011; Frings et al., 2020; Schuch & Koch, 2004), RR benefits in task repetitions originate from the strengthening of the association between the task-relevant stimulus category and the response. When the task repeats in the subsequent trial, it retrieves the associated response: this generates a benefit when the correct response coincides with the retrieved response, compared with when it does not coincide. On the contrary, a task switch in combination with a response repetition yields RR costs due to unbinding the response from the task, since the response was associated with the other task in the previous trial (Schuch & Koch, 2004).
The Binding and Retrieval in Action Control (BRAC) framework (Frings et al., 2020) incorporates notions from episodic retrieval theories, such as the Theory of Event Coding (TEC; Hommel, 1998, 2019) and the Instance Theory of Automatization (ITA; Logan, 1988). BRAC offers a parsimonious framework to explain the RR effect by postulating that the features of a trial are bound together into a coherent representation that can be retrieved from memory when a feature repeats. Therefore, whenever any of the retrieved features mismatch with the features of the current trial, this results in a performance cost (i.e., partial repetition cost). This pattern results in a significant interaction of switching versus repeating two or more features, such as the task and the response.
Binding of context in task switching
Koch, Frings, and Schuch (2018), Kandalowski et al. (2020) and Benini et al. (2022) set out to examine the binding of the task and the response with a task irrelevant feature (the context henceforth). The context is a further dimension of our stimuli and differs from a distractor, which is usually mapped to a response and might be task-relevant in other trials (cf. Moeller & Frings, 2014, p. 976). In Koch, Poljac, et al. (2018), Kandalowski et al. (2020), and Benini et al. (2022; Experiment 1), the context was the cue modality (auditory vs. visual), which was neither informative about the current task nor the current correct response. Furthermore, in Kandalowski et al. (2020), also target modality was manipulated on a trial-to-trial basis so that the target digits could be presented written on the screen or as a spoken number. In Benini et al. (2022; Experiment 2) instead, the context was the language of the cue.
Concerning task repetition performance, Koch, Frings, and Schuch (2018) and Benini et al. (2022; Experiment 2) found RR benefits that were reduced when cue modality switched than when it repeated (the same pattern was observed by Kandalowski et al., 2020 and Benini et al., 2022 [Experiment 1], albeit not significant). This suggests that the repeating context might have retrieved the features from the previous trial, thereby implying a benefit when also the task and the response repeated. Instead, the switching context mismatched with the repeating task and response, and, therefore, the RR benefit was strongly reduced.
Concerning performance in task switches, the three studies found RR costs, but no effect of cue modality relation on such RR costs. However, the notion of partial repetition costs (Frings et al., 2020) would predict for the task-switch/response-switch/context-switch condition to be faster and more accurate than any partial repetition condition. Thus, the observed data pattern was not completely in line with episodic-retrieval accounts. As we described above, the response-inhibition account predicts RR costs in task switches. However, this account is silent concerning the effect of an additional context. Since neither account can explain the whole data pattern, Koch, Frings, and Schuch (2018) suggested a hybrid account stating that binding and retrieval processes are likely to coexist with response inhibition processes in task switching.
The present study
In the present study, we introduced a visual context feature (i.e., the colour of the cue) to further explore the influence of a task-irrelevant context on binding and retrieval effects in task switching. For each task, the cue was one rectangle which could be either in blue or red, depending on the level of the context. Using the colour of the cue as the context allowed to increase the similarity of the task cue between the two different context levels (i.e., only the colour changed). In contrast, in Koch, Frings, and Schuch (2018) and Kandalowski et al. (2020), the cue of one task was quite different in the two contexts (e.g., a bar in the top half of the screen vs. a high-pitched tone). Our context operationalization was therefore an irrelevant dimension of the cue, which could be added or removed (it would have been instead impossible to remove the cue modality). Indeed, importantly, we also manipulated the onset of the context such that it may appear at three different times within a trial: at cue onset, 300 ms after cue onset, or at target onset. The effects of asynchrony between the target and other features were already investigated using paradigms different from task switching (e.g., Frings et al., 2017; Frings & Moeller, 2012; Maetens et al., 2009). To our knowledge, Kandalowski et al. (2020) were the only ones to examine cue-onset versus target-onset of the context, but they did not find an effect of the target-related context on the RR effect. In fact, they also did not replicate the effect of a cue-related context. Therefore, more studies are needed to draw inferences about the timing of context binding processes in task switching.
In our experiments, participants switched between two tasks, while the cue appearing before the target stimulus signalled the task to perform in each trial. In Experiment 1, each of the two tasks was cued by a vertical or a horizontal rectangle respectively, and the context was a task-irrelevant cue–colour dimension so that the rectangle could be red or blue. The cue colour did not provide any information functional for task identification and response selection. Importantly, in half of the blocks, cue–context onset asynchrony was 0 ms so that the cue appeared immediately coloured, either in blue or in red. In the other half of the blocks, cue–context onset asynchrony was 300 ms so that the cue appeared in black for the first 300 ms, before it became either blue or red.
In Experiment 2, the two cues were words semantically related to the tasks, and the context was the colour of the words’ ink. Furthermore, it was manipulated between participants whether the task-irrelevant context appeared simultaneously with the cue (i.e., it was the colour of cue-word) or the target (i.e., it was the colour of target word).
In these experiments, we expected RR benefits in task repetitions and RR costs in task switches. Based on previous studies (Benini et al., 2022; Frings et al., 2017; Kandalowski et al., 2020; Koch, Frings, & Schuch, 2018), we further expected the context to be bound with the task and the response, and to retrieve them when it repeats in the subsequent trial. Statistically, this should result in a three-way interaction of context relation with task relation and response relation.
Importantly, we expected such modulation to be weakened by a delayed context onset for two reasons. The first reason is that when the context appears later in the trial (i.e., 300 ms after cue onset in Experiment 1 or simultaneously to target onset in Experiment 2), also the context-based retrieval happens later. At that point in time, the task is already well prepared, since the cue-based activation of the task had already started, as suggested by a study by Rubin and Koch (2006).
Rubin and Koch (2006) systematically associated an irrelevant feature of the target with one task, thus creating a consistent colour-task association. After the first three blocks, they reversed the colour-task association without any warning. Then, they reversed the association again after three more blocks, and so on. This procedure yielded “invalid” trials, in which the task–colour association was opposite to what was previously learned, thus retrieving the competing task in some trials. In such “invalid” trials, they observed some bottom-up, stimulus-driven task interference, in the form of larger task repetition benefits. However, they found such interference only when the current task was not sufficiently prepared (i.e., with a cue–stimulus interval [CSI] of 100 ms in their study). In contrast, repetition benefits were the same in valid and invalid trials when the currently relevant task could be sufficiently prepared (i.e., CSI = 1,100 ms).
Nonetheless, the present study is different from Rubin and Koch’s (2006) for two important reasons: first, Rubin and Koch’s (2006) context was predictive of the current task, while in the present study it was perfectly orthogonal to the task, correct response, and target. Second, they varied the CSI, which was constant in our study, while we varied context onset delay (and, therefore, context presentation time), which was constant in Rubin and Koch (2006). The onset asynchrony of our task-irrelevant feature thus represents a novel manipulation, and it is aimed to shed more light on the retrieval mechanisms in task switching.
The second reason why a delayed context onset should reduce context modulation effects is that, specifically in Experiment 2, when the context appeared at target onset, target encoding, and response selection are triggered at the same time as the context-triggered task retrieval. Therefore, we expect context-based retrieval to be masked by the other ongoing processes (Frings & Moeller, 2012; Maetens et al., 2009) and to exert only small effects (if observable at all).
Experiment 1
We devised a laboratory version and an online version of this experiment. Little difference exists between the two versions so that we could merge their data. The differences are detailed in the following sections.
Materials and methods
Participants
Thirty-three students from RWTH University took part in the laboratory version of the experiment. For the online version, we recruited 13 RWTH students, whereas the rest of the sample was recruited via Prolific (https://www.prolific.co/). In Koch, Frings, and Schuch (2018), the size of the critical interaction effect between task relation, response relation, and context relation was large ( = .25) such that it can be detected with 88.5% power with 32 participants (Faul et al., 2007). Anticipating that data collected online would be noisier than lab data, we planned to recruit 96 participants for the online version (including RWTH students and Prolific participants). Thus, we stopped Prolific data collection when we reached 83 submissions having less than 100 errors (corresponding to an error rate less or equal to 26%). Psychology students between 18 and 35 years old were recruited via email and word of mouth and were compensated with partial course credits. Participants in Prolific could participate if they were between 18 and 35 years old and no other filter was used. They were rewarded with ₤2.75 (₤6.6/h). The online experiment was set up such that it was possible to take part only using a laptop or a computer (as opposed to tablets and smartphones) and using Chrome or Safari as a browser.
The complete sample including laboratory and online participants (N = 129) included 74 females (2 did not declare), the mean age was 23.8 (±3.6) years, and 14 participants were left-handed (one did not declare). 1 The data of participants showing an average error rate above 20% were removed from further analyses (1 lab participant and 4 online participants). Thus, our final sample included 124 participants with an average error rate equal to 7% (±4%).
The procedure of this study, as well as Experiment 2, received approval from the ethics committee of the Faculty of Arts and Humanities at RWTH Aachen University (ethics approval number: 2020_006_FB7_RWTH Aachen).
Neither Experiment 1 nor Experiment 2 was preregistered. All the data (http://doi.org/10.23668/psycharchives.5116) and the code used to run the analyses (https://doi.org/10.23668/psycharchives.6891), including auxiliary analyses, are made available online. The study materials are available on request from the corresponding author.
Stimuli, tasks, and responses
The lab experiment was run using Psychopy v1.84 (Peirce, 2007). Instructions and stimuli were presented in black colour over a grey background. Participants looked at the screen from approximately a 50-cm distance. Target stimuli were digits from 1 to 9, excluding 5. They were approximately 8 mm high and 4 mm wide and were presented in the middle of the computer screen (19 in. screen resolution: 1,280, 1,024 pixels). The cue was a filled rectangle of 3.5 × 1.47 cm size, centred at 7.5 cm above the centre of the screen. There was a black empty frame surrounding both the cue and the stimulus that appeared and disappeared together with the cue in each trial. The frame was a square with a 24 cm side, centrally presented. Participants were required to perform a magnitude task, indicating whether the stimulus was less or greater than five, or a parity task, indicating whether the stimulus was odd or even. The task to perform in each trial was signalled by cue orientation, which could be either horizontal or vertical. Cue–task mapping was counterbalanced across participants. Participants responded using A and L keys on a German QWERTZ keyboard so that response-key mapping overlapped across the two tasks. Response-key mapping was counterbalanced across participants.
The online experiment was hosted and run on Gorilla Experiment Builder (Anwyl-Irvine et al., 2020). Task and stimuli were the same as for the lab experiment, but the cue and the frame were smaller (cue was 2 × 1 cm and frame side was 11.5 cm) to ensure that these were visible on laptops with average to small screen sizes. The digits had the same size as for the lab experiment. The instructions and the digits were black as in the lab version; however, the background was white, and not grey, throughout the whole online experiment.
For both versions of the experiment (i.e., laboratory and online), the cue rectangles could be filled in blue or red, representing the task-irrelevant context. In blocks in which cue–context onset asynchrony was equal to 300 ms, the cue rectangles were presented in black for the first 300 ms.
Procedure
Participants in the lab version were tested individually in sound-proof chambers during sessions that lasted roughly 50 min. Participants provided written informed consent and accepted the data protection regulation, then they read the instructions and the experimenter ensured these were clear before starting the practice block. The experimenter would previously stick an A5 sheet reporting the task–cue and stimulus–response mappings below the computer monitor.
Participants in the online version were sent an email with the link to the experiment (RWTH students), or they accessed the experiment via Prolific. In both cases, they could start when it pleased them and they had a maximum of 1 hr, against an average completion time of 25–30 min. They gave written informed consent and accepted the data protection regulation before they could continue with the experiment. The information concerning the key-response mapping and the cue–task mapping was presented at the bottom of the screen in each training and experimental trial.
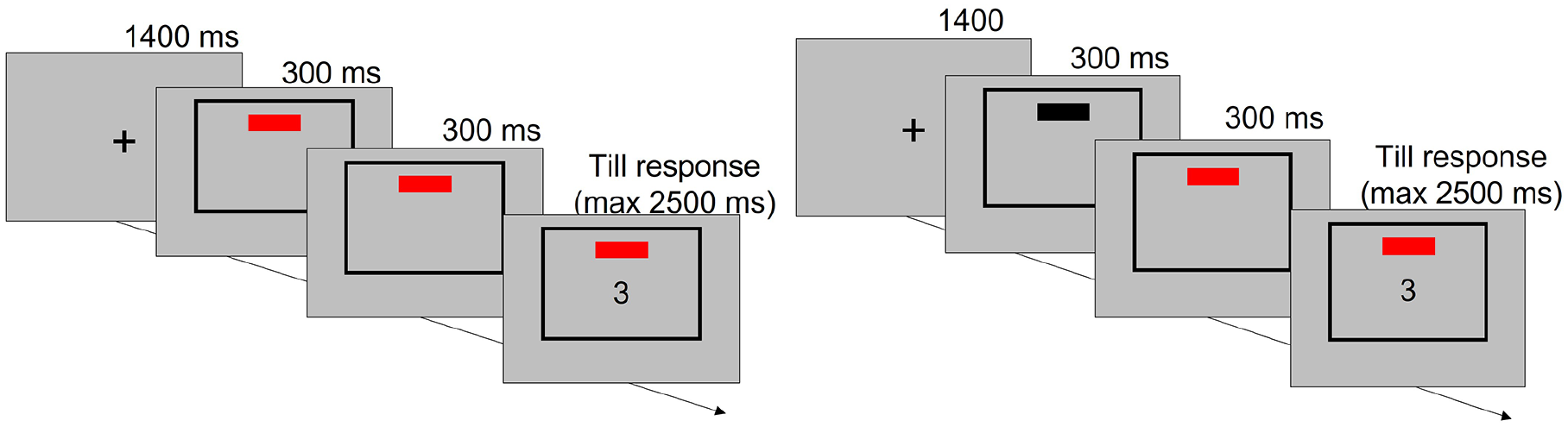
Comparison of the timeline of trials with 0 ms (left) versus 300 ms (right) cue–context onset asynchrony in Experiment 1.
The instructions specified that the cue rectangle would appear in blue or red and that, however, this was not relevant to their task. The instructions were in German in the lab version of the Experiment, but in English in the online version. After the instructions, participants performed 16 practice trials that covered all the possible combinations of stimuli and tasks (eight stimuli × two tasks). In this phase, feedback was given for wrong answers and/or for answers slower than two seconds. It consisted of a sentence saying: “The answer is incorrect! Please try to be more careful!” and/or “Please try to be faster!”, which appeared in black on the screen. In the online version, the accuracy feedback was given through a green tick or a red cross. During training trials, the colour was never added to the cue that was consequently always black.
When laboratory participants declared to the experimenter to be ready, the experiment began, which was composed of eight blocks of 96 trials. Similarly, also online participants could decide to re-read the instructions and do the training again, or they could decide to begin. To comply with general recommendations concerning online studies (Grootswagers, 2020; Sauter et al., 2020), we halved the duration of the online experiment, administering four blocks instead of eight.
Each trial started with a black fixation cross (1,400 ms) followed by the simultaneous appearance of the cue and the frame. Both cue and frame remained on the screen for 600 ms in total, after which the stimulus appeared. The three objects remained on the screen until a response was given or a deadline of 2,500 ms was reached. The following trial started immediately after, with the onset of the fixation cross. No feedback was provided for correct, incorrect, or slow responses in the experimental blocks.
The cue–context onset asynchrony manipulation was implemented block-wise so that it was the same each second block. Thus, four blocks had 300 ms cue–context onset asynchrony and the other four had 0 ms cue–context onset asynchrony (two and two in the online version). It was counterbalanced across participants whether the blocks with 300 ms cue–context onset asynchrony were the even- or the odd-numbered blocks. In each block, half of the trials were magnitude trials and half parity trials, in each task, the cue was blue in half of the trials and red in the other half; and each digit was presented exactly 12 times. Furthermore, the probability of a certain colour given a certain task and a certain stimulus was balanced; the number of task switches was approximately the same as task repetitions trials (in each block there was always exactly one task switch more or one task repetition more) and it was never allowed for the same stimulus (the digit) to repeat in subsequent trials. In the trial sequences that we obtained, each colour was presented almost as often in task switches as in repetitions, and the same holds for response switches and repetitions so that no feature was predictive of the other features in each trial.
Design
The experiment had a 2 × 2 × 2 × 2 within-subjects design. To test our hypotheses, we ran two analyses of variance (ANOVAs) with reaction times (RTs) and error rates as the dependent variables, and task relation (repetition vs. switch), response relation (repetition vs. switch), context relation (repetition vs. switch), and cue–context onset asynchrony (0 vs. 300 ms) as independent variables.
Results
The analyses were conducted in R Studio (R version 4.0.0, R Core Team, 2020), using the afex package (Singmann et al., 2020).
Analyses of RTs
For this ANOVA, error and post-error trials were removed, together with the first trial of each block and the responses faster than 200 ms: this implied removing 13.2% of the initial dataset.
We found a significant main effect of task relation, F(1, 123) = 198.87, p < .001, = .62, indicating task-repetition benefits of 101 ms (751 vs. 852 ms, for task repetitions vs. switches, respectively). We found a significant effect of response relation, F(1, 123) = 11.06, p = .001, = .08, indicating general RR costs (808 vs. 795 ms, for response repetitions vs. switches, respectively). Response relation also entered in a significant interaction with task relation, F(1, 123) = 46.39, p < .001, = .27, indicating RR benefits in task repetitions (10 ms) but costs in task switches (−38 ms).
The main effect of context relation was significant, F(1, 123) = 8.58, p = .004, = .06, indicating general context-repetition benefits of 9 ms (797 vs. 806 ms, for context repetitions vs. switches, respectively). Critically, context relation significantly interacted with task relation, F(1, 123) = 4.14, p = .044, = .03, so that task-repetition benefits were smaller in context switches (96 ms) than in context repetitions (107 ms).
Importantly, this interaction was further qualified by a significant three-way interaction including response relation, F(1, 123) = 4.33, p = .04, = .03. To follow-up on this interaction, we ran separate ANOVAs for task repetitions and task switches. In task repetitions, context relation significantly interacted with response relation, F(1, 123) = 7.81, p = .006, = .06. As shown in Figure 2, RR benefits in task repetitions are larger when context repeated (22 ms) than when context switched (−1 ms). In contrast, in task switches, this interaction was not significant, F(1, 123) = 0.06, p = .801, < .001. That is, RR costs in task switches were almost unaffected by a context repetition versus a switch: we observed a cost of 39 ms in context repetitions and 36 ms in context switches.
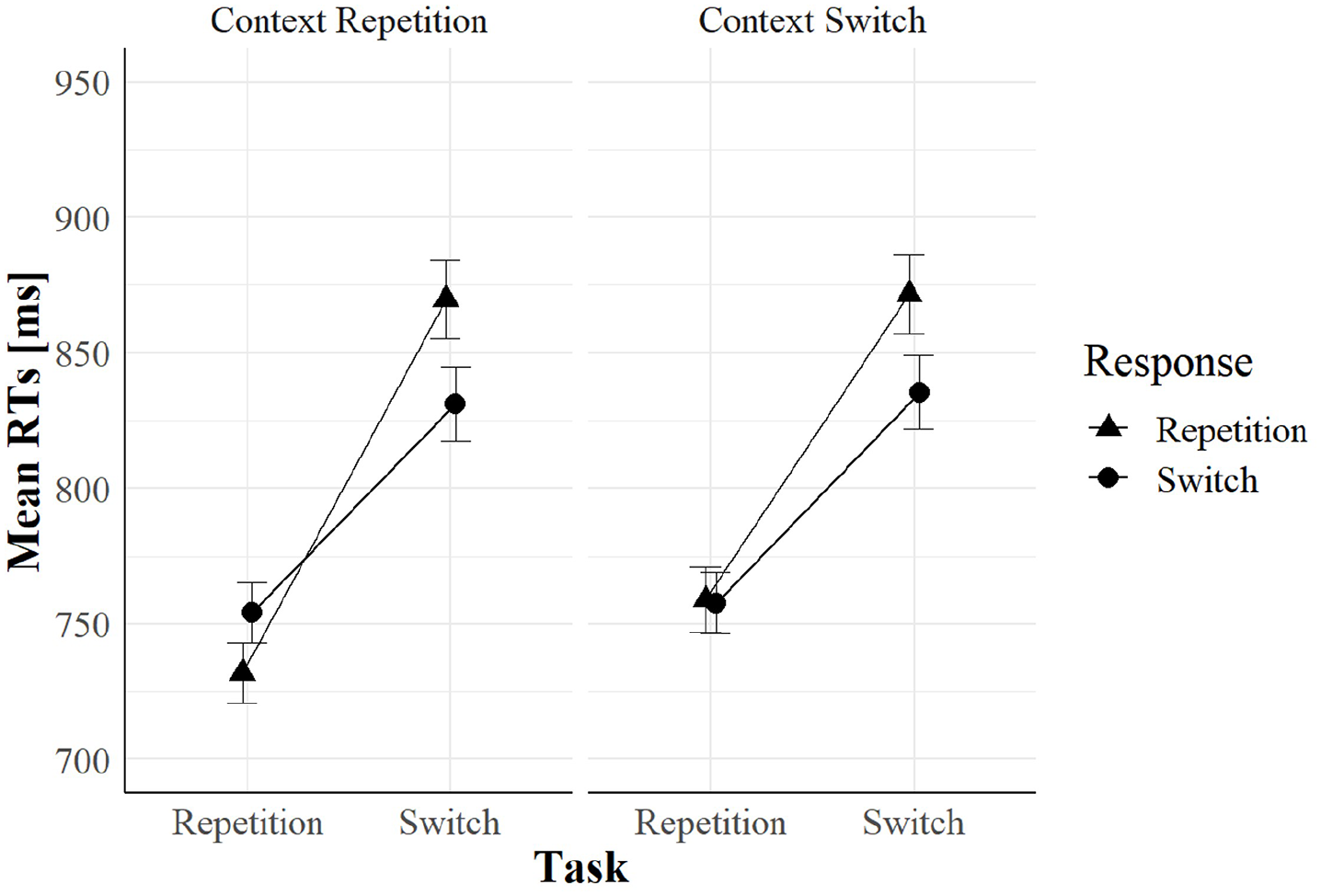
Mean RTs as a function of task relation, response relation, and context relation in Experiment 1.
Task relation entered also another significant three-way interaction with context relation and cue–context onset asynchrony, F(1, 123) = 5.82, p = .017, = .04. We ran two separate ANOVAs on 0 and 300 ms cue–context onset asynchrony trials: context relation significantly interacted with task relation only when cue–context onset asynchrony was 0 ms, F(1, 123) = 8.63, p = .004, = .06, but not when it was 300 ms, F(1, 123) = 0.33, p = .565, < .01. The interaction in the former subset was driven by task-repetition benefits that were smaller in context switches (87 ms) than in context repetitions (116 ms), as shown in Figure 3 (see also Figure 4).
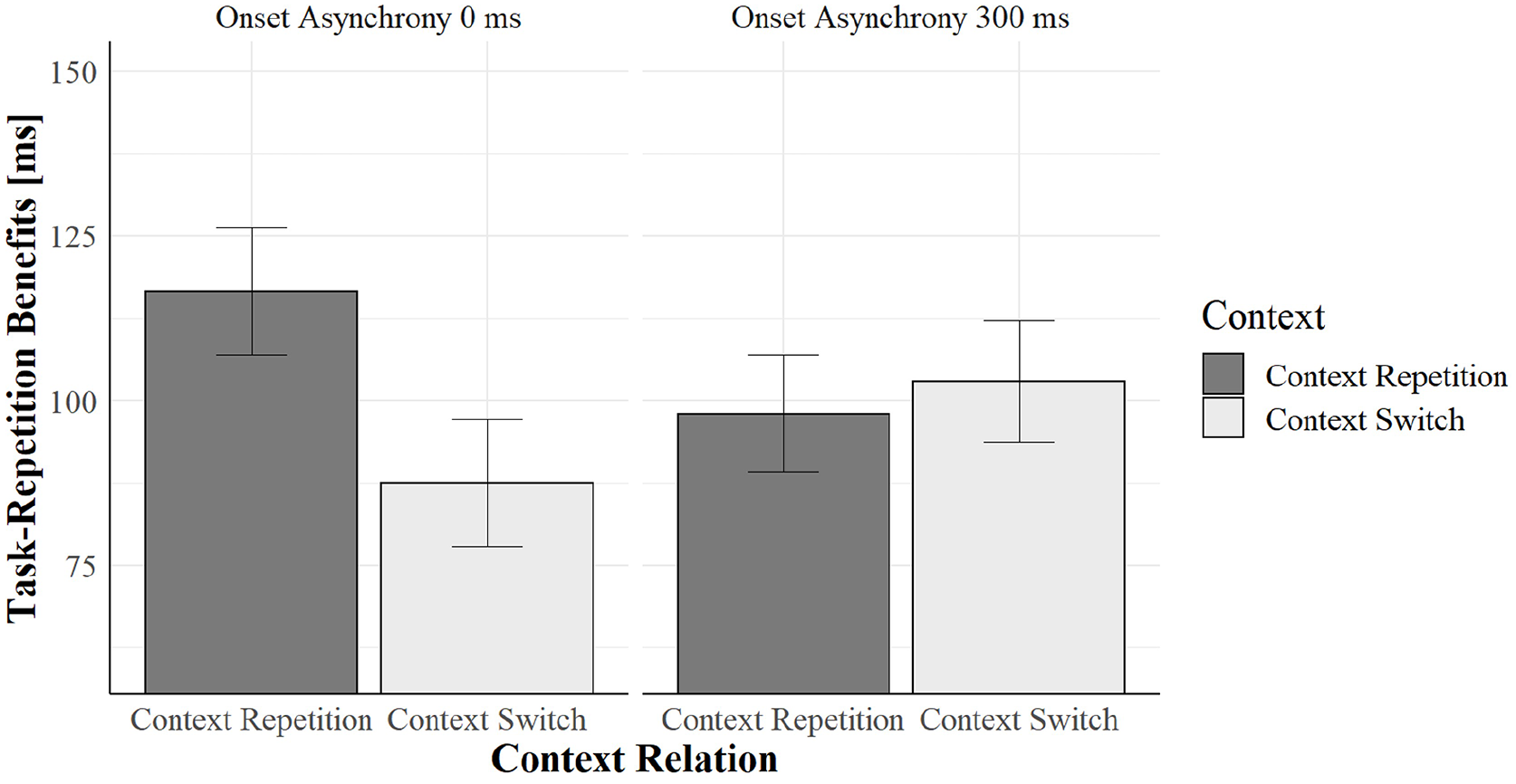
Task-repetition benefits as a function of context relation and onset asynchrony in Experiment 1.
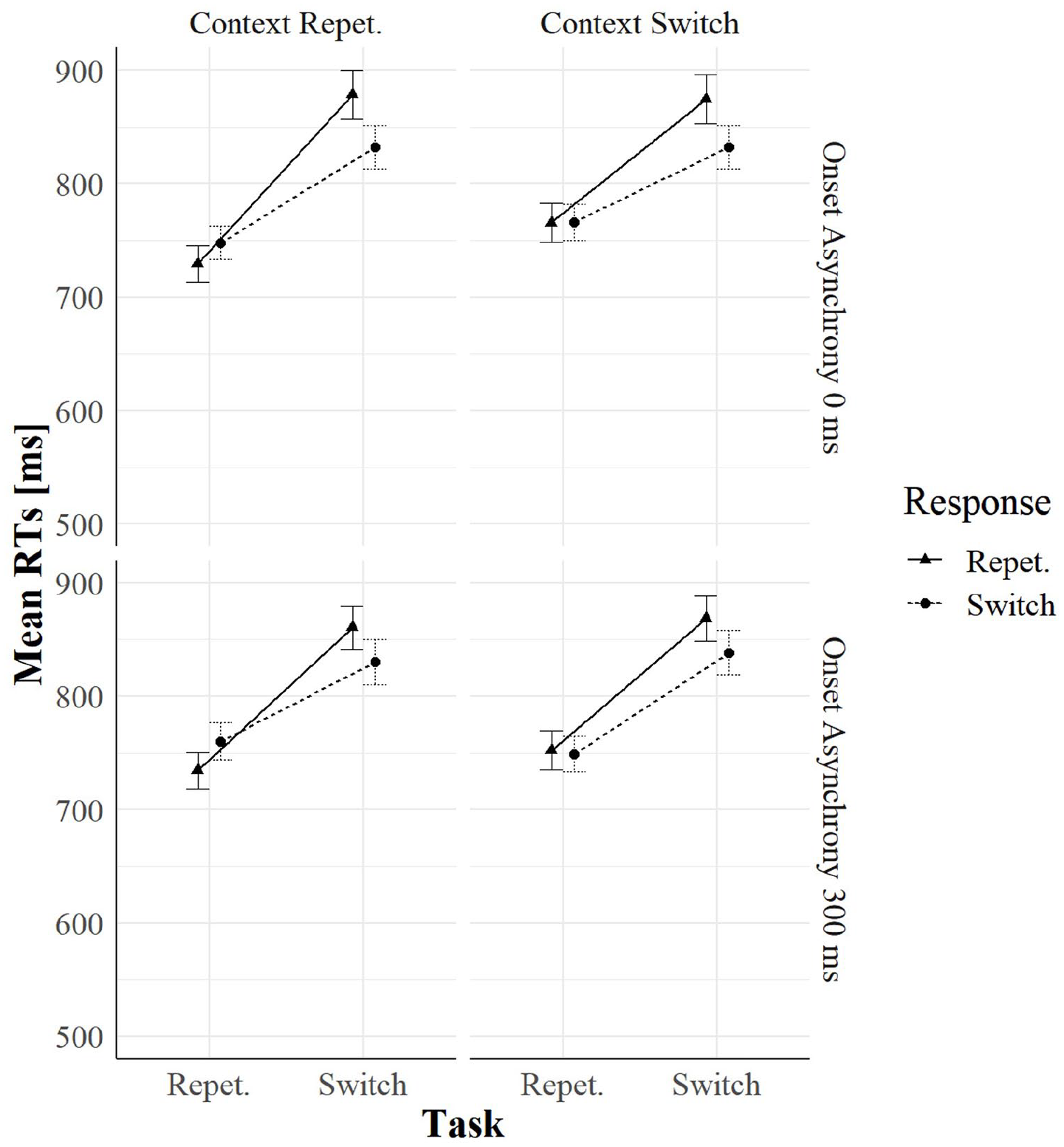
RTs as a function of task relation, response relation, context relation, and onset asynchrony in Experiment 1.
No other main effect or interaction was significant at the 5% alpha level (all Fs < 2.81 and all ps > .096). 2
Analyses of error rates
For this ANOVA, we removed the first trial of each block, the responses faster than 200 ms and post-error trials: this implied removing 7.6% of the initial dataset.
We found a significant main effect of task relation, F(1, 123) = 68.02, p < .001, = .35, indicating task-repetition benefits (5.1% error rate in task repetitions and 7.7% in task switches). We also found a significant main effect of response relation, F(1, 123) = 93.82, p < .001, = .43, indicating RR costs at 7.7% versus 5.2%, for response repetitions versus switches, respectively. Task relation and response relation entered in a significant two-way interaction, F(1, 123) = 35.79, p < .001, = .22, indicating smaller RR costs in task repetition (0.9%) than in task switches (4.1%). Thus, in the ERs we did not find RR benefits in task repetitions, as we found in the RTs.
The main effect of context relation was significant, F(1, 123) = 4.14, p = .044, = .03, indicating small context-repetition costs (6.6% vs. 6.2%, for context repetitions vs. switches, respectively). In the RTs, we found context-repetition benefits instead.
All the other main effects and interactions were not significant (all Fs < 2.93, all ps > .089) that is, context relation did not interact with task relation or response relation as happened in the RTs.
Discussion
In this experiment, the colour of the cue affected performance, although it was a task-irrelevant contextual feature. Specifically, repeating or switching the context had an interactive effect on repeating vs. switching the other, task-relevant variables: task and response. We only observed this effect in participants’ RTs, while context did not interact with any other variable in participants’ error rates. In the error rates, we only found a main effect of context relation, indicating greater error rates in context-repetition compared with context switches. For this reason, we will not further discuss error rates results.
Context relation modulated task repetition benefits, namely context switch trials reduced task repetition benefits. Importantly, such modulation was further affected by the cue–context onset asynchrony. Only when the colour appeared simultaneously with the cue, task-repetition benefits were greater in context repetitions than in context switches. When the colour appeared 300 ms after the cue onset, task-repetition benefits were substantially the same in colour switch and colour repetition trials (see Figure 3).
Furthermore, we observed RR benefits in task repetitions only when the context also repeated; the RR benefit disappeared when the context switched (see Figure 2). If a task-irrelevant context is bound into a representation of the previous trial (an event file), it should retrieve the features of the previous trial only when it repeats in the current trial. Therefore, performance should improve when the task-relevant features repeat as well: this is what we observed in RTs.
In contrast, context relation did not modulate performance in task switches. This result closely replicates the finding of Benini et al. (2022), Kandalowski et al. (2020), and Koch, Frings, and Schuch (2018) who found no effect of context relation in task switches.
In Experiment 2, we sought to extend the current findings by showing the same pattern with different tasks and materials and by modifying the cue–context onset asynchrony manipulation in two ways. First, we increased the asynchrony to 600 ms, so that the delayed context appeared together with the target. Second, the cue–context onset asynchrony manipulation was administered between subjects. Therefore, in the cue-related context group, the context onset coincided with the cue onset, but in the target-related context group, it coincided with the target onset. In the latter case, we explored the effect of further increasing the cue–context onset asynchrony, but in such a way as to increase the relevance of the context: being a feature of the target, it was necessarily encoded.
Experiment 2
In Experiment 2, the cue–context onset asynchrony was either 0 ms (i.e., cue and context onset coincided) or 600 ms (i.e., target and context onset coincided). Apart from this, Experiment 2 differed from Experiment 1 in the set of tasks (discriminate objects based on their human-made vs. natural origin or based on their size), stimuli (words), and procedure (the cue disappeared when the target appeared). We additionally included single-task blocks to investigate whether context relation interacted with response relation when the situation did not require task switching. However, we will focus on the results of the mixed-task blocks, as these data can be compared with those from Experiment 1. As in Experiment 1, the context was the irrelevant colour of the stimuli. Using words as cues and targets allows to have the same operationalization of the context for the cues and the targets, that is, the words’ ink colour.
In Experiment 1, context-binding effects were largely absent when the context appeared only after the cue. In Experiment 2, we were specifically interested to see if such effects would be present if the context is a feature of the target and thus it needs to be encoded. In Experiment 2, for the cue-related context, we expected the same pattern of results as in Experiment 1, which will demonstrate that context binding effects extend beyond a specific experimental set-up.
Materials and methods
Participants
Participants were recruited via Prolific (https://www.prolific.co/) and among psychology students at RWTH University. We did not devise a lab version of this experiment. Based on Benini et al. (2022), Kandalowski et al. (2020) and Koch, Frings, and Schuch (2018) and Experiment 1, we decided to collect 48 participants per group. Considering the interaction effects as between-group difference scores, we ran a sensitivity analysis in G*Power (Erdfelder et al., 2009; Faul et al., 2007): with 48 participants per group, we had an 80% probability to detect a medium effect (Cohen’s dz = 0.58) with alpha = .05 with a two-tailed two-sample t test. We collected data from 3 students, who were rewarded with partial course credits. Prolific participants represent most of the sample: we stopped Prolific data collection when we reached 93 submissions having error rates less or equal to 20% when averaged across the conditions. Participants were rewarded with ₤3.3 (₤6.6/h). The experiment was set up such that it was possible to take part only using a laptop or a computer (as opposed to tablets and smartphones) and using Chrome as a browser. Prolific participants were selected based on their age (between 18 and 35 years old), German being their mother tongue, and on being fluent in English. Furthermore, they could not take part if they had already completed similar experiments from us via Prolific. No other filter was used.
Stimuli, tasks, and responses
The target stimuli were drawn from a set of 24 English words representing common objects or animals (see Table 1), and the cues were the words “KIND” and “SIZE.” Target words and cues were presented all capitalised on a white background (800 × 600 pixels) in Arial font size 40 so that they occupied 37 pixels vertically and from 81 to 188 pixels horizontally. Cues were presented 100 pixels above the centre of the screen, while target words were presented at the centre. The task to perform in each trial was signalled by the cue word. The word “SIZE” indicated categorising the word based on its size: bigger or smaller than a shoebox; the word “KIND” indicated categorising the word based on the origin of its referent: human-made or natural. Participants gave their responses by pressing either the A or the L key on their keyboard so that response-key mapping overlapped across the two tasks. Response-key mapping was counterbalanced across participants.
Target stimuli of Experiment 2 and the correct responses to the two tasks.
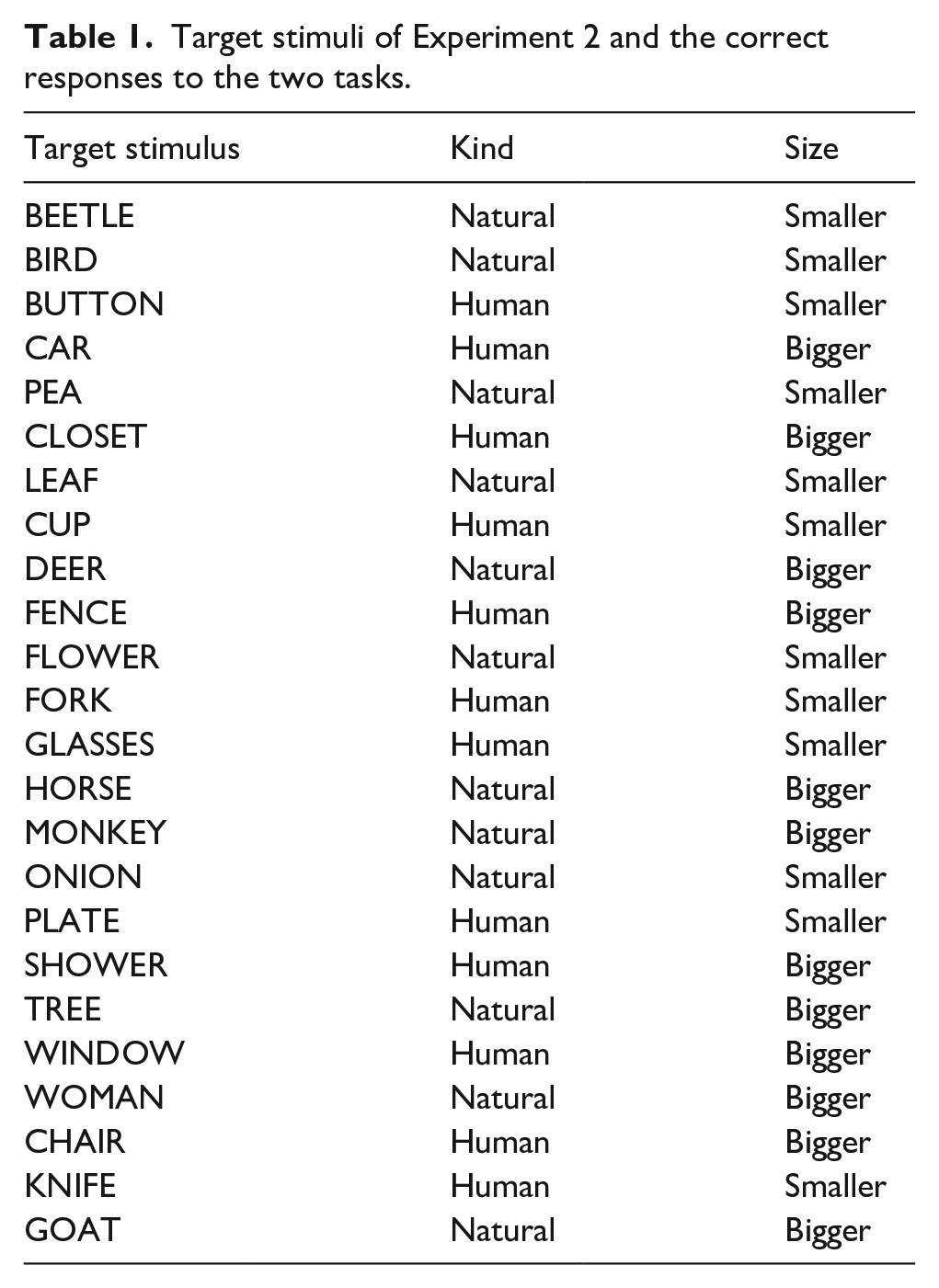
We added a task-irrelevant contextual feature by manipulating the colour of the stimuli. For half of the participants, the context appeared simultaneously with the cue (the cue-related context group): they saw the cue words written in red or in blue, while the target word was always black. Each cue word was written in blue half of the time and red in the other half, and its colour changed (or repeated) unpredictably on a trial-to-trial basis. For the other half of the participants, the context appeared simultaneously with the target (the target-related context group): they saw the target words either in red or in blue, while the cue words were always black. Each target word appeared in blue in half of the times and red in the other half; the target colour switched or repeated unpredictably on a trial-to-trial basis.
Procedure
The experiment was programmed and run using Gorilla Experiment Builder (Anwyl-Irvine et al., 2020). Participants at RWTH Aachen University, who accepted to take part in the experiment, were sent an email that contained a link to access the experiment and a nickname to log in. They could start whenever they wanted, and they had a maximum of 1 hr and 15 min to complete it. Participants in Prolific accessed the link through their accounts on the Prolific website.
Upon clicking on the link, participants were advised to pick a quiet place where they could sit comfortably in front of their laptop/computer. Then they read and accepted the informed consent and the data protection regulation. Before the proper experiment started, participants performed a short test to ensure that they understood the meaning of the 24 target words. Since the categorization tasks require participants to link the written word to the correct object, we asked them to name 24 images representing the 24 targets. In each trial, an image representing a target word appeared together with six possible words: the correct response and five other words randomly drawn from the set of target words. Each word was tested once and thus the test consisted of 24 trials. Participants could proceed to the following image only if they selected the correct response. A maximum of three errors was allowed, above which participants were rejected. In case of a rejection, the participants recruited via Prolific were compensated for the time spent in the experiment. The participants that made three or fewer errors proceeded and read the instructions: these were in English and mentioned the presence of the colour in the cue words (or in the target words), explaining that the colour was not relevant for their tasks.
After the instructions, the participants performed 24 training trials that resembled a portion of a proper experimental block, except for the presence of feedback that was not given in the experimental blocks. Only in such training trials, feedback was provided for both correct and wrong responses and responses slower than 2,000 ms. Each trial started with a black fixation cross (1,400 ms, see Figure 5) followed by the appearance of the cue above the centre of the screen (600 ms). When the cue disappeared, the stimulus appeared at the centre of the screen and remained on the screen until a response was given or 2,500 ms elapsed. The following trial started immediately after, with the onset of the fixation cross.
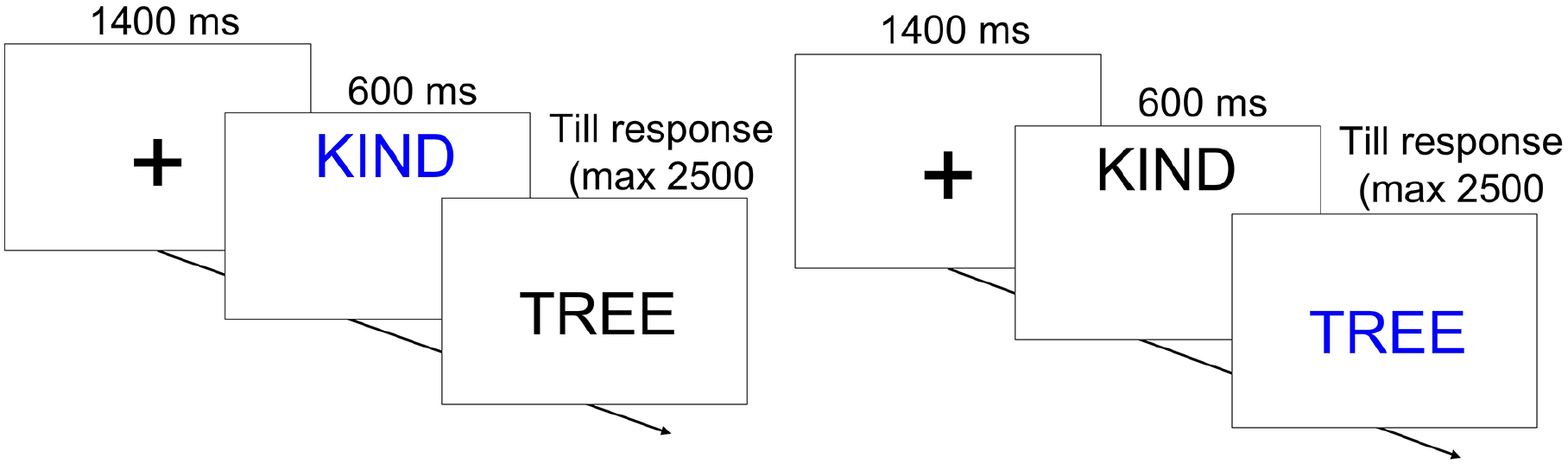
Comparison of the timeline of trials with 0 ms (left) versus 600 ms (right) cue–context onset asynchrony in Experiment 2.
Upon completing the training, participants could decide to read again the instructions (and to perform the training again or not), or they could start the experiment. The experiment consisted of eight blocks of 48 trials each. In each block, the tasks, the target words, and the colours were perfectly uncorrelated so that it was not possible to predict any feature of the current or the upcoming trial. Furthermore, it was never allowed for the same target word to repeat in subsequent trials.
Blocks 1, 2, 7, and 8 were single-task blocks: participants performed only the origin task in two of them, and only the size task in the other two. It was counterbalanced between participants which blocks were origin- or size-only blocks (with always one origin and one size block before and after the mixed-tasks blocks). Blocks 3–6 included were mixed-task blocks, in which origin and size tasks alternated unpredictably, in such a way to minimise the difference between the number of n – 1 task switches and of task repetitions (in each block there was always exactly one task switch more or one task repetition more). Each colour was also associated approximately as often with a task repetition versus switch and a response repetition versus switch. Before each block started, participants were aware of whether the upcoming block contained only origin-task trials, size-task trials, or both.
After completing the eight blocks, participants were thanked and debriefed through a written text explaining the research questions of the experiment.
Design
The design was a 2 × 2 × 2 × 2 mixed design. To test our hypotheses, we ran two ANOVAs with RTs or error rates as the dependent variables, and task relation (repetition vs. switch), response relation (repetition vs. switch), context relation (repetition vs. switch) as the within-subjects independent variables and cue–context onset asynchrony (cue- vs. target-related context) as the between-subjects independent variable.
Results
Analyses of RTs
For this ANOVA, we discarded single-task blocks. 3 Error and post-error trials were also removed, together with the first trial of each block and the responses faster than 200 ms: this implied removing 13.1% of the trials in the four mixed-tasks blocks.
We found a significant main effect of task relation, F(1, 94) = 47.14, p < .001, = .33, indicating task-repetition benefits (842 vs. 894 ms, for task repetitions vs. switches, respectively). Task relation also interacted with response relation, F(1, 94) = 20.2, p < .001, = .17, such that there were RR benefits in task repetitions (16 ms) but costs in task switches (−28 ms).
We found a significant four-way interaction of task relation, response relation, context relation, and context onset, F(1, 94) = 10.36, p = .002, = .10. To further investigate this interaction, we divided the data according to cue–context onset asynchrony (i.e., cue-related vs. target-related context), and we ran a three-way within-subjects ANOVA on each group. Only when the context appeared simultaneously with the cue (see Figure 6), task relation, response relation, and context relation entered in a significant three-way interaction, F(1, 47) = 11.57, p = .001, = .12, replicating the pattern already observed in Experiment 1. In contrast, the same interaction was not significant in the target-related context (see Figure 7), F(1, 47) = 1.21, p = .277, = .02. Following the same procedure as in Experiment 1, we ran two separate within-subjects ANOVAs on the cue-related context group dividing task repetitions from task switches. Results showed that context relation significantly interacted with response relation in task repetitions, F(1, 47) = 4.67, p = .036, = .09, and in task switches, F(1, 47) = 7.08, p = .011, = .13. In task repetitions, we found greater RR benefits in context repetition (27 ms) than in context switches (−13 ms); this is the same pattern that we observed in Experiment 1. In task switches, we found greater RR costs in context repetition (−60 ms) than in context switch trials (−7 ms), a pattern that we did not find in Experiment 1. No other main effect or interaction was significant (all Fs < 2.92 and all ps > .091).
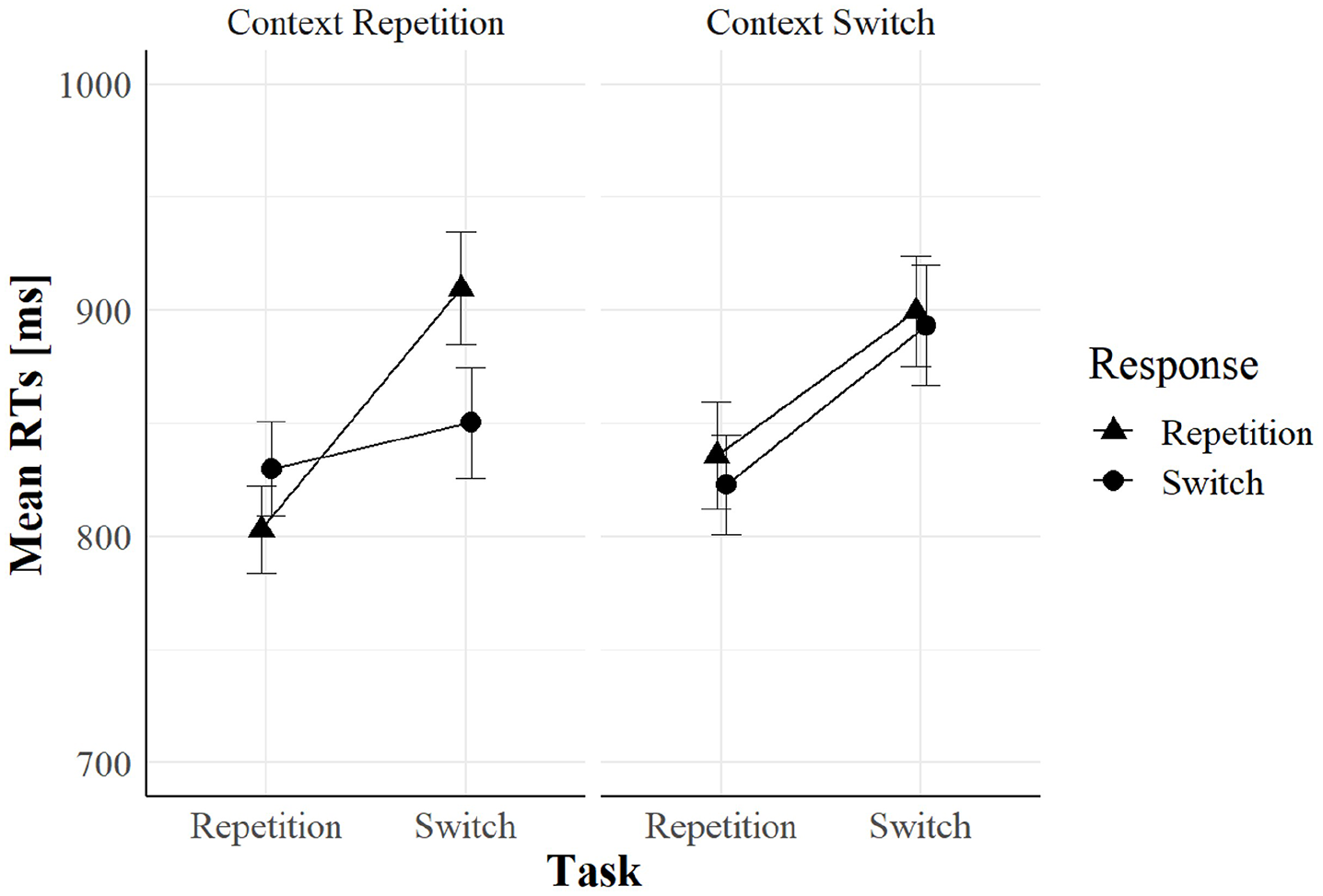
Mean RTs when cue–context onset asynchrony equal to 0 ms (the cue-related context) as a function of task relation, response relation, and context relation in Experiment 2.
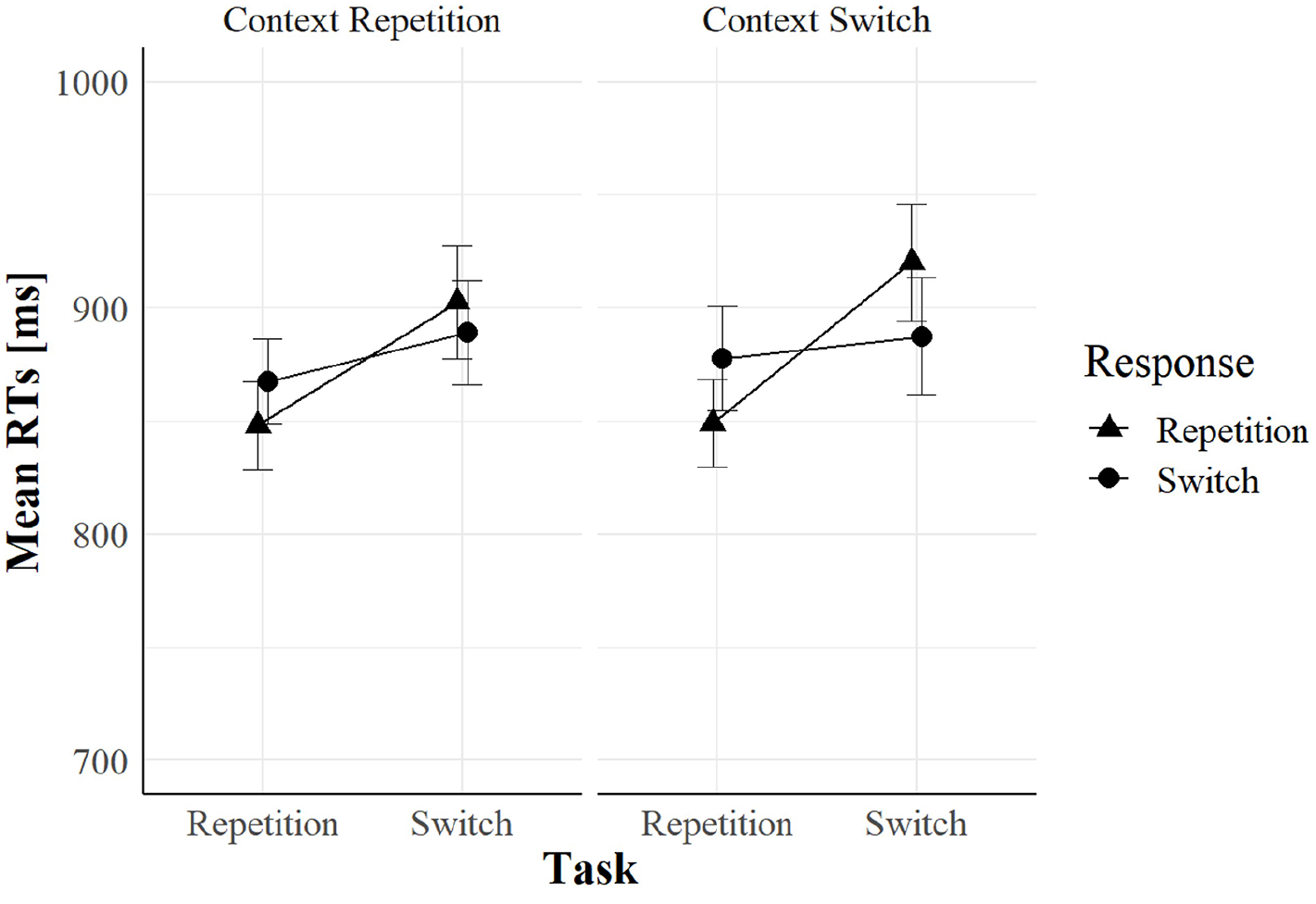
Mean RTs when cue–context onset asynchrony equal to 600 ms (the target-related context) as a function of task relation, response relation, and context relation in Experiment 2.
Analyses of error rates
For this ANOVA, we discarded single task blocks 4 together with the first trial of each block, trials faster than 200 ms and post-error trials. This implied removing 7.9% of the four mixed-tasks blocks.
We found a significant main effect of task relation, F(1, 94) = 33.99, p < .001, = .26, indicating task-repetition benefits (4.5% vs. 7.1%, for task repetitions vs. switches, respectively). Differently from the RTs, the main effect of response relation was also significant, F(1, 94) = 31.04, p < .001, = .25, indicating general RR costs (7.1% vs. 4.4%, for response repetitions vs. switches, respectively). Task- and response relation further entered in a significant two-way interaction, F(1, 94) = 28.05, p < .001, = .23, so that RR costs were smaller in task repetitions (−0.52%) than in task switches (−4.84%). Thus, as in Experiment 1, repeating the response in task repetitions yielded costs in the ERs but benefits in the RTs.
No other main effect or interaction was significant (all Fs < 2.21, all ps > 0.141). Hence, we observed no main effect of context relation and neither did it interact with task relation and response relation, as we found in the RTs.
Discussion
With the present experiment, we replicated the main findings of Experiment 1 regarding a task-irrelevant context feature in an independent sample. Context relation did not affect the error rates in this experiment; therefore, we will only discuss RTs results.
As in Experiment 1, we found evidence suggesting that the cue colour may be bound into an event file with task and response in each trial. If so, it will retrieve the event file when it repeats in the following trial. Therefore, if also the other stored features repeat, performance should improve compared with context switches, and this is what we found in the cue-related context group in Experiment 2.
In the target-related context group, we found no modulation of context relation. Visually inspecting Figure 7, the data pattern is very similar between the context repetitions and context switches in the target-related context group, with the typical crossover interaction of response relation and task relation. The same crossover pattern was also found in the cue-related context group when the cue colour repeated (see Figure 6, context repetitions, left panel). These findings are consistent with each other, given that every trial was a cue–colour repetition in the target-related context group (the cue word was always black).
We argue that the absence of context effects in the target-related context group is due to the context onset timing. In other words, applying the context to the target yielded the same effect as applying a 300 ms cue–context onset asynchrony in Experiment 1. This finding is of importance as one could argue that the delayed context was ignored in Experiment 1 (and, thus, had no influence). Yet, the context could not be ignored in the target-related context group in Experiment 2. On the contrary, a target-related context, as well as a cue-related context, must be encoded to successfully execute the task. We will address this issue in the “General discussion.”
General discussion
In the present study, we investigated how task-switching performance was affected by the trial-by-trial repetition (vs. switch) of a task-irrelevant feature. Since the feature was neither relevant to the task, nor was it associated with a response, we refer to it as “context.” Importantly, we varied the timing of context onset: in Experiment 1, the context could appear either synchronously with the task cue, or 300 ms later (i.e., 300 ms before target onset). In Experiment 2, it could appear either synchronously with the task cue or with the target (i.e., 600 ms after the cue).
In Experiment 1, the context was the colour of the cue rectangle, while the orientation of the rectangle indicated the task to perform. In those blocks in which cue–context onset asynchrony was equal to 300 ms, the cue was black for the first 300 ms and then it turned blue or red. In Experiment 2, the context was the colour of the ink used for cue or target words. In the cue-related context group, cue words were written either in red or blue and target words were always black. In the target-related context group, cue words were always black, while target words were either blue or red.
In both experiments, the effect of context relation accrued from two main findings. The first concerns the modulating effect that context relation exerted on task and response relation. We found RR benefits in task repetitions when the context (i.e., the colour) repeated in the subsequent trial, but not when the context switched from the previous trial (see Figures 2 and 6).
The second main finding concerns the effect of varying context onset. In Experiment 1, cue–context onset asynchrony modulated the interaction of task relation and context relation. When cue–context onset asynchrony was 0 ms, so that the colour appeared together with the cue, task repetition benefits were greater in context repetition than in context switch trials. In contrast, when cue–context onset asynchrony was 300 ms, task-repetition benefits did not differ in context repetitions and context switches. In Experiment 2, context onset determined the presence of RR benefits in task repetition trials when the context repeated (vs. the absence when the context switched). In other words, we found larger RR benefits in task repetitions when the context repeated in the cue-related group but not in the target-related group.
Binding and retrieval of a task-irrelevant context feature
Episodic retrieval accounts, which postulate feature binding and retrieval processes, provide a parsimonious framework to discuss the effect of switching versus repeating a task-irrelevant context feature in task switching. For example, the BRAC framework (Frings et al., 2020) maintains that partial repetition costs in sequential paradigms, like task switching, arise due to feature binding (in trial n − 1) and retrieval (in trial n) processes. Indeed, we found partial repetition costs as the performance was better in full repetitions (when also the context repeated) than in partial repetitions (when the context switched). The pattern is consistent with the context being bound together with the task and response in the previous trial so that, when the context repeated in the following trial, it retrieved the task and the response (i.e., context-triggered retrieval due to context repetition), resulting in RR benefits in task repetitions.
BRAC also maintains a distinction between binding and retrieval processes. We argue that RR benefits differed in context repetitions versus context switches due to differences in retrieval as opposed to differences in binding. For the effect to be visible in context repetitions, the context needed to be always bound during the previous trial. Since participants could not predict whether the context would have repeated in the following trial, this implies that the context was bound in every trial, and when the same colour appeared again, the stored event file could be retrieved. With respect to our second finding, below we argue that cue–context onset asynchrony might modulate performance by modulating context binding, as opposed to retrieval, especially in Experiment 1.
However, the data pattern observed in our study is not fully in line with an episodic retrieval account and the assumption of a cue-triggered retrieval process. More specifically, if the repetition of the cue colour triggered retrieval of the previous episode, we would also predict worse performance in task switches/response switches in which the cue colour repeated (partial repetitions) than in task switches/response switches in which the cue colour switched (full switches). That is, both TEC (Hommel, 1998, 2019) and BRAC (Frings et al., 2020) claim that a full switch should be faster when also the context switches since no retrieval takes place when all features switch. In contrast, in our study, context relation did not systematically modulate performance in task switches. The only effect occurred in the cue-related context group of Experiment 2, in which the condition in which task, response and context all switched was slower than the partial repetition condition in which the context repeated but the task and the response switched. Therefore, we did not observe systematic benefits in full switches as compared with partial repetitions.
Although the absence of a systematic influence of context repetitions versus switches on task switches is not fully consistent with the assumption of a context-triggered retrieval of the previous episode, it would be consistent with a cue-triggered retrieval of the previous episode. Accordingly, repeating the task-relevant cue dimension (i.e., cue orientation) retrieves the previous episode. If both the response and the context repeat from the previous trial, we would observe a performance benefit (i.e., full repetition benefits). If, however, either the context or the response switches, we would observe a performance cost (i.e., partial repetition costs). Moreover, switching the cue orientation would not retrieve the previous episode, thus, performance would not be affected by a context switch versus repetition. In this way, the context repetition itself would not trigger the retrieval of the previous episode so that performance in task switches would not be affected by context relation. However, a context repetition/switch would nevertheless cause performance benefits/costs in task repetitions.
In Experiment 1, in task repetitions, an equally oriented rectangle is presented in the same colour on context repetitions, but in a different colour in context switches. Thus, our data are also consistent with a further alternative explanation that it is the combined occurrence of cue and context that triggers retrieval of the previous episode. In terms of episodic binding, one would then argue that only the repetition of a cue–context combination would retrieve the previous episode. This might also explain why the context only influenced performance when it was combined with the cue but not when it was presented later than the cue.
Such combined repetition of both the cue and the context is nothing but an exact cue repetition. Therefore, benefits observed in this condition are also consistent with a cue-encoding priming phenomenon, as put forward by Logan and Bundesen (2003) who used a 2:1 cue:task mapping. Later studies that used a 2:1 cue:task mapping (e.g., Forrest et al., 2014; Monsell & Mizon, 2006; Schneider & Logan, 2011) consistently found smaller task repetitions benefits when the cue switched. This suggests that cue-encoding priming contributes to task repetition benefits, albeit not entirely. Importantly, the context manipulation we used (see also Benini et al., 2022; Kandalowski et al., 2020; Koch, Frings, & Schuch, 2018) allows a cue repetition with a context switch, which is not possible with a 2:1 cue:task mapping. Therefore, despite both accounts (i.e., cue-encoding priming and features binding) unsurprisingly predict benefits for repeating the task, the context and the response, the present study is designed to tackle a different research question, namely context binding in task switching.
The effect of cue–context onset asynchrony on feature bindings
The second main finding of our study tackles a modulation of binding and retrieval processes due to feature onset asynchrony. We did not observe any effect of switching versus repeating the context when there was a cue–context onset asynchrony.
In contrast to our study, Rubin and Koch (2006) presented a delayed context (together with the target) and found evidence for context-triggered retrieval. However, they found such evidence only when the current task was not sufficiently prepared (i.e., with a short, but not with a long CSI).
Along these lines, we argue that a context that appeared 300 or 600 ms (and not 0 ms) after the cue triggered retrieval processes when the currently relevant task was already prepared to a sufficient extent due to cue-based activation. Interestingly, this argument is consistent with Arrington and colleagues’ (2007) suggestion that the time needed to encode the cue and retrieve the task may be around 300 ms (p. 496).
In general, the absence of context-based retrieval effects with 300 or 600 ms context onset asynchrony is consistent with a horserace model (e.g., Tipper, 1985; see also Frings & Moeller, 2012). In order to observe context-based retrieval effects, the context needs to be encoded early enough and for a sufficient amount of time: delaying its onset, therefore, reduced the impact of context-based retrieval processes compared with the other ongoing processes triggered by task-relevant features or stimuli. In fact, the horserace model would predict a larger retrieval effect if the context appeared before the cue.
Yet, there might be an alternative explanation for reduced context effects in conditions with a delay between cue and context. That is, in Experiment 1, a 300 ms cue–context onset asynchrony arguably further reduced the salience of the context, which could have been ignored. Put differently, there was no need to encode the context when it was not associated with a task-relevant feature. As a consequence, as far as Experiment 1 is concerned, asynchrony might have weakened retrieval effects by weakening context encoding, or even context binding in the first place. If this was the case, a context that appears before the cue would yield smaller retrieval effects, as it would lose salience by losing belongingness to the episode.
However, this explanation is not valid for Experiment 2, in which the delayed context appeared together with the target. This manipulation did not reduce the salience of the context, which must have been encoded since the target must be encoded, just like the cue in Experiment 1. Here, pairing the context with the target might have caused the context-triggered retrieval processes to be masked by the target-triggered processes like target encoding and response selection.
Speculating that (the absence of) binding and/or retrieval of the context might be responsible for the influence of cue–context asynchrony implies a distinction between binding and retrieval processes (cf. Frings et al., 2020). Whereas we assume that retrieval processes (rather than binding processes) can explain the influence of context relation on RR effects per se (our first main finding), we argue that also binding processes might contribute to explain the modulation of context effects due to the cue–context asynchrony (our second main finding).
Conclusion
In conclusion, across two cue–task–switching experiments, we showed that a task-irrelevant contextual feature affected participants’ performance. Particularly, our data are consistent with an episodic retrieval account postulating that every feature of a trial is bound together into a mental representation of the episode, which is retrieved upon repetition of at least one feature. Importantly, binding effects were observed only when the irrelevant context feature onset coincided with the cue onset, and not when the onset was delayed by 300 ms or it coincided with the target onset (i.e., a 600-ms delay). Along the lines of the horserace model (e.g., Tipper, 1985; see also Frings & Moeller, 2012), we claim that a delayed onset of the context disadvantaged the processes triggered by such an irrelevant feature compared with the task-relevant processes, thus impacting retrieval effectiveness. Alternatively, we suggest that a delayed context was less strongly bound with the other features. Thus, whereas the presence of partial repetition costs is consistent with context retrieval processes (but not binding), their modulation by context onset asynchrony is consistent with both (the absence of) context-binding and/or retrieval processes.
In the present study, we showed that performance in multitasking settings is affected by feature binding mechanisms that can also involve task-irrelevant features. However, these mechanisms are observable only when the irrelevant features appear early enough.
Footnotes
Acknowledgements
The authors thank Růžena Ceralova for her valuable help throughout experiment programming, data collection, and data preprocessing.
Authors’ note
Some of the data reported in the manuscript were presented at the 63rd edition of the Tagung experimentell arbeitender Psychologen (Conference of Experimental Psychologists) in March 2021.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: the research reported in this article was supported by the Deutsche Forschungsgemeinschaft (Research Group: FOR 2790 [Grant No. PH156/4-1]).
Data availability
All the data (http://doi.org/10.23668/psycharchives.5116) and the code used to run the analyses (![]() ), including auxiliary analyses, are made available online. The study materials are available on request from the corresponding author.
), including auxiliary analyses, are made available online. The study materials are available on request from the corresponding author.