
Abstract
Reactive and proactive cognitive control are fundamental for guiding complex human behaviour. In two experiments, we evaluated the role of both types of cognitive control in navigational search. Participants searched for a single hidden target in a floor array where the salience at the search locations varied (flashing or static lights). An a-priori rule of the probable location of the target (either under a static or a flashing light) was provided at the start of each experiment. Both experiments demonstrated a bias towards rule-adherent locations. Search errors, measured as revisits, were more likely to occur under the flashing rule for searching flashing locations, regardless of the salience of target location in Experiment 1 and at rule-congruent (flashing) locations in Experiment 2. Consistent with dual mechanisms of control, rule-adherent search was explained by engaging proactive control to guide goal-maintained search behaviour and by engaging reactive control to avoid revisits to salient (flashing) locations. Experiment 2 provided direct evidence for dual mechanisms of control using a Dot Pattern Expectancy task to distinguish the dominant control mode for a participant. Participants with a reactive control mode generated more revisits to salient (flashing) locations. These data point to complementary roles for proactive and reactive control in guiding navigational search and propose a novel framework for interpreting navigational search.
Searching for misplaced keys around the house can be a frustrating, tiring, and time-consuming activity. For most people, habitual behaviours and a good memory probably offset this disruption, but for others, their ability to function efficiently in everyday search tasks is easily compromised. Unlike small scale visual search (e.g., Wolfe, 1996), navigational search, in which the participant has to move through the environment to find the target, places a physical and cognitive demand on the searcher. Navigational search requires a goal focus (e.g., where are my keys most likely to be placed?) and resisting distraction (e.g., should I look for a spare set of keys?) or repetitious search errors (e.g., I’ll check my coat pocket yet again). As the decision to continue the search for keys (even in a previous location) can be costly, cognitive control of monitoring goals and deploying attention to navigational search would seem important to search success. Here, we report two studies that probed the cognitive control of navigational search in a partially controlled laboratory environment that assessed search locations visited and search errors made across a relatively dense array of competing search locations.
Foraging refers to search for single or multiple targets across multiple locations where there are rewards associated with finding the target(s) (Stephens & Krebs, 1986). Foraging is a fundamental and central adaptive behaviour in all animals because it supports finding food and maintains reproductive success (Hill & Dunbar, 2002). Navigational search is a subtype of foraging in which the search is for a single target, and the reward is less directly linked to finding the target (c.f. finding food). A novel, automated paradigm has been developed to measure navigational search in humans (Smith et al., 2005, 2007). In this paradigm, the goal is to find a single target location among multiple locations in a specially adapted floor in which is embedded an array of coloured lights. Using this paradigm, visually guided navigational search was found to be a good analogue of visual search (Gilchrist et al., 2001; Smith et al., 2007), although another variant of this paradigm where the target was hidden was not analogous to visually-guided search (Smith et al., 2005). In a study with children, Smith et al. (2005) asked participants to engage in navigational search for a hidden red target light. The target light was “hidden” under a green light and could only be activated by a simple switch (button press) at each potential location. In this hidden variant, navigational search showed fewer search errors to previously visited locations (revisits) compared with classic visual search (see also Gilchrist et al., 2001; Smith et al., 2008) implying that navigational search is not a simple corollary of visual search. Navigational search informs different search problems, such as exhaustive search for hidden targets in open ground (Riggs et al., 2017), or rummaging for unknown targets within complex environments (Riggs et al., 2018).
Successful human foraging is due, in part, to our ability to accrue learning-based knowledge to shape navigational search (Bock, 2004; Goldstone & Ashpole, 2004; Riggs et al., 2018). In the navigational search paradigm, effective search has been linked to an individual’s spatial working memory capacity and to their sensitivity to the probabilistic likelihood of where the target is located over multiple trials (Smith et al., 2005, 2010). As noted by Smith et al., the ability to learn across multiple trials in this paradigm originates from the ability to remember and update both allocentric and egocentric visual representations of the search space in working memory during search (Smith et al., 2010). Working memory updating is valuable for learning about the characteristics of changing search environments, but a broad range of automatic and controlled cognitive processes can also motivate search. Within the classic Norman–Shallice framework of attentional control of executive function (Norman & Shallice, 1986; Shallice & Burgess, 1996), an action schema, such as walking to a specific location, is familiar and can be activated in a relatively automatic manner. When multiple action schemas compete, then inhibition and suppression of competing action schemas result in the activation of one “optimal” action schema. This framework further posits a Supervisory Attentional System that supports deliberate planning of action, with flexible use of strategies to generate novel, or non-habitual, action schemas. Experiment 1 reported here aimed to clarify the relation between cognitive control systems and navigational search.
To date, the automated navigational search paradigm has provided a range of evidence for the role of cognitive control in this class of tasks. Smith et al. (2005) found that children’s use of their non-dominant hand compromised search efficiency. Navigational search was less efficient when conflict arose between the resource demands of updating working memory to motivate search and the need to inhibit automatic and prepotent behaviour (using the dominant hand). Pellicano et al. (2011) observed fewer instances of systematic and optimal navigational search by children with Autistic Spectrum Disorder that was associated with their impaired performance on tasks which measured cognitive control. Finally, Longstaffe et al. (2014) identified three search indices presumed to require resource allocation: (a) the ability to direct attention to likely target locations, (b) the ability to remember previously visited locations, and (c) the ability to inhibit distracting information in the search array. Longstaffe et al. (2014, Experiment 1) explored perceptual salience of locations within the search space by having flashing and static lights at different locations across the search display. Having multiple search locations creates competition among different possible action schemas, the allocation of attentional resources to resolving this competition should improve success and efficiency of search. Likewise, any automatic re-direction of attention towards the perceptually salient flashing locations would have consequences for resolving conflict between action schemas. Longstaffe et al. found that the presence of perceptual salient flashing locations increased the likelihood of revisits to previously visited locations. By assuming that working memory supports the retention of spatial co-ordinates (i.e., maps) of possible locations, one explanation was that the effortful “tagging” of visited locations allows an updated record of the search space (for active and visited locations) to be maintained. These data implied that the allocation of attention to salient flashing locations impinged on the ability to update working memory after a search event, and therefore maintain search efficiency.
In a further study, Longstaffe et al. (2014, Experiment 2) found that the salience effects on search errors were diminished when the illuminated locations were extinguished after being visited. Assuming effortful “tagging” of visited locations placed demands on working memory updating under conditions of continued illumination, then these visited locations were no longer required to be “tagged” when lights were extinguished. This manipulation increased the cognitive resources available for retaining and updating the search display leading to more efficient search. Longstaffe et al. (2014) therefore posited evidence in favour of a single resource-driven account of navigational search where search at perceptually salient locations competes for common resources that are shared with the working memory load of retaining visuo-spatial locations within the search display. One problem with this account is that it fails to consider “higher level” processes of supervisory control that facilitate strategic governance of navigational search, and distinguish these from resources for “lower level” processes that automatically resolve interference between competing action schemas to select the most optimal action schema. Certainly, our everyday experience suggests that explicit rules do motivate search (e.g., my keys are kept on the hook).
One contemporary theory of cognitive control (dual-mechanisms control theory; Braver, 2012; Braver et al., 2007, 2009) builds on the classic formulation of different components of control within the Norman–Shallice attentional framework. This model postulates two distinct mechanisms of proactive and reactive control. Proactive control orients to the goal-relevant information that biases action, perception, and attention systems to maintain the goal-relevant behaviour (Miller & Cohen, 2001). As such, proactive control reflects the sustained maintenance of goal-relevant information in an anticipatory fashion. By contrast, reactive control is transient and stimulus-driven and can occur post-event to direct goal re-activation. It incorporates interference demands and episodic associations with the context of event stimuli and goal activity. Behavioural evidence for the dual-mechanisms framework of cognitive control comes from a range of cognitive tasks including variants of the Stroop task and task switching (Bugg & Braver, 2016; Gonthier et al., 2016) and the AX-Continuous Performance Task (AX-CPT; Cohen et al., 1999; MacDonald, 2008) which we use in Experiment 2 of the current study.
Experiment 1 aimed to explore the utility of a dual-mechanisms account of cognitive control in accounting for performance in the navigational search paradigm. To do this, a rule-manipulation was implemented at the start of search, and the perceptual salience of search locations was manipulated by having both “flashing” or “non-flashing” locations in the search array. Extending the dual-mechanisms account to navigational search, proactive control implies search behaviour is sensitive to explicit rules (provided before search commences) by allocating resources to sustaining a rule, and by minimising distraction to adhere to a rule. For each participant, two rule manipulations were introduced, participants were told at the start of each block that either (A) the target was more likely to be at a non-flashing (static) location or (B) the target was more likely to be at a flashing location. Here, the a-priori rule was directly linked to the perceptual salience of locations, not other aspects of the search display. If proactive control facilitates the maintenance of the a-priori rule to minimise distraction, then rule-congruent search should occur as participants (a) attend to the perceptual salience of the display and (b) resolve the cognitive demands of resisting competing action schemas to alternative locations, predicting more button presses at rule-congruent locations, that is, more presses to static locations under Rule A and more presses at flashing locations under Rule B. These predictions differ from a simpler hypothesis that only the perceptual salience of the display orients navigational search leading to more button presses at the salient flashing locations, irrespective of the rule.
The second feature of search behaviour in this task is revisiting previously searched locations; revisits are costly search errors with respect to the goal of finding the target. The findings of Longstaffe et al., (2014) that increasing the uncertainty of whether a location had been visited or not (e.g., by keeping targets switched on after a search event) increased revisits, but not the overall number of button presses, implying different factors influence these measures. According to a dual-mechanisms cognitive control account, reactive control corresponds to late-stage recruitment of resources to respond to new information by monitoring stimuli-driven aspects of the display and to re-instate the goal. Each search event (of reaching down to check a specific target) potentially increases the need to engage reactive control to monitor the information provided at the search location, for example, by actively updating working memory, and to re-instate the search goal by resolving competition between multiple action schemas. If the demands for reactive control increase, then updating working memory and goal-reinstatement will be compromised by diminished proactive control and more search errors will be made. As the allocation of reactive control is a direct response to stimuli and context, flashing locations will induce more conflict between different action schemas, influencing activity towards these locations. Under a reactive control hypothesis, we predict that participants will make more revisits to flashing search locations, and more revisits under the flashing rule that requires monitoring of flashing locations, as search efficiency is compromised.
Experiment 1
Method
Participants
A total of 47 participants (20 males, 27 females) were recruited as undergraduate volunteers, with participation in lieu of course credits. All participants gave informed written consent and were debriefed about the experiment. The study was approved by the local ethics committee. All participants self-reported no hearing problems and an average level of fitness.
Task design and materials
Automated navigational search paradigm
The paradigm consisted of a darkened room in a dedicated 4 × 4 m2 space surrounded by black featureless curtains so that all discernible landmarks were removed. The space containing 49 search locations embedded in the floor in a honeycomb arrangement. Each search location consisted of a steel button (diameter = 2.5 cm) surrounded in a ring of light-emitting diodes (LEDs, diameter = 6 cm). The centre of each light was 38 cm from the centre of the adjoining light on each side and 50 cm from the nearest light in a circular trajectory around the border. One location was consistently lit as orange at the start of each trial to identify a starting location. A set of 20 active search locations were lit green and were selected randomly from the array of possible locations on each trial. Of these, 10 locations were selected to flash in the salience manipulation (see Figure 1).

Example trial in the navigational search space with locations displayed as flashing (dark grey dot pattern), static (light grey), and the starting location (in black).
The flashing lights alternated at less than 2 Hz to minimise the risk of epileptic seizure (Bancaud et al., 1981) and alternated at varying frequencies to ensure that the set did not flash in a synchronised manner. The viewing range of locations depends on their head and body orientation at any given time. Each trial began at the same start location as indicated by an orange LED and commenced with a single button press at this location. Participants were then required to engage in navigational search for a hidden target and inspect locations with a button press to the centre of a single green, illuminated LED. The target was “found” when the colour at the location switched from green to red when the location was “inspected.” All non-target locations remained illuminated as green after inspection. The trial ended when the target location was found. The search task consisted of two blocks of 20 search trials. For each block, participants were provided with one rule (of two) to bias their knowledge of the likely location of the target. Under Rule A, participants were told that 60% of the targets would be at static locations across the block. Under Rule B, participants were told that 60% of the targets would be at flashing locations across the block. The order of presentation of rules was counterbalanced. Search behaviour was measured by the number of button presses overall and the number of revisits to previously searched locations on each trial.
Procedure
Participants attended a single test session that was split between a dedicated navigational search room and an adjoining laboratory room. On entering the search room, participants were introduced to the search array and informed that all non-illuminated buttons served no function and that each trial had only one stationary target, indexed by a switch from a green to red light when pressed. Participants were instructed on the goal of searching across the illuminated locations, at their own pace, to determine the location of the hidden red target. Participants were then positioned in the starting location such that all locations in the search array were visible. At the beginning of each block, participants were informed of a specific rule regarding the likely location of the target. The experimenter then vacated the room and instructed the participant from outside the room, with access via remote camera, to commence the trial. After the start button was activated, participants walked through the search array and pressed illuminated locations until the target location was identified. On finding the target location, the search array was reset and the participant was instructed to return to the starting position for the following trial. Participants were offered a short break between the two blocks, and were advised of the second rule, prior to the second block. Overall testing time was approximately 40 min and participants were fully debriefed.
Data analysis
Using JASP (The JASP Team, 2018), data sets were examined with analysis of variance (ANOVA).
Results
Figure 2a illustrates the effect of the rule on the relative number of button presses to flashing and no-flashing locations. The red line indicates the balance point in which the number of presses to flashing and non-flashing locations is matched. Overall, the trend is for button presses to be congruent with rule. To test the specific prediction of rule-congruent navigation in the overall button presses, one-way comparisons were conducted for presses at flashing locations under each rule. For the flashing Rule B, more button presses were made to flashing locations, M = 0.53, t0.5(45) = 3.17, p < .01, Cohen’s d = .47, 95% CI = [0.16, 0.77]. For the static Rule A, fewer button presses were made to flashing locations although this approximated to chance performance, M = 0.48, t0.5(45) = −1.26, p = .18, Cohen’s d = −.17, 95% CI = [−.48, 0.11]. There was evidence for rule-adherent search at flashing locations when implementing the flashing rule. If rule-adherence to the flashing rule is governed by the ability to maintain the rule in working memory, then the length of search could influence the probability of maintaining the rule in memory. That is, longer search durations could impede the likelihood of maintaining rule-governed behaviour. To probe this, search time to locate the target were extracted on a trial-by-trial basis across conditions, for each person. Trials with Slow and Fast search times were categorised based on a median split for each person (Mslow = 30 s and Mfast = 9.7 s) and within each condition. If rule-adherence to the flashing rule can only be maintained over short durations, then we anticipated a rule × speed interaction, where the advantage for maintaining the flashing rule would be evident in the Fast trials only. To test this, a 2 (rule; A or B) × 2 (salience of target location; Flashing or Static location) × 2 (search duration; Fast or Slow trials) ANOVA (Type III) was conducted on search time. Search times were not significantly faster according to rule A or B, F(1, 45) = 3.42, p = .07,
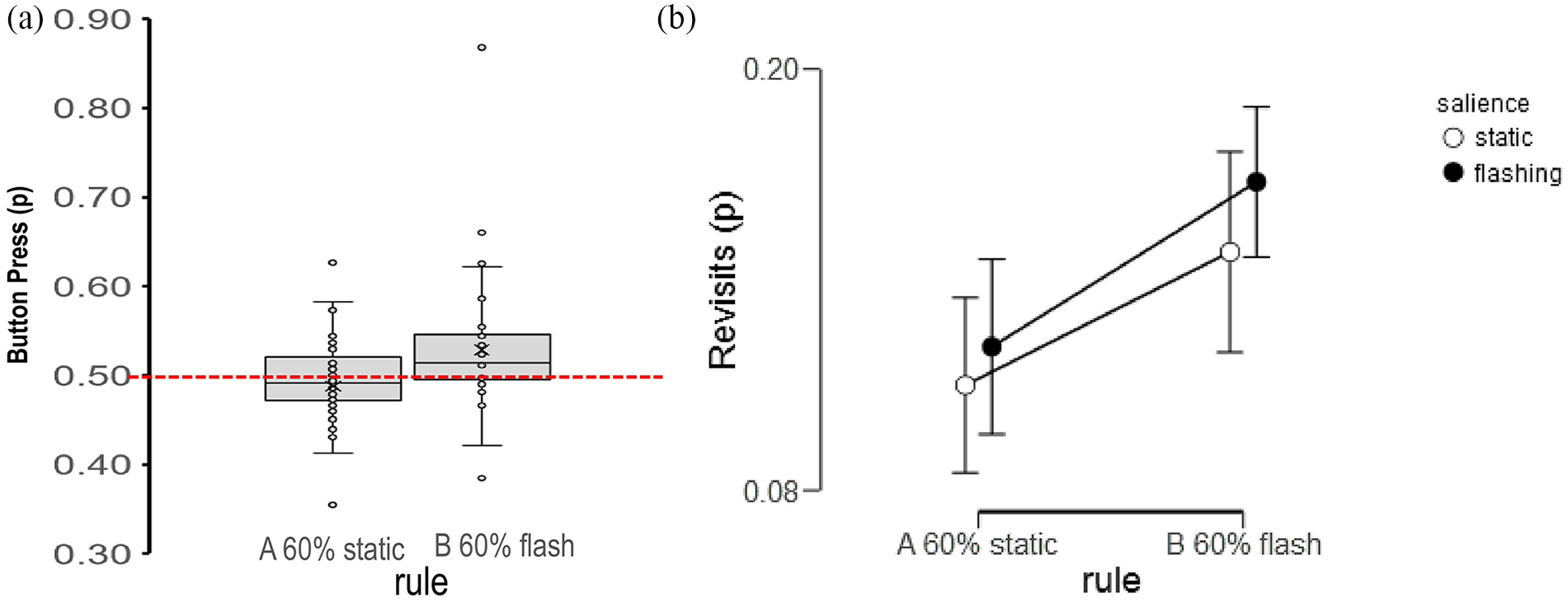
Displays the average proportions of (a) button presses and (b) revisits generated under a probabilistic rule for locating targets at static (Rule A) and flashing locations (Rule B). (a) Boxplots for flashing locations only due to non-independence. (b) Revisits as a function of the salience of search locations.
Figure 2b plots the proportion of revisits by rule. There were both more revisits under the flashing rule and more revisits to flashing locations. In a 2 (rule; A and B) × 2 (salience of target location; Flashing and Static) repeated measures ANOVA (Type III) on the proportion of revisits, there were significantly more revisits under the flashing rule, compared with static rule, F(46) = 4.64, p = .04,
Discussion
The role of proactive and reactive control in navigational search was investigated by manipulating the a-priori goal and the salience of search locations in the display. We used the overall proportion of button presses and the proportion of revisits as measures of navigational search performance. The findings were consistent with a dual-mechanism account of cognitive control (Braver, 2012) that distinguishes proactive and reactive control, and provides a novel interpretation framework for navigational search behaviour. Proactive control was anticipated to govern navigational search by supporting participant’s adherence to an a-priori rule and so leading to participants pressing buttons at spatial locations congruent with the rule. The findings provided evidence for rule-adherence in search, when the rule specified the target was more likely at flashing locations. For revisits to previous locations, there was evidence that instigating the flashing rule led to more revisits overall (irrespective of location) consistent with reactive control. There was no evidence that search durations varied systematically across conditions or that the sensitivity of search durations to the flashing rule occurred only for faster trials. Navigational search times almost certainly tap into the participant’s agility and motoric fitness and so should not be considered similar to search times in visual search tasks. Specifically, the duration of navigational search times corresponds largely to time spent travelling between locations, not time allocated to a specific location. Since search times were not directly sensitive to the salience of display characteristics, then the allocation of cognitive control in guiding search behaviour can offer a valuable explanation beyond describing navigational search as a function of the display.
Although the dual-mechanisms account offers an interesting explanatory account of search patterns, we acknowledge the limits of post hoc interpretation of these search data. To address this, Experiment 2 aimed to provide evidence of associations between navigational search and dual mechanisms of cognitive control, as indexed by the Dot Pattern Expectancy (DPE) task. This task is commonly used to identify control modes (Lopez-Garcia et al., 2016; MacDonald, 2008; MacDonald et al., 2005; also meta-analysis by Janowich & Cavanagh, 2018). As a variant of the expectancy-CPT (Cohen et al., 1999; MacDonald, 2008; MacDonald et al., 2005), the DPE uses visual patterns of simple dot arrays as cues and probes (instead of familiar letters) and was developed to be more appropriate for high-functioning individuals and non-clinical populations (Lopez-Garcia et al., 2016). The DPE task requires participants to distinguish one target pairing of a cue–probe association (e.g., cue A–probe X) from among three types of alternative (non-target) cue-probe pairs that are all presented in a continuous sequential stream. As a monitoring task, participants need to maintain cognitive control for detecting and implementing positive identification of the target (e.g., cue A–probe X) and negative responses to the non-target pairs (e.g., cue B followed by probe Y as a non-target baseline). Critically, the target (A-X) pair is high frequency (typically 65%–70% of cue–probe associations) compared with non-target pairings; this generates a strong associative and prepotent response tendency for positive identification of A-X pairs.
Dual-mechanisms control on the DPE task is evident from responses to non-target “interference” trials, for example, when cue A is followed by probe Y (A-Y pairing) or when cue B is followed by probe X (B-X pairing). Proactive control co-ordinates the anticipatory maintenance of a goal, such as determining a correct response only to the A-X pairs. After presentation of an A-cue, the A-Y pairing generates conflict between anticipation of an X-probe, and negative response to the Y-probe. This generates errors and/or longer latencies of responses to the non-target A-Y pairing, as evidence of proactive control mode. Reactive control governs the late-onset allocation of resources to resolve a response in the event of detecting the stimuli and facilitates re-instatement of the goal. For the non-target baseline (B-Y pairing), response certainty is high given low conflict with the target A-X pair, but a non-target X probe (in B-X pairings) will generate conflict that is resolved by late-stage allocation of reactive control mode. As patterns of errors and response latencies distinguish the A-Y and B-X pairings from the target (A-X) and baseline (A-Y) trials, individual biases for control mode can be derived. Individual bias scores for proactive and reactive control mode will be used to identify a more direct association between navigational search and a DPE task that taps dual mechanisms of cognitive control.
Experiment 2
First, we aimed to replicate the findings of Experiment 1 using an identical navigational search task. To provide more direct evidence for the involvement of proactive and reactive control in this task, we hypothesise that individuals with a bias for engaging reactive control mode would make more revisits, compared with their peers who are biased to engage proactive control mode to maintain the rule and update working memory. Although individual bias for a dominant control mode was not established for participants in Experiment 1, one could posit a general bias for proactive control mode is most likely for these healthy young adults (18–30 years) compared with younger children and slightly older healthy adults (40–45 years) who perform more in reactive mode (Chatham et al., 2009; Janowich & Cavanagh, 2018). Both modes of control were evident from the rule-congruence of button presses and the strong bias for revisits under the flashing rule. For individuals with dominant proactive control, their bias for maintaining the rule predicts a stronger tendency for more button presses to rule-congruent locations, compared with reactive control mode.
Exploratory analysis of revisits in relation to rule-adherence and distraction by flashing locations will be compared for individuals who vary in their bias for proactive and reactive control. Where proactive control is dominant, revisits were hypothesised to be more likely at rule-congruent locations in accordance with maintenance of the probabilistic rule, implying no strong bias for either rule or location. If reactive control mode is dominant, revisits will be less sensitive to rule-adherence (with diminished proactive control) but more allocation of resources to perceptual monitoring of the display. Together, these factors imply that reactive control is more likely to generate conflict between orienting to the more salient (flashing) locations, and re-instating the search goal, leading to more search errors at the flashing locations compared with individuals in proactive control mode.
Method
Participants
A total of 47 participants (9 males, 38 females) were recruited as undergraduate volunteers, with participation in lieu of course credits. All participants gave informed written consent and were debriefed about the experiment, and the study was approved by a local ethics committee. All participants self-reported no hearing, or cardiovascular problems and an average level of fitness.
Task design and materials
Automated navigational search paradigm
The search task was identical to Experiment 1.
Mouse-tracking variant of DPE task
For the DPE task, a novel mouse-tracking variant was developed using MouseTracker Analyzer™ software (Freeman, 2018; Freeman & Ambady, 2010; Freeman et al., 2011) to give a dynamic component to responses to the visual stimuli while drawing on visuo-spatial processing and long-term memory: cognitive abilities that also contribute to navigational search behaviour. On each trial, a visual-spatial (A or B) cue preceded a visual-spatial (X or Y) probe. Four trial-types (A-X, A-Y, B-X, B-Y) were included to reflect different cue-probe combinations and presented in the following distribution (A-X, 60%; A-Y, 15%; B-X, 15%; and B-Y, 10%). In total, 10 practice trials preceded 40 trials presented in a randomised order. The A cues consisted of a vertical row of three filled black circles. Two B-cue variants were used with either four or five white circles. The X-probes consisted of three circles (two blue, one white). The Y-probes comprised two rotated variants of X-probes (see Figure 3).
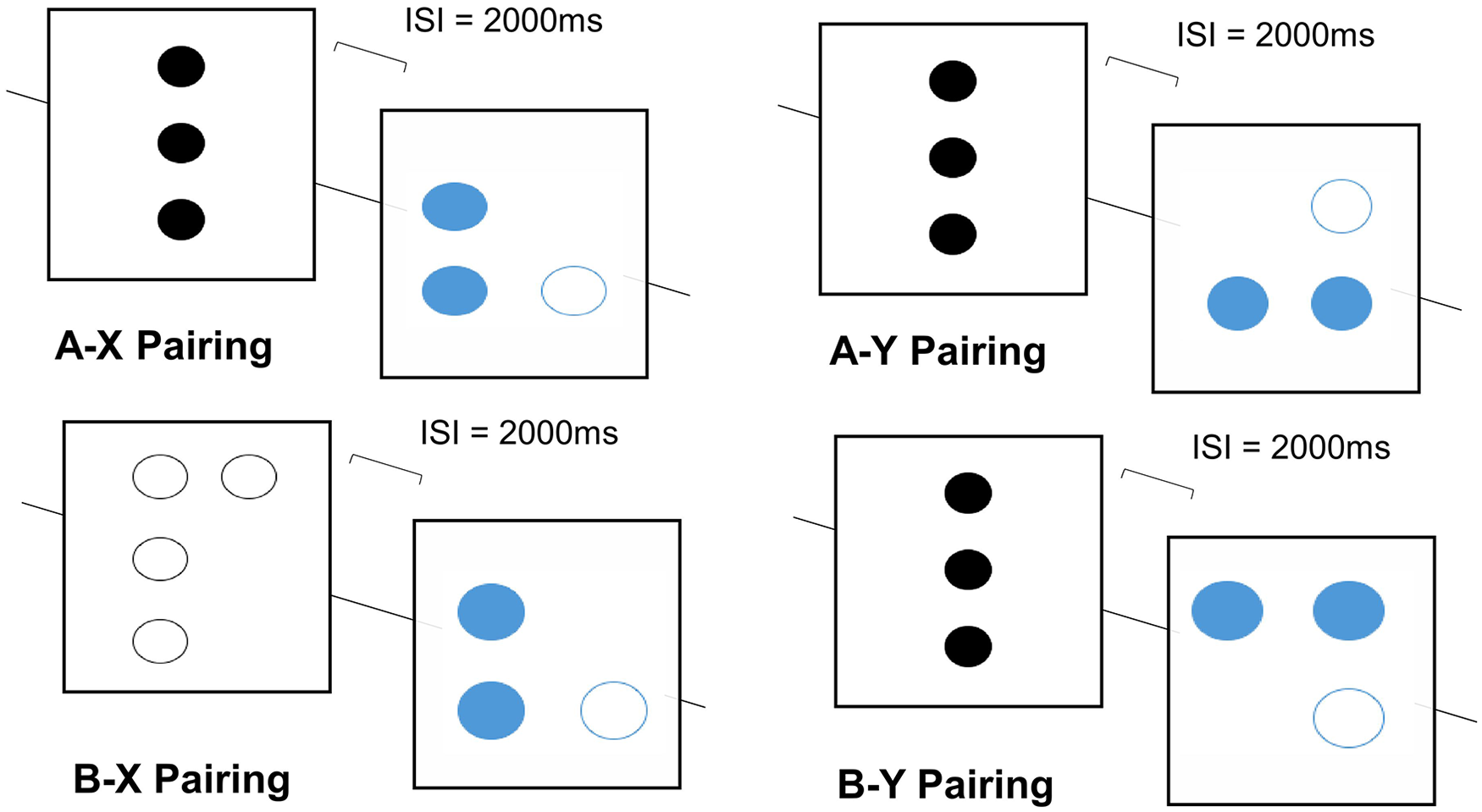
Example trials for four cue-probe associations, where A and B cues (and also X-probes) represented by one single dot array, but Y probes represented by two discrete dot patterns (shown here in A-Y and B-Y pairs) that occurred equally often with A and B cues. Intervals between pairings were 1,000 ms, with a 6,000-ms response window.
For each trial, dynamic motion of a hand-held computer mouse from an onscreen “start button” towards a Yes/No target box at the top right and left corners of the screen were recorded. Although mouse trajectories can give insights to the evolving dynamic competition during decision-making (Fischer & Hartmann, 2014; Spivey & Dale, 2006); here, errors and response latencies are reported and used to derive individual preferences for proactive or reactive control.
Procedure
Participants attended a single test session of approximately 50 min that was split between a room with dedicated search apparatus and an adjoining laboratory. For all participants, the DPE task was administered, followed by the automated navigational search task. Each DPE trial started with a 300-ms cue stimuli (A or B), followed by a 2,000-ms delay interval, prior to presentation of a 300-ms probe stimuli (X or Y). Participants were instructed to respond “Yes” to a “correct” A-X pairing, and that they should generate a “No” response to any other stimuli pairings (as an “incorrect” pairing). Participants were told to respond as quickly and accurately as possible by clicking the onscreen (START) button centred in the lower middle of the screen that released the mouse for data recording and then clicking their response at one of two locations in the top left- and right-hand corners of the display. The response window was set to 6,000 ms, after which participants received a time-out reminder to start moving earlier, even if they were unsure of their response.
Data handling
As in Experiment 1, data from the navigational search task were examined with ANOVA using JASP (JASP Team, 2018). For the DPE task, errors and response latencies were obtained from analysis conducted within MouseTracker Analyzer software (Freeman & Ambady, 2010; Freeman et al., 2011). Due to data recording errors, navigational search data from one participant and three from the DPE task data were unavailable.
Results
Navigational search
Similar to Experiment 1, the proportion of correct button presses was explored under two manipulations; by rules (A and B; 60% likely under static and flashing locations respectively) and by salience of the target location (either flashing or static locations). Figure 4 illustrates a search bias in the proportion of button presses made consistent with both rules.
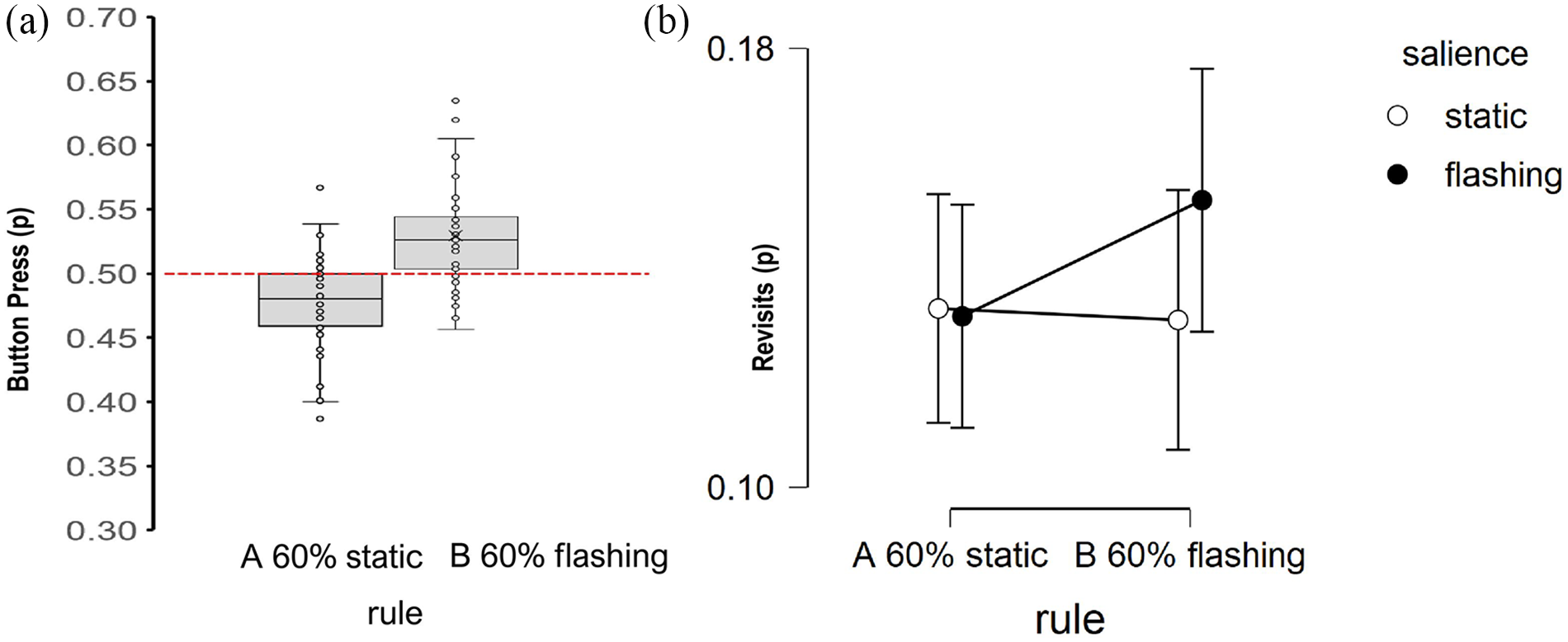
The average proportions of (a) button presses and (b) revisits generated under a probabilistic rule for locating targets at static (Rule A) and flashing locations (Rule B). (a) Boxplots for flashing locations only due to non-independence. (b) Revisits as a function of the salience of search locations.
Consistent with the findings of Experiment 1, Figure 4a shows button presses were made to flashing locations that aligned with, or against, the flashing and static rules respectively. To test for rule-congruent navigation, one-way comparisons were conducted for button presses at flashing locations under each rule. For the flashing rule B, more button presses were made to flashing locations, t0.5(45) = 4.95, p < .001, Cohen’s d = .73, 95% CI = [0.40, 1.05], and for the static Rule A, fewer button presses were made to flashing locations than expected by chance, t0.5(45) = −4.01, p < .001, Cohen’s d = −.59, 95% CI = [−0.90, −0.27]. Consistent with Experiment 1, there was strong evidence for rule-adherence, consistent with proactive control mode.
Figure 4b plots the effect of both salience and rule on the proportion of revisits. A 2 (rule type) × 2 (salience of target location) ANOVA (Type III) identified no main effect of either rule type, Rule A versus Rule B, F(1, 45) = 0.31, p = .58,
Dual-mechanisms control on the DPE task
For the DPE task the proportional error rates was low across all trial types (MAX = 0.01, MAY = 0.02, MBX = 0.05, MBY = 0.01) and so were not analysed further. Response latencies (within 3 SD of the condition mean) were slower than typically reported on expectancy tasks (~1,300 ms). Responses to AX trials (MAX = 1240, SDAX = 176) appeared faster than latencies for non-target trials (MAX = 1372, SDAX = 249; MBX = 1289, SDBX = 226; MBX = 1341, SDBY = 284) consistent with the strong expectancy established for the AX targets. Critically, a planned comparison of latencies for the AX-AY contrast favoured longer latencies for the AY trials; tAX-AX(43) = 4.2, p < .001, d = .63, that implicated interference from proactive control, despite the need to engage reactive control for successful resolution of late-stage conflict between the AX expectancy and the Y probe (Braver et al., 2009; Janowich & Cavanagh, 2018). Similarly, for the BX trials, the B cue invokes proactive control for the preparation of a non-target response, but where the late-onset of the X-probe creates conflict, and therefore interference from reactive control induces more error and slower latencies relative to AX and BY trials. Response latencies on BX trials were significantly slower than AX trials, tBX-AX(43) = 1.74, p < .05, d = .26, but not compared with the latencies for BY trials, tBX-BX(43) = −1.3, p = .89, d = −.19, giving only anecdotal evidence for slowing on the BX trials. Since the overall pattern of slowing was consistent across the exception (non-target) trials, the critical contrast of AY-BX latencies was used to capture the balance between proactive and reactive control modes within individuals using the Behavioural Shift Index (Braver et al., 2009; Janowich & Cavanagh, 2018), calculated as (MAX – MAY)/(MAX + MAY) from response latencies.
Individual variation in control mode and rule-adherence in search behaviour
Participants were split into two subgroups with a bias for proactive (n = 27) and reactive (n = 16) control modes in the DPE task, using the Behavioural Shift Index to determine whether a dominant control mode predicted variation in the navigational search task. Figure 5 plots the proportion of button presses to flashing and static locations, for both subgroups, for the corresponding rules. In both subgroups, there was a general bias to search at locations congruent with the rule.
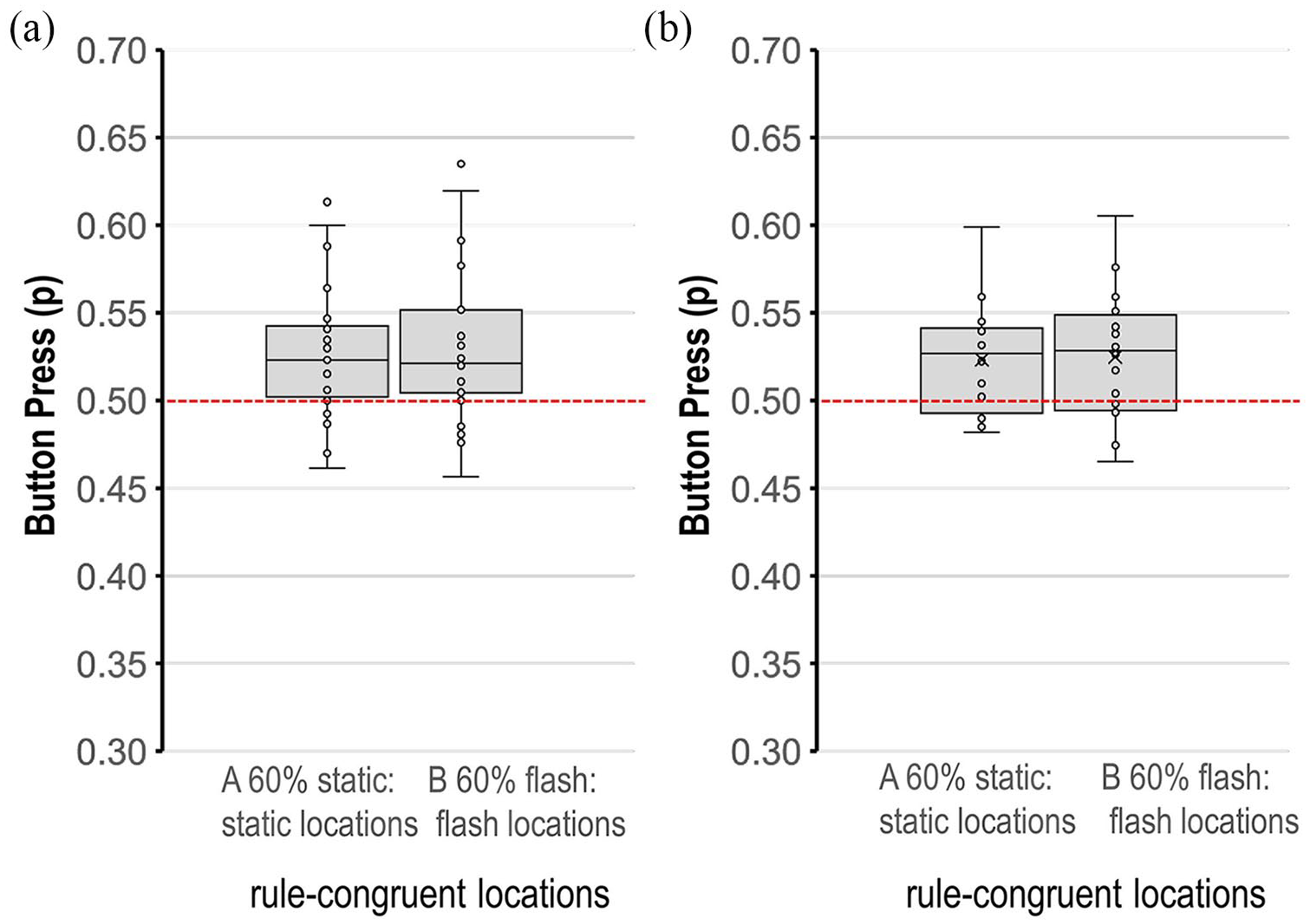
Boxplots of the proportion of rule-congruent button presses to flashing locations (as 60% more likely under the flashing Rule B and to static locations (as 60% more likely under the static Rule A) for the (a) proactive and (b) reactive subgroups.
As shown in Figure 5a, under the flashing Rule B, both subgroups showed a preference for flashing locations (proactive M = 0.53, SD = 0.04; reactive M = 0.52, SD = 0.04) with little evidence of a group difference, t = 0.52, p = .60, d = .16. As shown in Figure 5b, under the static Rule A, both subgroups showed a preference for static locations (proactive M = 0.48, SD = 0.04; reactive M = 0.48, SD = 0.03) with no evidence of a group difference, t = −0.19, p = .88, d = −.06. A general tendency for rule-congruence was evident in both subgroups.
Individual variation in control mode and revisits as search errors
For the revisits, the two subgroups appeared to vary in their sensitivity to the a-priori rule, with more revisits in the proactive subgroup under the flashing rule, as shown in Figure 6.
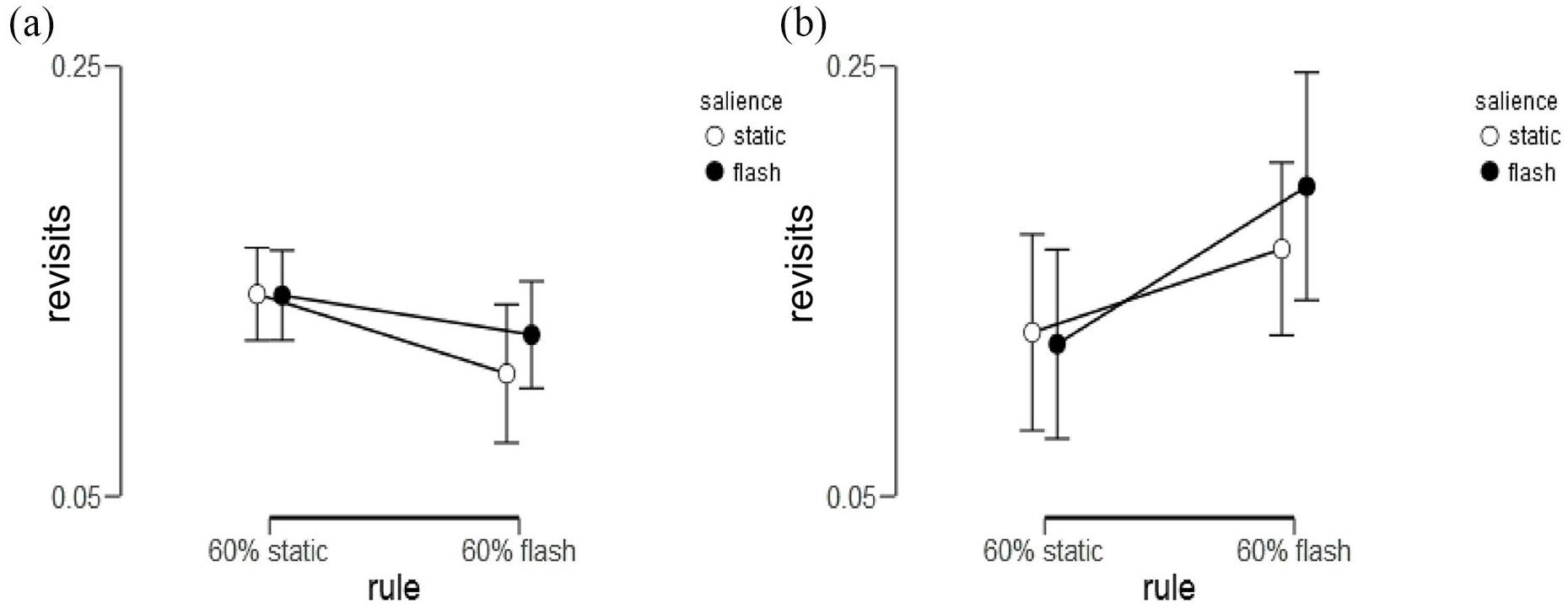
Displays revisits as a function of the rule and salience of search locations for the (a) proactive and (b) reactive subgroups.
Figure 6b displays a trend for more revisits under the flashing rule for the reactive group, compared with the proactive group (6a). In a 2 (group) × 2 (rule) × 2 (salience of target locations) ANOVA (Type III) for group (Reactive vs. Proactive) by rule (A or B) and salience (Flashing vs. Static locations) on the proportion of revisits, the groups were matched for the amount of revisits overall, F(1, 41) > 1, p = .36,
Summary of findings
In this replication of Experiment 1, the effects of the rule manipulation on button presses in the automated navigational search paradigm reflected a strong tendency for rule-adherence. That is, the button presses were more likely at static locations under the static rule and more button presses to flashing locations occurred under the flashing rule. A slightly different pattern was observed for revisits in Experiment 2 compared with Experiment 1. Revisits were more likely only to (rule-congruent) flashing locations under the flashing rule in Experiment 2. This did not replicate the findings of Experiment 1, where revisits were more likely under the flashing rule (and to flashing locations overall) but were not biased only to rule-congruent locations.
Experiment 2 further sought to provide a clearer association between the dual-mechanisms account of cognitive control and navigational search, by focussing on individual bias for proactive and reactive control modes. The novel variant of the DPE task captured dual-mechanisms of control embedded in motor behaviour and was used as a basis for relating individual differences in dual-mechanisms of control to navigational search behaviour. In the context of monitoring a highly frequent response schedule for the target A-X pairs, proactive control was captured by longer latencies on the A-Y trials, relative to A-X trials, as indicative of conflict between the anticipatory maintenance of a target response (elicited by an A-cue) and reactive control of responses towards a non-target (to successfully respond to the Y-probe). Some slowing on the BX trials was evident relative to the A-X pairs, with more atypical slowing on the BY trials than was anticipated. The Behavoural Shift Index was used to estimate individual bias using the contrast of AY-BX trial latencies to determine the balance of control modes. Typical of the undergraduate population sampled, more participants were characterised by proactive control mode than reactive control mode. Group sizes were sufficient to compare these groups for distinct patterns of navigational search behaviour. Participants in both reactive and proactive control modes demonstrated rule-adherence so that button presses were more likely to occur in locations that corresponded to an a-priori rule. Contrary to the prediction that the proactive subgroup would show a stronger bias for rule-adherence, the degree of rule-adherence was well-matched across subgroups. Interestingly, the reactive subgroup did not engage simply with pressing buttons at the more salient (flashing locations) in the display, rather, they too showed rule-adherence when responding with button presses at static locations that were less salient under the corresponding rule.
A final prediction was that the groups would vary in the pattern of revisits with the proactive group generating search errors at rule-congruent locations in accordance with maintenance of the probabilistic rule. Interestingly, the proactive group responded to the revisiting of search locations in a relatively unbiased way, suggesting that they were less sensitive to features of the display that were more salient. The reactive group showed greater susceptibility to revisits under the flashing rule where a salient feature of the display converged with the implementation of the (flashing) rule. Search errors were distributed across both static and flashing locations; only the implementation of the flashing rule influenced the proportional increase in behaviour towards flashing locations for individuals in reactive control mode. Across two experiments, the labile distribution of revisits in relation to the rule and perceptual aspects of the display points to the importance of individual differences. Notably, the sensitivity of search errors to the imposition of the flashing rule in the reactive group corresponded closely to the pattern of revisits seen in Experiment 1. This suggested a high proportion of individuals in reactive control mode were sampled in the first experiment.
General discussion
Earlier studies of navigational search have proposed a role for cognitive control in the search behaviour of typical adults (Longstaffe et al., 2014) and children with autism (Pellicano et al., 2011). In the present experiments, participants were given an a-priori rule at the start of each block of trials. Registering and maintaining a probabilistic rule requires allocation of working memory resources prior to, and potentially during, navigational search. That is, the rule could be maintained before, and re-instated after, each search event, to guide search behaviour. An active search event in the task means that participants stop, reach down and press the button to determine whether that location reveals the target or not. It is likely that the combination of “stopping to search,” updating of any representation of the spatial array for non-target locations and the re-instatement of the goal will engage cognitive control. Revisits are search errors to previously visited locations, presumably as a consequence of a failure of sufficient “tagging” of visited locations in working memory for the spatial array (see also Longstaffe et al., 2014). The question is how and when does navigational search require cognitive resources, especially in this paradigm, that combines the knowledge of a probabilistic rule with varying perceptual salience of the search environment.
Simple explanations of search behaviour that are determined either by the rule or by the perceptual salience of the display can be ruled out. That is, search was not driven only by the perceptual salience of locations that commanded attention, and so directed search behaviour to those locations or by the rule alone. In two experiments, button presses were biased towards the search locations congruent with the rule, rather than features of the display per se. Some evidence from Experiment 1 suggested the flashing locations were revisited more often as search errors, indicating some search bias towards those locations, however this was not observed in Experiment 2. Critically, revisits were more likely to be influenced by an a-priori rule to search flashing locations. This finding partly concurred with Longstaffe et al. (2014) where more search error (indexed by revisits) occurred when there was uncertainty about which locations had been previously visited (i.e., when lights did not extinguish after a button press). Since the tendency to make search errors was stronger in relation to the (flashing) rule, rather than location, the origin of search errors was likely to stem from cognitive load for updating working memory at each search event not simply a perceptual salience bias towards the flashing locations.
In Longstaffe’s account of revisits drawn from perceptual load theory of attention (Lavie, 2005, 2010), revisits occur from the need to “tag” previously visited locations in working memory that competes for a finite amount of cognitive resources with the perceptual load generated by (more salient) flashing locations. Extending Longstaffe’s account to consider the rule-congruence of search behaviour generates a different hypothesis. That is, if participants engaged with the flashing rule, searching task-relevant locations should be more exclusive towards flashing than distractor (static) locations, reducing the cognitive load of search due to the congruence between rule and perceptually salient features of the display. Conversely, if participants engaged with the static rule, searching task-relevant locations should be more exclusive to static than distractor (flashing) locations. Here, the cognitive load of search is higher given the perceptual salience of the distractor locations. This view predicts that search errors would be more likely under conditions of high cognitive load, due to the finite recruitment of attentional resources. Therefore, more revisits would be induced by the static (not flashing) rule. Contrary to this extension of Longstaffe’s account, both experiments reported here indicated that the flashing rule led to more revisits, suggesting a complex interplay of perceptual salience with the demands for cognitive control of resource allocation.
Dual mechanisms of cognitive control that distinguish mechanisms of control at different time-points, provide a stronger account of navigational search data than previously advanced by Longstaffe et al. (2014). Proactive control is a control mechanism which would maintain the probabilistic search rule (goal) in working memory, by regulating conflict from the stimuli-driven aspects of the display and the selection from automatic action schemas that enable multiple locations to be visited. In both experiments, search was consistent with the anticipatory maintenance of the probabilistic rule that biased the proportion of button presses made to rule-congruent locations. This provided indirect evidence of proactive control of navigational search. Reactive control mode co-ordinates attentional and cognitive mechanisms to re-orient action and perception systems (post-event) towards the goal or stimuli context. In the navigational search paradigm, the search event offers new information regarding the goal of finding the target, requiring cognitive resources to be allocated to updating spatial memory and so guide a new action schema that will achieve the search goal. Search errors, as revisits to previously visited locations give insight to the failure of control process, either in the updating of spatial memory or in the re-instatement of the search goal. As each search location is checked, reactive control is mobilised to provide a mechanism for resolving conflict between multiple action schemas and the stimulus-driven updating of the search environment to re-instate the goal.
The novel variant of the DPE task captured dynamic attributes of control mechanisms embedded in motor behaviour, even over a relatively small number of trials. Both the mouse-tracking variant of the DPE task and the search paradigm provided complex environments for monitoring and generating a motor response to spatial aspects of an environment, in addition to the demands of updating and regulating behaviour in the event of new information. Latency data in critical interference trials were sufficiently stable to identify modes of cognitive control in two subgroups. Including the DPE task therefore allowed a more direct evaluation of search behaviour for individuals who were predominantly in proactive and reactive control mode. For button presses, there was no evidence that control mode biased search for the proactive group through a stronger anticipatory tendency towards rule-congruent search behaviour. This could imply that instating the rule at the outset, rather than anticipatory maintenance of the rule was sufficient to bias button presses to rule-congruent locations.
Mechanisms of control should be evident in search errors, as seen in the pattern of revisits in relation to the dominant control mode. If proactive control mode is dominant, we predicted that revisits would be biased towards rule-congruent locations since new action schemas are governed by anticipation of the likely location of the goal. Since reactive control is diminished in this mode, the search errors are more likely to correspond to rules with less sensitivity to the task context. The findings were consistent with this prediction, as resilience to search errors were relatively unbiased. In this mode, maintaining and reinstating the relevant rule after each search event, without undue distraction from the search environment, characterised performance. Where reactive control mode is dominant, then we predicted search errors will be less sensitive to rule-adherence as new information is updated from the search event and with close perceptual monitoring of the display. In Experiment 2, there were more revisits to rule-congruent (flashing) locations made by participants in reactive control mode implying that closer monitoring of perceptually salient flashing locations led to more search errors.
In these experiments, the search task varied the perceptual salience of flashing and static locations with a consequence for resource allocation (see also Longstaffe et al., 2014). Although there was some evidence that flashing locations received more revisits overall than static locations in Experiment 1, the effect was relatively small, compared to the sensitivity to the flashing rule. We propose that the greater distraction from monitoring flashing locations engages the automatic orientation of reactive control towards the stimulus display. By allocating attention to the perceptual salience of the display guided by the flashing rule, potentially in combination with weaker updating of working memory for the search array, more conflict is generated between action schemas that are consistent with the flashing rule, including those for previously visited locations. The combination of more conflict between action schemas and diminished proactive control that reinstates the search goal for a target at a new location, these setting conditions led these participants in reactive control mode to make more revisits to flashing locations. Previous evidence suggests that proactive mechanisms are closely associated with the recruitment of working memory resources (Redick, 2014). Therefore, under proactive control mode, participants were more adept at recruiting and updating working memory to support search behaviour, possibly by creating more opportunity for updating the “tagged” locations in working memory. Participants in proactive control mode were less attentive to the local features of the display due to diminished reactive control and generated fewer search errors linked to the salience of flashing locations.
In conclusion, the evidence suggests that navigational search operates through the involvement of cognitive control processes, as could be expected for an energetic and costly exercise that co-ordinates cognitive resources for an optimal behavioural outcome. The dual-mechanisms framework accounts for the complex pattern of data, as well as providing a route to the study of individual bias in cognitive control modes. In the present study, the categorisation of participants by dominant control mode reflected an intra-individual bias that was generated from the task context, rather than being a stable factor derived from multiple measures or from repeat testing. The sample was characteristic of a typical population of young adults, with a dominant mode of proactive control. Rather than a stable trait, dominant mechanisms of control can fluctuate over short and immediate time-frames, consistent with life skills and training (Arbula et al., 2016; Chang et al., 2018). Indeed, intra-individual biases in proactive control can altered by training, for example, with a cognitive intervention over several weeks (e.g., Li et al., 2018). Future work should ascertain how shifts between the two control modes occur over short and medium time-frames, perhaps due to extrinsic factors that alter goal-maintenance or from the meta-cognitive regulation of resource allocation (e.g., Chevalier et al., 2015). As some classes of navigational search do benefit from training (e.g., Riggs et al., 2018), dual mechanisms of control can help explain how extrinsic and intrinsic goals can be recruited to serve goal-directed behaviour within complex search. More insight to how and when mechanisms of cognitive control are deployed during navigational search could offer valuable support to people with impaired cognitive function, especially for simple, everyday challenges, such as finding one’s keys before leaving the house.
Footnotes
Acknowledgements
The authors would like to thank Shweta Maheshwari, Amelia Frett, and Emma Mackey for help with data collection. They would also like to thank the editor and two reviewers of this manuscript for constructive and insightful comments on an earlier version.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.