
Abstract
A recently published article by van Heijst et al. attempted to reconcile two research approaches in the science of emotion—basic emotion theory and the theory of constructed emotion—by suggesting that the former explains emotions as bioregulatory states of the body whereas the latter explains feelings that arise from those state changes. This bifurcation of emotion into objective physical states and subjective feelings involves three misleading simplifications that fundamentally misrepresent the theory of constructed emotion and prevent progress in the science of emotion. In this article we identify these misleading simplifications and the resulting factual errors, empirical oversights, and evolutionary oversimplifications. We then discuss why such errors will continue to arise until scientists realize that the two theories are intrinsically irreconcilable. They rest on incommensurate assumptions and require different methods of evaluation. Only by directly considering these differences will these research silos in the science of emotion finally dissolve, speeding the accumulation of trustworthy scientific knowledge about emotion that is usable in the real world.
van Heijst et al. (2025) recently attempted to integrate basic emotion theory (BET) with the theory of constructed emotion (TCE). Their central claim was that BET explains emotions proper, whereas the TCE explains feelings. In presenting this contrast, van Heijst et al. defined emotions as “bioregulatory states of the body” (p. 384) and feelings as “the mental representation of the physiological changes that occur during an emotion” (p. 384). Separating an instance of emotion into an objective, physical state and the felt experience of that state is not a novel maneuver, however, having been suggested by others who have attempted to understand and integrate the two theories (e.g., Adolphs, 2017; Damasio, 2004; de Waal, 2019). We—the primary architects of the TCE—have repeatedly noted that this bifurcation, although consistent with the assumptions of BET, fundamentally misrepresents the TCE (e.g., Adolphs et al., 2019; Barrett, 2015, 2017b; Barrett & Lida, 2024). This maneuver ignores several key hypotheses (which we refer to as Error 1) and much of the multidisciplinary evidence that supports the viability of those hypotheses (which we refer to as Error 2). To characterize the magnitude of the problem, we offer an analogy. Most people would not stir a spoonful of ketchup into their coffee. The two fundamentally do not go together. But you can force them to go together if you strip out most ingredients from the ketchup and leave only the sugar, which mixes with coffee just fine. Likewise, van Heijst et al. stripped away most of the TCE and then attempted to mix the remains into BET. Our analogy, although silly, illustrates the problem of oversimplifying something by ignoring its key elements.
van Heijst et al. compounded their two errors of omission with a third involving their evolutionary analysis. They offered an evolutionary account of emotions as evolved phenotypic adaptations, which is an assumption of BET. Their analysis was structured using the four questions proposed by the ethologist Niko Tinbergen (Tinbergen, 1963/2005). Tinbergen fashioned his questions as a recipe, the answers to which would provide a complete biological account of any behavior. van Heijst et al. suggested that the two emotion theories attempt to answer different questions posed by Tinbergen and therefore aim themselves at emotions and feelings as different phenomena, consistent with this logic. Unfortunately, this analysis sidestepped major scientific advances in evolutionary biology since Tinbergen articulated the four questions. These advances are wholly inconsistent with BET but are key to understanding the evolutionary foundation of the TCE.
Here we unpack van Heijst et al.’s three misleading simplifications and the resulting errors. Our goal here is not to provide a comprehensive treatment of our theory or of the science of emotion in general. Instead, we begin by showing that van Heijst et al. ignored key hypotheses in the TCE to achieve their proposed bifurcation of emotion (as objective physical states) from feeling (as subjective experience). This simplification by omission extends to their treatment of the published empirical evidence, which we comment on next. We then explain the errors of omission in their evolutionary analysis. We end by pointing out that van Heijst et al.’s oversimplifications and errors are not unique—they have been published numerous times by other authors, resulting in an unfortunate tradition of mischaracterizing the TCE. This situation arises from a failure to recognize the incommensurate assumptions of the two theories, creating a serious obstacle to reconciling existing research silos in the science of emotion. To present evidence of this obstacle, we end with an error in the science of emotion that has persisted for more than a century, born from an integration of two other theories that held incommensurate assumptions about the nature of emotion.
Error 1: The Theory of Constructed Emotion Is More Than a Theory of Feeling
Table 1 compares our formulation of the TCE, van Heijst et al.’s oversimplified depictions of the theory, and their descriptions of BET, sorted and organized by Tinbergen’s four questions. Our theory, when properly articulated, constitutes hypotheses about emotions as “bioregulatory responses” in the service of survival (to paraphrase their wording). Emotions are not special in this regard, however. Every moment of life involves the brain’s bioregulation of the body. A brain’s role in coordinating movement broadly defined (to include the movements of the viscera, of skeletomotor muscles, of chemicals within the blood, etc.) is central to our theory. Furthermore, in our theory, feeling cannot be meaningfully separated in any biologically justified manner from the brain’s regulation and coordination of the various systems of the body. We formulated these hypotheses on the basis of decades of empirical evidence from other disciplines—evidence that is fundamentally about how biological and other complex systems work. Much of this evidence is often overlooked by some scientists who search for basic emotions.
The TCE and BET Through the Lens of Tinbergen’s Four Questions
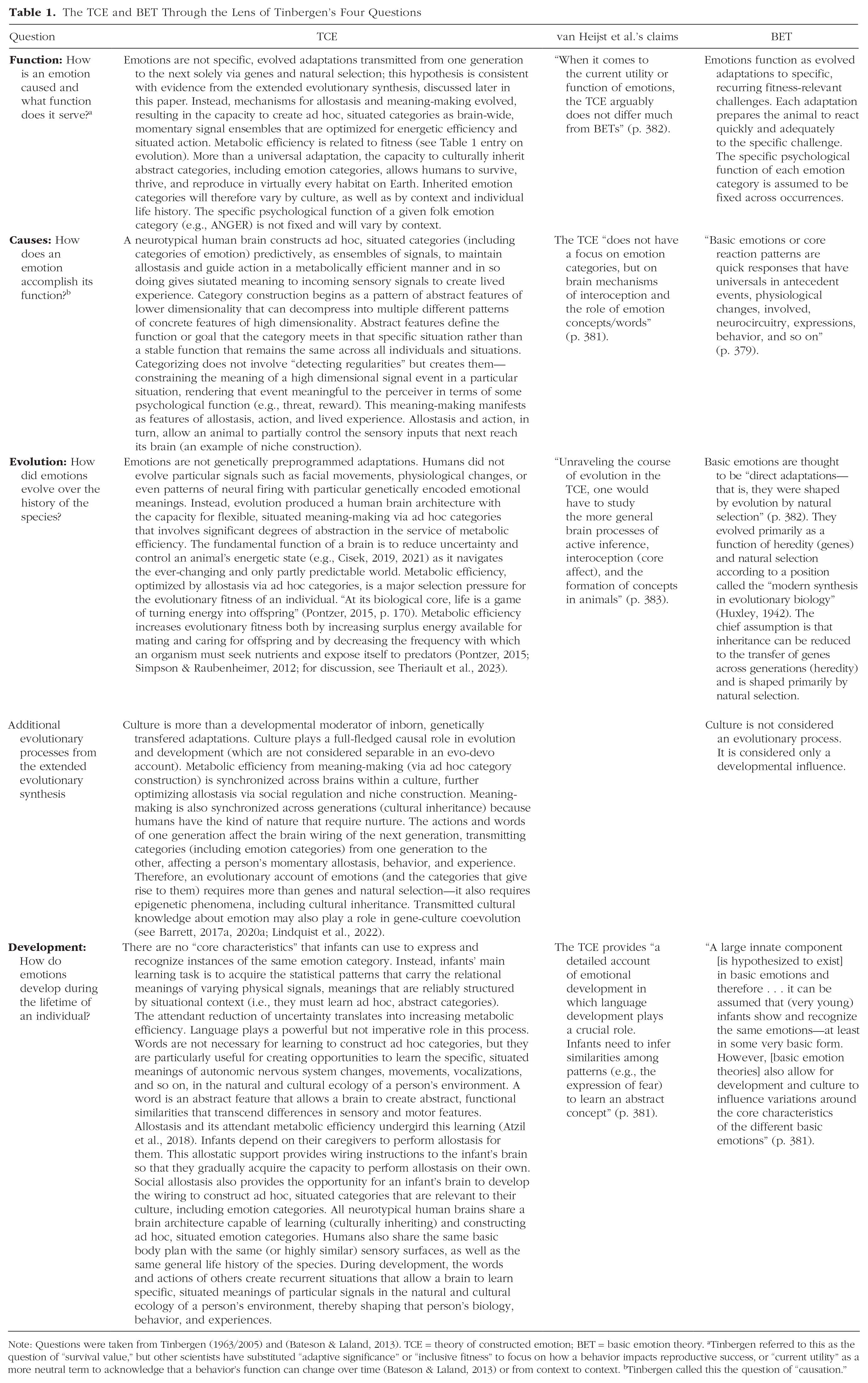
Note: Questions were taken from Tinbergen (1963/2005) and (Bateson & Laland, 2013). TCE = theory of constructed emotion; BET = basic emotion theory. aTinbergen referred to this as the question of “survival value,” but other scientists have substituted “adaptive significance” or “inclusive fitness” to focus on how a behavior impacts reproductive success, or “current utility” as a more neutral term to acknowledge that a behavior’s function can change over time (Bateson & Laland, 2013) or from context to context. bTinbergen called this the question of “causation.”
The theory of constructed emotion in brief: Bioregulation and feeling cannot be separated in a biologically meaningful way
The TCE is an integrated collection of hypotheses describing how a brain regulates the systems of its body, controls action, and in the process constructs the mental events that constitute a mind, including (but not specific to) occurrences of emotion. This focus on the role of the brain in regulating the body, as well as the direct role of this regulation in constructing lived experience, makes our theory of emotion different from other constructionist theories that have been in the published literature (sometimes in nascent form) at least since the mid-1800s (Gendron & Barrett, 2009). In addition to this emphasis on biology and physiology, our evolutionary hypotheses (discussed later) likewise distinguish the TCE from other constructionist theories, meaning that a discussion of our theory cannot stand in for a more general discussion of the constructionist research tradition. Therefore, when van Heijst et al. stated that a “constructionist approach to emotion has existed for well over a century and the debate between BETs and the TCE is by no means a recent development” (p. 378), they overlooked the ways in which our theory of emotion contains distinctive biological, physiological, and evolutionary features. The first article on the TCE was published less than 20 years ago. A brief history of the theory’s development and its relation to other constructionist theories can be found in Barrett and Lida (2024).
The TCE is transdisciplinary—it integrates evidence and concepts from a range of scientific disciplines beyond psychology, including biochemistry, physiology, evolutionary and developmental neurobiology, ethology, neuroanatomy and neuroscience, anthropology, and linguistics, with additional conceptual tools from philosophy, history, and engineering. The resulting hypotheses transcend the boundaries of any single discipline. The transdisciplinary nature of our hypotheses makes the theory’s full scientific potential difficult to summarize in simple sentences.
To date, we have published more than 20 theoretical works that lay out the conceptual foundations of the theory in more detail, review the multidisciplinary evidence that justifies our hypotheses, and discuss empirical strategies for their proper evaluation (e.g., Atzil & Gendron, 2017; Atzil et al., 2018; Barrett, 2012, 2017b, 2020a, 2022; Barrett & Finlay, 2018; Barrett & Lida, 2024; Barrett & Satpute, 2019; Barrett & Theriault, in press; Barrett & Westlin, 2021; Barrett et al., 2019; Bliss-Moreau, 2017; Chanes & Barrett, 2016; Gendron & Barrett, 2018; Hoemann et al., 2017; Hoemann, Wu, et al., 2020; Hoemann, Xu, & Barrett, 2019; Lindquist, 2013; Lindquist et al., 2015, 2022; Shaffer et al., 2022). Our hypotheses are aimed at understanding occurrences of emotion as whole-brain events that coordinate and regulate the internal operations of the body in addition to coordinating and controlling skeletomotor movements (i.e., control “bioregulatory states of the body”) while at the same time creating lived experience in a physical and social world (including but not limited to “feeling”). The fact that we study whole-brain events means that the brain’s regulation of the body and its construction of feeling cannot be bifurcated into distinct phenomena caused by separate mechanisms. van Heijst et al. missed this key implication in their treatment of our theory. In addition, several published meta-analyses and many empirical works that support the TCE focused on the brain’s regulation of the body and not directly on feeling. A partial list of these publications can be found in Table 2. Additional empirical evidence can be found in Table 1 of Barrett (2012) and in Barrett (2017b). van Heijst et al. did not discuss this evidence or its relevance to their arguments.
Sample of Published Studies Supporting the Theory of Constructed Emotion’s Population Thinking (and Disconfirming Basic Emotion Theory’s Typological Thinking)

A brain regulates and coordinates the systems of its body
The TCE hypothesizes that a brain’s main function is to anticipate the needs of the body, prepare to meet those needs before they arise, and, in so doing, control and coordinate action at the level of the whole body. This ongoing process, termed “allostasis” (Sterling, 2012; Sterling & Laughlin, 2015), includes the coordination of the visceral organs, immune system, and endocrine system, collectively referred to here as “allostatic control.” Allostatic control is necessary for and biologically intertwined with the control of skeletomotor action. This hypothesis rests on decades of empirical evidence in evolutionary biology, in neuroanatomy, on the metabolic costs of electrical and chemical signaling, and on the brain’s predictive capacities (e.g., see Barrett, 2017b; Gee, 2018; and Hutchinson & Barrett, 2019). van Heijst et al. mentioned the term “allostasis” in their treatment of the TCE but incorrectly defined it as “the process of maintaining and returning the body to homeostasis” (p. 378). Allostasis is not homeostasis. Homeostasis is reactive—its goal is to return a system to a set point. Allostasis is predictive and does not involve set points. The goal of allostasis is efficient energy regulation at all levels of metabolic output. Energy regulation can be efficient when you are asleep and when you go for a morning run. Individual systems may work homeostatically, but collectively they are being coordinated and regulated by the broader allostatic efforts of the brain. van Heijst et al. correctly acknowledged at the outset of their article that the TCE contains a hypothesis that “the primary function of the brain is to maintain allostasis and that emotions are consequences of this primary function” (p. 378), but they did not seem to realize the implications of this statement, for example, that allostasis does not cause separate instances of emotion but manifests them (as well as other brain events that psychological science assigns to other folk categories such as perception, cognition, decision-making, etc.).
Feeling is an emergent feature of the brain’s regulation of the body
On the basis of the aforementioned evidence from evolutionary biology, neuroanatomy, signal processing, and predictive processing, the TCE hypothesizes that efferent copies of the same allostatic signals that control and regulate the body simultaneously create feeling and other features of lived experience (in combination with afferent signals from the sensory surfaces of the body and reafferent signals from the thalamus; see Barrett, 2017b, Fig. 5; Hutchinson & Barrett, 2019, Fig. 1; Shaffer et al., 2022, Fig. 2). By efferent copies, we mean that the action potentials that traverse axons originating from primary allostatic control neurons in the brain (such as those in the anterior cingulate cortex) simultaneously send both motor control signals to the spinal cord and prediction signals that help construct the features of lived experience, including the mental features that describe feeling, perceptions, and sensations. This hypothesis is based on decades of anatomical evidence (e.g., Barbas, 2015). van Heijst et al. mentioned this hypothesis in their initial summary of our theory: “All mental events are a consequence of how the brain coordinates bodily regulation (allostasis) and plans action accordingly” (p. 378). They sidestepped this hypothesis and its implications, however, in their later oversimplification of our theory: “The TCE has focused on defining the brain circuitry and mechanisms that are at the basis of the formation of emotion concepts (and other mental events)” (p. 384). Clearly, the TCE contains hypotheses about the neural basis of felt experience, but another of its key hypotheses is that bodily regulation and feeling are biologically entwined (as are action and perception). It is impossible to bifurcate them into different phenomena in a way that is scientifically legitimate and justified.
Holism, not reductionism
Importantly, our theory’s hypotheses are holistic, not reductionist like BET. Holistically, an instance of emotion is understood as a complex causal ensemble (i.e., a whole-brain event) whose simultaneous features include bodily regulation and the mental features of experience. A reductionist approach, by contrast, understands an instance of emotion in terms of separate, simpler, mechanistic causes localized to different parts of the brain. Note that these two approaches are diametrically opposed. A reductionist approach understands the whole (an instance of emotion) in terms of its simpler causes. A holistic approach understands the causes in terms of the whole. van Heijst et al. did not acknowledge or grapple with this irreconcilable difference.
Our theory explicitly avoids the dualism inherent in reductionist approaches to emotion by explicitly defining “mental events” to include allostasis, skeletomotor actions, and the features of lived experience. Each event is thought to arise as complex relations among electrical and chemical signals. Some signals are intrinsic to the brain as a complex dynamical system, and some signals are inputs that continually arrive from the sensory surfaces of the body (e.g., the rods and cones in the retina of each eye, the cochlea in each ear, the olfactory receptors in the nose, the glucose receptors in the gut and muscle cells, the stretch receptors in muscle tendons, the aortic and carotid bodies that detect pH changes in the blood). These inputs arise in the body and in the world and constrain the intrinsic dynamics of the brain’s own signals. Conversely, intrinsic signals give meaning to incoming signals that they would not otherwise have, making them worthy of certain actions (or not) and the metabolic costs entailed. Psychologists describe this meaning in terms of mental features, such as pleasant, unpleasant, arousing, threatening, effortful, bright, dull, loud, urgent, and so on. These are the qualities of felt experience. They are not properties of the intrinsic brain signals alone or of the incoming sensory signals alone but of their relations (Barrett & Theriault, 2025). 1 We think of mental features as lower dimensional summaries of higher dimensional sensory and motor signals once their temporal and spatial redundancies have been removed (Barrett & Theriault, 2025). Mental features correspond to compressed, multimodal summaries of signals that arise from the brain’s ongoing bioregulation of the body in an ever-changing and only partly predictable world. van Heijst et al. either ignored or misunderstood this hypothesis in their analysis because they applied the reductionist causal logic of BET to the holistic TCE that explicitly eschews reductionism.
Bifurcation and the blind spot
In attempting to neatly separate emotions into bits that are objective, mind-independent physical states of the body versus bits that are subjective “mental representations” of those physical changes, van Heijst et al. committed an example of what philosophers of science have long called “the bifurcation of nature” (Whitehead, 1920). Perceiver-dependent phenomena, such as feelings, are assumed to exist only in a perceiver’s mind, and typically they are considered epiphenomenal to the objective phenomena that they represent. With this maneuver, van Heijst et al. followed in the footsteps of others who separated “an emotion” from the “experience of an emotion” as distinct phenomena (e.g., Adolphs, 2017; Damasio, 2004; de Waal, 2019). The implication is that features of lived experience, defined as representations computed in the brain, have no significant causal role in the brain’s bioregulation of the body and its outcomes.
The bifurcation of nature into “objective” and “subjective” categories is considered a philosophical blind spot (Byers, 2011; Frank et al., 2024) that has caused serious scientific problems in physics, biology, cognitive science (Frank et al., 2024), and psychological science, including the science of emotion (Barrett & Theriault, 2025). Notably, cleaving emotions as objective states from subjective feelings produced a century-old error in the science of emotion, as we discuss in the final section of this article. This bifurcation has also manufactured a “correspondence problem” in the science of emotion (e.g., Gross, 1998; Mauss et al., 2005; Ventura-Bort et al., 2022). The physical (and presumed objective) changes that occur during instances of a given emotion category rarely show consistent covariations with the mental features of experience during those instances (e.g., changes in heart rate or respiratory rate during instances of the folk category ANGER or FEAR do not correspond to self-reports of anger or fear). 2 Heart rate can increase, decrease, or remain the same (relative to a baseline measurement) when people report feeling angry or afraid (McVeigh et al., 2023; Siegel et al., 2018). A lot of ink has been spilled trying to resolve this dualism, without much success, probably because scientists have largely assumed that the problems are rooted in methodological problems rather than the theory-laden assumptions that ground those investigations in the first place.
More generally, Frank et al. (2024) suggested that the blind spot constitutes a scientific worldview with unexpectedly pernicious consequences. 3 In the science of emotion, for example, the blind spot contributes to the widespread failure of pharmaceutical companies to develop drugs that cure depression and anxiety. Scientists working in the basic emotion tradition routinely categorize freezing in nonhuman animals as a feature of the category FEAR and the assembly of neurons that help to control this behavior is categorized as a “fear circuit.” Other scientists—in the hopes of curing posttraumatic stress disorder or other anxiety disorders—focus their empirical efforts on discovering drugs that suppress this circuit on the assumption that all (or most) instances of FEAR issue from the same circuit. Physicians, and patients struggling with “fear-related disorders,” assume that the drugs in question will tame anxiety-related symptoms, but they do not work as expected because FEAR, like other folk emotion categories, is a collection of situated, variable instances that are caused by a variety of situated, complex ensembles of factors (as discussed in the next section). The result is widespread failure of research using animal models of FEAR or ANXIETY to develop medications for anxiety disorders, which led pharmaceutical companies to abandon further research and development (LeDoux, 2015).
The blind spot certainly contributes to the current assumption, found in the basic emotion tradition and in the industry, that emotional states can be read objectively in facial movements and physiological changes and can therefore be used to safely decide people’s employment and educational, medical, and judicial outcomes, despite considerable evidence to the contrary (see Andrews et al., 2024; Barrett, 2020b, 2022; Barrett et al., 2019; see also Frank et al., 2024, Chapter 7). This same manifestation of the blind spot also shapes judicial behavior within the U.S. legal system, in which defendants found guilty in capital cases received a death sentence rather than life in prison because they did not express remorse in a stereotypically American manner. A jury’s perception of whether or not a defendant is remorseful largely determines whether they recommend the death penalty (Lynch & Haney, 2011).
Error 2: Ignoring Empirical Evidence That Disconfirms Basic Emotion Theory
van Heijst et al. discussed empirical evidence that they took to be consistent with one emotion theory or the other. Unfortunately, they failed to mention much of the empirical evidence listed in Table 2 and elsewhere (Barrett, 2012, 2017b) that directly supports the TCE, as we already noted. Importantly, much of this evidence also directly disconfirms a key hypothesis of BET. In the words of van Heijst et al., “in BETs different basic emotions or core reaction patterns are quick responses that have universals in antecedent events, physiological changes, involved neurocircuitry, expressions, behavior, and so on (for an overview of BETs, see Tracy & Randles, 2011)” (p. 379). This hypothesis is an example of what evolutionary biologist Ernst Mayr called “typological thinking” (Mayr, 2004). Our goal in this article is not to provide a detailed review of the evidence that disconfirms typological thinking but to simply highlight that it exists because van Heijst et al. did not.
Typological thinking in basic emotion theory
Consider the folk category FEAR. It is a population of specific instances, each of which unfolds in a specific spatiotemporal context involving a specific individual. BET hypothesizes that these instances are a family of biologically prepared states that are functionally equivalent to one another by virtue of their family resemblance to a prototypic pattern of core features. Some variation within a category is permitted such that occurrences of FEAR may vary by context, culture, and species, but the occurrences are hypothesized to retain sufficient family resemblance to the prototype FEAR and sufficient difference from the prototypes of other categories. For example, BET would predict that there exists a family of FEAR expressions that share common configural properties (i.e., share a family resemblance) that also distinguish FEAR expressions from the family of ANGER expressions. If the occurrences create a distribution of FEAR patterns, then the prototype of this distribution, according to BET, should remain stable across time (i.e., is immutable).
In typological thinking, the summary of the distribution of a category’s instances—the category’s hypothesized prototype—is assumed to be real in nature. The observed variation in the distribution of instances is thought to reflect the intensity of the occurrence, regulatory efforts, or cultural norms in expressing emotion (Ekman 1992; Ekman & Cordaro, 2011), such as display rules or cultural dialects (e.g., Cordaro et al., 2018; Elfenbein, 2013; Levenson, 2011; Matsumoto, 1990), stochastic error (as suggested by Roseman, 2011), or methodological concerns (e.g., perhaps the stimulus situation was too weak to evoke prototypic emotions; see Levenson, 2011). The hypothesized prototype is thought to give instances of FEAR their unique identity and make them similar enough to be interchangeable yet different enough from the instances of other categories so as not to be confused with them; that is, FEAR is hypothesized to be a biological and psychological “type” (hence, “typological thinking”). And although instances of FEAR may vary somewhat in their features, they should share a family resemblance with the prototypic pattern that accurately predicts new instances of FEAR with high reliability and specificity. The presumed similarity among instances of the same category allows scientists to believe that they can curate and observe a small sample of FEAR instances (e.g., observing instances of FEAR in different participants who are placed in a specific situation), aggregate across those instances, and then confidently generalize their inferences to the larger population of FEAR instances, past, present, or future, regardless of context, individual, culture, and sometimes even species. Scientists working in the basic emotion tradition use the phrase “an emotion” to refer both to a single instance and a whole category of instances, consistent with this assumption. For example, the word “fear” refers both to an instance of FEAR and the entire category FEAR. This conflation is reasonable only if one assumes that each emotion category has an immutable prototype. Yet there has always been evidence of substantial within-category variation and across-category similarity that makes immutable prototypes unlikely. And as we discuss later, the TCE’s treatment of variation is very different from that of BET’s.
Evidence of structured variation disconfirms typological thinking
Typological thinking in the science of emotion is one endpoint of a scientific continuum. At the other end is population thinking, also named by Mayr (2004); see Figure 1. The TCE is rooted in population thinking (Barrett, 2017a; Hoemann, Khan et al., 2020; Le Mau et al., 2021; Siegel et al., 2018; Westlin et al., 2023). Variation among a category’s instances is hypothesized to be structured and meaningfully related to the situations in which those instances emerge. For example, the TCE would predict that the folk category FEAR contains substantially more variation than could be described by “a family of expressions that share common configural properties.” A person might cry in fear, narrow their eyes and shout in fear, smile in fear, or even laugh in fear. The variation is real and intrinsic to emotional meaning. The variation reflects a complex ensemble of spatial and temporal contextual factors, including what is occurring in the world and in the person’s own body as well as what just occurred and what is predicted to occur. Any overall summary of the population variation, such as an unchanging prototype, is a simplification that does not exist in the real world. This critical viewpoint directly contradicts BET and is fundamental to the TCE. One cannot integrate a theory that requires unchanging prototypes with a theory that denies their existence.

Typological and population thinking in the science of emotion arranged as a continuum. Typological thinking in the science of emotion assumes that each folk emotion category, such as FEAR, is a biological and/or psychological type (e.g., a genetically inherited adaptation with its own unique neural assembly, family of facial expressions and physiological patterning, and/or function). Accordingly, the features of any occurrence of FEAR have inherent emotional meaning. Instances of FEAR can vary somewhat from one another, but they are assumed to share a family resemblance to a stable set of prototypic features that define them as FEAR and not members of some other emotion category. Typological thinking uses reductionism as a scientific strategy. Phenomena, such as instances of FEAR, might vary from one another in their facial expressions, their physiological patterning, and so on, but it is assumed that this variation can be best understood in terms of simpler, more fundamental causes. Accordingly, typological thinking utilizes simple cause-and-effect mechanisms. Scientific studies are designed to find within-category similarities and between-category differences. Some variation within a category is permitted, as is overlap between categories (i.e., fuzzy boundaries), but the instances are assumed to retain sufficient family resemblance to their category prototype and sufficient difference from the prototypes of other categories. Context is assumed to moderate what is inherently biological; life experience, normal stochasticity, differences in the triggers for emotion, emotion-regulation efforts, and cultural norms for expressing emotion, such as, for example, display rules or cultural dialects, have all been proposed as moderators that tweak or otherwise add variation to universal expressive tendencies. Lived experience, such as being socialized within a given culture, is assumed to play no substantive role in the transition and evolution of emotion categories. Typological thinking assumes that the categories of reality, such as categories of emotion, exist independently of any human mind and are fixtures of reality to be discovered (i.e., emotion categories are perspective-independent), called “traditional realism.” At the other end of the continuum, population thinking assumes that FEAR and other emotion categories are constructed dynamically in a fully contextual and situated manner. One situated category for FEAR can vary substantially from another. Emotion categories, as situated constructions, are not genetically inherited adaptations, although their ingredients evolved via both natural selection and epigenetic influences. Lived experience, such as being socialized within a given culture, is assumed to play a key role in the transmission and evolution of emotion categories. The features of any occurrence of FEAR, such as physiological changes, facial movements, and so on, derive their emotional meaning relationally within the larger ensemble. No stable and immutable prototype exists for FEAR or any other emotion category. Population thinking uses holism as a scientific strategy. Instances of FEAR are expected to vary substantially from one another in their facial expressions, their physiological patterning, and so on, and emerge from complex causation. Scientific studies are designed to observe and model variation that is structured within spatiotemporal contexts (and the physical condition of the body is an important source of context). Context is assumed fully causal, on par with any genetic influences that are inherited via natural selection. Population thinking assumes that every category of FEAR and the other categories of reality are perspective-dependent, called “relational realism” (Barrett & Theriault, 2025).
Population thinking is not unique to the TCE. According to Mayr, population thinking was pioneered by Charles Darwin (1859). Darwin defined a species as a grouping of individuals with variable physical features whose fitness is inherently relative to the conditions of the immediate environment. 4 William James (1890) adapted Darwin’s insights in his writings on emotion over a century ago:
The trouble with the emotions in psychology is that they are regarded too much as absolutely individual things. So long as they are set down as so many eternal and sacred psychic entities, like the old immutable [pre-Darwinian] species in natural history, so long all that can be done with them is reverently to catalogue their separate characters, points, and effects. (p. 449, emphasis in the original)
James continued: “But if we regard them . . . as ‘species’ are now regarded as products of heredity and variation, the mere distinguishing and cataloguing becomes of subsidiary importance” (p. 449). 5 Since then, population thinking has been periodically revisited in psychological science under various names (e.g., Estes, 1956; Gallistel, 2012).
A partial list of the evidence for population thinking in emotion—that variation within each folk emotion category is substantial and structured by context—is provided in Table 2. Studies designed to sample and model structured variation routinely find supporting evidence in motifs of facial movements (e.g., Barrett et al., 2019; Durán and Fernández-Dols, 2021; Gendron et al., 2018; Gendron, Hoemann et al., 2020; Jack et al., 2016; Le Mau et al., 2021; Srinivasan and Martinez, 2021), autonomic nervous system physiology (e.g., Hoemann, Khan et al., 2020; Khalaf et al., 2020; McVeigh et al., 2023; Kragel and LaBar, 2013; Siegel et al., 2018; Stemmler et al., 2007; Stephens et al., 2010), and brain signal changes (e.g., Azari et al., 2020; Lebois et al., 2020; Singh et al., 2021; Wang et al., 2024; Wilson-Mendenhall et al., 2011, 2015). This variation consistently goes well beyond what could be described as a basic emotion family of signal patterns that share common configural properties.
Even the supposed prototypic brain “biomarkers” for a given emotion category vary substantially across contexts and studies (e.g., Horikawa et al., 2020; Kassam et al., 2013; Kragel and LaBar, 2015; Saarimäki et al., 2016; Wager et al., 2015; Wang et al., 2024; Wilson-Mendenhall et al., 2015; Zhou et al., 2021) and suggest the presence of meaningful, structured variation within an emotion category that goes well beyond anything that could be meaningfully described as a prototype or group of signal patterns with a family resemblance. A mathematical simulation demonstrates that even a single brain biomarker is a statistical fiction: a summary that need not be found in any participant’s data even if successfully classified (Clark-Polner et al., 2017).
Evidence for substantial cultural variation in emotion perception is now considerably stronger and more robust than evidence for universal similarities (Gendron et al., 2018), including in hunter-gatherers who live a foraging lifestyle that is similar to the niche in which presumed basic emotion categories were thought to evolve (Gendron, Hoemann, et al., 2020). Even the function of an emotion category can vary with the situation, inconsistent with the functionalist hypothesis that all instances of the same emotion category share a common, dedicated function (Adolphs, 2017; Levenson, 2011; Sznycer et al., 2017; Tooby & Cosmides, 2008; van Heijst et al., 2025, pp. 378, 382, and 386). For example, ANGER can serve different functions in different situations, including “remove an obstacle,” “punish,” “compete,” “identify with a group,” or “avoid harm,” and so on (e.g., Ceulemans et al., 2012; Sinaceur & Tiedens, 2006; Van Kleef & Côté, 2007; van Zomeren et al., 2004). ANGER can even signal investment or care (Pillsbury, 1989; for other examples of functional variation in the same folk emotion category, see de la Harpe et al., 2024; Lindquist et al., 2022; Mesquita, 2022; Uchida & Kitayama, 2009; Wong & Tsai, 2007).
The substantial, situated variation in emotion, as predicted by the TCE, is too large in magnitude to be consistent with the hypothesis that emotion categories are families of immutable prototypes issuing from an inherited genotype, triggered by a unique assembly of neurons, with inherent emotional meaning across situations, people, and cultures, as proposed in BET. As a consequence, empirical support for population thinking disconfirms the existence of biological and psychological emotion types as hypothesized in BET. 6 The reverse is not necessarily the case. When occurrences of the same emotion category share common features, the commonality is not necessarily caused by genes or dedicated neural circuits, as discussed elsewhere (Barrett, 2012, 2022; Barrett et al., 2019; Hoemann, Crittenden, et al., 2019).
Ad hoc categories in the theory of constructed emotion
In the TCE, population thinking extends to categories and not just their occurrences. A category is a group of instances that are similar enough to serve a function and guide allostasis and action in a specific context. A category is relational, situated, and ad hoc—constructed in the brain as a momentary, contextualized dynamic event (Barsalou, 1991; Barrett et al., 2015; Casasanto & Lupyan, 2015; see also Hoemann, 2023; Hoffman et al., 2018; Mesquita et al., 2010; Schyns et al., 1998; Spivey, 2008; Wilson-Mendenhall et al., 2011; Yee & Thompson-Schill, 2016). In other words, a category is something you do, not something you have. In BET, in contrast, a category is a static element of reality—a collection of instances that share a static prototype. These two views of categories are fundamentally incompatible, an observation that van Heijst et al. did not address when they discussed emotion concepts and categories in our theory.
Ad hoc categories
Each dynamic, ad hoc category event begins as a sample of possible, situated instances and ends as a single context-dependent instance. The category is hypothesized to begin as a sample of prediction signals—a collection of temporally evolving patterns of intrinsic signals assembled across the entire brain that share a similar control plan for allostasis and action in the next moment (something like an action tendency). This is similar to the idea of “policies” in reinforcement learning or action concepts (Barrett & Finlay, 2018). These signals might also share possible mental features of lived experience that arise from the motor plans, such as “goal,” “value,” “threat,” “reward,” “valence,” “arousal,” “novelty,” and so on. They could even share concrete sensory or motor features. The brain continually checks its prediction signals against signals that it receives from the body’s sensory surfaces, which derive from the state of the body and world. These incoming sensory signals help to select the pattern of prediction signals and in the process create the instance of the category that will manifest in allostasis, action, and lived experience. Once selection occurs, the incoming sensory signals and the allostatic and skeletomotor changes are said to be categorized and explained. A category event that begins as a group of brain-wide signal ensembles ends as a situated instance, such as a specific spatiotemporal instance of ANGER that can vary in its physical and mental features from other instances of ANGER for that person and for other people (i.e., in its neural basis, physiological changes in the body, facial movements, etc.). van Heijst et al. did not discuss this hypothesis or its relevance to the main hypothesis of their article.
Across a person’s lifetime, their brain constructs a population of n possible categories that can contain an emotion word as a feature, such as “anger.” This is a distribution of ad hoc categories (e.g., a distribution of ad hoc ANGER categories). A person’s distribution of ad hoc ANGER categories is full of variation because each category is created in the moment to serve a particular function or goal in a particular situation. This variation is not random; it is structured by context, and some of the necessary context is in a person’s brain (see Table 1). This structured variation, known as “emotional granularity,” “emotional complexity,” or “emotional expertise,” is necessary for the functional utility of emotion in everyday life (Barrett, 2017a; Hoemann et al., 2021; Springstein et al., 2024). One individual’s distribution of ad hoc ANGER categories may have a prototype, but that prototype is a simplification of situated variation that does not necessarily exist in reality. This is population thinking.
The vocabulary of ad hoc categories (emotion or otherwise) is hypothesized to vary across people and cultures in structured, predictable ways. Distributions of features will be more similar across people to the extent that their development, life history, and physical bodies are similar. For example, people who come from different backgrounds, with different opportunities to culturally inherit different knowledge about emotion, may have different distributions. As a consequence, English words such as “angry,” “sad,” and “fearful” name folk psychology categories that are populations of diverse, contextualized events such that the neural circuitry involved, the physiological changes, the expressive behaviors, the mental features, and so on, vary across situations and individuals (see Table 2). The summary of this overall distribution is what van Heijst et al. and others have hypothesized to be a prototype that sufficiently describes and predicts all instances of the folk category across contexts, people, cultures, and even species.
Relational meaning
The TCE—in direct contrast to basic emotion theories—hypothesizes that signals such as facial movements, vocal acoustics, changes in heart rate, changes in respiration, action potentials, chemical concentrations, and so on have no inherent, biologically prepared emotional meaning (for additional discussion and evidence, see Barrett, 2022). Instead, an instance of emotion is hypothesized to emerge from ensembles of relations among these richly differentiated and variable signals. In short, emotional meaning is relational (Barrett & Theriault, 2025). Instances of emotions emerge as incoming sensory signals are made meaningful when constrained and categorized by signals in the brain (e.g., Doyle & Lindquist, 2018), as evidenced by different brain-wide distributed patterns of blood-oxygen-level-dependent signal for instances of the same category (i.e., degeneracy; e.g., Doyle et al., 2022; Lebois et al., 2020; Wang et al., 2022; Wilson-Mendenhall et al., 2011, 2015). Different combinations of sensory and motor signals in different contexts can have the same emotional meaning (e.g., Goel et al., 2024; Hoemann, Khan, et al., 2020). You can scowl in anger, cry in anger, and laugh in anger (e.g., Le Mau et al., 2021). Likewise, the same signal can have different meanings (e.g., Goel et al., 2024; see also Barrett et al., 2019). And although a signal cannot have infinite meanings, it can have more than one. You can smile in happiness at a party, smile in anger during a family meeting, and smile in sadness at a funeral (e.g., Girard et al., 2021; Rychlowska et al., 2017). The variety of muscle movements that produce a scowl may variously accompany an instance of ANGER, CONCENTRATION, or a bout of INTESTINAL GAS.
With its focus on relational meaning, the TCE jettisons the deep commitments of BET to stable, universal, perspective-independent categories as collections of contextless objects, processes, and states. This focus echoes relational approaches to quantum mechanics in modern physics (see Rovelli, 2021). “Relational” here does not mean “subjective” (Ashton, 2020; Bloor, 2011). It does not mean that emotional reality exists only in a person’s head. Nor does it mean “arbitrary.” “Relative” means only that instances of emotion are conditioned on some perspective. “Relative” is the opposite of absolutism, such as the idea of universal, basic emotion categories. A perspective-independent emotion does not exist in any meaningful sense, and perspective-independent emotion is exactly what BET hypothesizes. This is another reason why the two theories of emotion are fundamentally incompatible.
Error 3: Failing to Understand the Extended Evolutionary Synthesis
There are two research traditions in evolutionary biology, one uniquely consistent with BET and the other uniquely consistent with the TCE. BET relies on the older modern synthesis (Hamilton, 1963; Huxley, 1942; Williams, 1966), which is a gene-centered view of evolution that integrated Darwin’s theory of natural selection with Mendelian genetics. The TCE relies on the newer extended evolutionary synthesis, which questions the basic, foundational assumption that phenotypic characteristics are genetically preprogrammed adaptations (e.g., Bateson & Laland, 2013; Jablonka & Lamb, 2005; Kirschner et al., 2006; Laland, 2017; Lewontin, 2000; Odling-Smee et al., 2003; Richerson & Boyd, 2004; for an accessible but detailed summary, see Ball, 2024). Genes certainly play a role in the extended evolutionary synthesis, but not as an immutable blueprint for an organism’s phenotype. They are considered to be inert ingredients that cells use, in interaction with the environment, to create phenotypes. Considerably more causal impact is accorded to influences that emerge during embryonic development and the lifetime of the individual, hence the nickname “evo-devo” for the extended evolutionary synthesis.
van Heijst et al. (2025) referred to the modern synthesis as “the cornerstone of evolutionary biology” (p. 378), which is factually incorrect (for an accessible yet comprehensive review, see Ball, 2024). They also oversimplified the extended evolutionary synthesis, characterizing it as a mere extension of the older modern synthesis, but this claim does not sit well with most scientists who developed the evo-devo approach (e.g., for accessible explanations, see Lewontin, 2000; Richerson & Boyd, 2004) or who utilize it, including us. We began explicitly incorporating many of these influences into our constructionist approach almost 20 years ago. One must understand the differences between the two evolutionary frameworks to appreciate how the TCE’s evolutionary foundations differ vastly from (and are incompatible with) those of BET.
Basic emotion theory and the modern synthesis
The modern synthesis is a reductionist approach in which genes, not organisms, are the target of natural selection. Organisms are assumed to be mere vehicles for genes, which is where all the evolutionary action is assumed to occur. The basic assumption—that genes are the primary units of inheritance and that phenotypic characteristics are genetic adaptations maintained by natural selection—is generally preferred in evolutionary psychology and is consistent with the assumptions of basic emotion theory.
“Basic emotions” are said to be genetically inherited phenotypic characteristics that are specific adaptations with fixed physical forms and fixed functions across time. For example, instances of FEAR are thought to manifest with a small degree of variation around a universal pattern of physiological changes, expressive behaviors, and so on (see van Heijst et al., 2025, p. 377). This pattern is hypothesized to have evolved as an adaptation to navigate the recurrent fitness challenge of encountering predatory threat. Its cause is reduced to a single, innate (hardwired) neural circuit that does not change or function differently across time, context, or individual. Sure, the assumed universal pattern of core features can be tweaked by contextual factors, but any variation in how instances of, for example, fear manifest is assumed to be overlaid on the basic form of a fear response that was designed by natural selection. As a result, it should be possible to use the presumed universal pattern for FEAR to identify instances of fear across time, context, and individuals (and maybe even species depending on whose version of BET you read). FEAR’s expression or triggers can be modified or tweaked after the fact by lived experience, culture, or other phenomena such as regulation strategies. These latter modifications, being separate from the genetic adaptations, would not influence the inheritance of FEAR or any other presumed basic emotion from generation to generation. Causally, each phenotypic characteristic is reduced to some combination of genes that have been selected to serve some function that increases evolutionary fitness. Gene transfer across generations (inheritance) is primarily, if not solely, a function of natural selection.
Whereas BET aims at discovering the presumed stability and sameness of a category’s function and the emotional meaning of physical properties across time and space, the TCE aims at explaining structured variation and complexity (see Error 2). Our theory is more consistent with the assumption that there is no genetically prespecified physical or functional forms of FEAR or of any other emotion category, and therefore emotions cannot be meaningfully viewed or understood through the lens of the modern synthesis. In the TCE, the function of neural assemblies, physiological changes, facial and bodily movements, and other components of any so-called fear pattern vary with the local context (including the momentary biological condition of the individual), as well as the individual’s development and life history (see discussion of ad hoc categories above). Structured variation and its dependence on local (contextual) conditions are assumed to be the norm. As a consequence, the TCE, with its focus on contextual variation and complexity, is more consistent with the extended evolutionary synthesis as a theory of evolution.
The theory of constructed emotion and the extended evolutionary synthesis
The extended evolutionary synthesis questions the existence of fixed adaptations designed by natural selection (for a discussion, see Bateson & Laland, 2013). Instead, it focuses on flexible, contextual responses. As Bateson and Laland (2013) stated: “Animals often have capacities to respond flexibly to local conditions with adaptive responses, many of which are themselves knowledge-gaining processes . . . apparent design can be derived in many different ways . . . and need not imply adaptation, or even selection” (p. 713). Gene expression is heavily regulated moment by moment. It is not a binary on/off, protein/no-protein situation but rather a continuous, context-dependent expression that is regulated ad hoc by numerous factors that are dynamic and malleable to environmental impact (see Ball, 2024). Put differently, contextual influences are assumed to be fully causal rather than merely modulatory. Causation is assumed to be complex, not simple and mechanistic.
To help explain this structured variation and complexity, the extended evolutionary synthesis has documented various evolutionary processes that contribute to phenotypic variation beyond the ways in which natural selection impacts gene frequencies across generations in the historical past. These processes include (but are not limited to) various epigenetic influences, including cultural inheritance, and niche construction. Together, they exert a powerful influence on phenotypic characteristics during the development and lifetime of the individual, dissolving the boundary between evolutionary and developmental influences (e.g., Bateson & Laland, 2013; Jablonka & Lamb, 2005; Laland, 2017; Lewontin, 2000; Odling-Smee et al., 2003). These evo-devo phenomena, each of which is backed up by substantial empirical evidence, as well as gene-culture coevolution, contribute to the transdisciplinary hypotheses that are integral to the TCE. Evo-devo considerations have only passing relevance for BET in its current form, however, in part because development in BET is considered to be an unfolding and shaping of innate adaptations.
Cultural inheritance of emotion categories
Here is one example of a hypothesis from the TCE that incorporates the extended evolutionary synthesis and cannot be comfortably situated within the modern synthesis. A caregiver’s allostatic support of an infant (e.g., feeding, soothing, changing diapers, shared attention, singing, speaking) is hypothesized to create an environment full of influences (a niche) that allows infants to learn the language and categories of their culture, including emotion categories (Atzil et al., 2018; Barrett, 2017a, 2020a). What we are describing here is the cultural transmission of category and language development, with an emphasis on the cultural inheritance (e.g., Boyd et al., 2011; Richerson & Boyd, 2004) of emotions rather than genetic inheritance alone (e.g., Atzil & Abramson, 2021; Barrett, 2017a; Gendron, Mesquita, et al., 2020; Lindquist et al., 2022). Humans depend on one another for survival early in development (in infancy) and in the longer term (throughout the life span). We impact each other’s metabolic efficiency, creating a necessary context for cultural inheritance via mechanisms of learning and neural plasticity. These dynamics help set the stage for the further evolution of category construction across generations (Barrett, 2017a, 2020a; Lindquist et al., 2022).
In the TCE, natural selection is hypothesized to keep certain genes in place that not only allow for but also require the influence of developmental, life-history, and ecological influences. Genes are necessary but not sufficient. They open the door for a human brain to be continually shaped by (and coupled to) signals from the body and the world, including signals from the actions and words of others with whom they interact. Those signals are mandatory inputs for a human brain to learn to construct categories. The phenomena that emerge from those categories, such as emotions, cannot exist without them. During development and the processes that scientists call “socialization,” the words (Barrett, 2017a; Gelman & Roberts, 2017; Hoemann, Xu, et al., 2019; Shablack & Lindquist, 2019) and actions of others (Atzil & Gendron, 2017; Atzil et al., 2018; Barrett, 2020a; Gendron, Mesquita, et al., 2020; Mesquita, 2022) create recurrent situations that allow a brain to learn specific, situated meanings of autonomic nervous system changes, movements, vocalizations, and so on in the natural and cultural ecology of a person’s environment (see also Lupyan & Lewis, 2019; Ruba et al., 2022; Tenenbaum et al., 2011).
To claim that a neural mechanism for FEAR or any emotion is homologous across species, solely as the result of particular genes, requires that those genes are under an unusually strong selection pressure not to change (i.e., that individuals containing mutations or distributional changes in those genes literally die before reproducing). This possibility is extremely unlikely. 7 A more empirically defensible hypothesis is that phenomena emerge because natural selection requires signals from the body and environment to instruct and shape brain wiring (i.e., humans have the kind of nature that requires nurture). The biological system must learn from proximal life experience; this covers phenomena such as a visual system that develops the ability to see, specifically, out of eyes that are a certain physical distance apart in someone’s skull, as well as the phenomena that are transferred across generations by cultural inheritance. Natural selection keeps the necessity of cultural influences in place, and as a result, some ensembles of signals (i.e., information) are transmitted across generations not by genes alone but by mechanisms that require experience to develop their proper function.
Our hypotheses regarding the cultural inheritance of emotion categories are an example of the “cultural-drive hypothesis” (Laland, 2017): Human ancestors learned to control fire to cook food, which reduced the size of the gut, freeing up more metabolic resources. They also figured out how to cooperate, copy each other, communicate efficiently, and so on. Together, these innovations allowed our ancestors to live longer and grow bigger, more complex brains that could construct categories of greater abstraction (including emotion categories) and transmit those categories across generations via the practices of socialization and culture (Barrett, 2020a; see also Heyes, 2018, 2020; Tomasello, 2019). This information transfer across generations is hypothesized to influence who survives long enough to bear children, how many children they have, and the survivability of those children, in effect influencing gene frequencies across generations. In this way, culture and biology mutually influenced one another (e.g., culture-gene coevolution). The evolution of the capacity to construct ad hoc emotion categories is hypothesized to be integral to, not separate from, this epigenetic landscape.
According to the TCE, emotion words are culturally inherited tools that help to organize the world in particular, culturally relevant ways. They are tools of niche construction. Words are useful and important (although strictly speaking not necessary) as powerful invitations to learn categories and develop concepts about them (Giffin et al., 2017; Waxman & Gelman, 2010; Waxman & Markow, 1995), even for very young infants (Vouloumanos & Waxman, 2014). A word’s function, therefore, goes beyond mere labeling. van Heijst et al. (2025) misrepresented our hypothesis about the role of words in the TCE by focusing solely on the labeling of experiences (p. 380; in addition, labeling is more than surface-level classification because its functions can be both social and biological; see Chater & Christiansen, 2010; Hoemann, 2024).
All animals use categories to reduce the complexity of signals they process to navigate and shape their niche (i.e., the part of the world that is relevant to them). This is meaning-making in action. Every moment of life can be described this way. For vertebrates, meaning-making includes signals that continuously arrive from the sensory surfaces of the body, the dynamic intrinsic signaling in the brain, and low-level motor signals that control allostasis and skeletomotor movements (e.g., signals that control the contraction of certain smooth and skeletal muscle fibers, the angle of certain joints, the local pH for certain collections of cells). The many, many signals form a high-dimensional pattern (in which each signal is a dimension) that is massively complex. For n independent signals, there are 2n outcomes. The complexity must be reduced in meaningful ways for a brain to function. Categories reduce complexity by creating momentary equivalences. Research suggests that a word prompts a search among instances for some underlying sameness (features of equivalence) that transcends any noticeable differences (e.g., Booth & Waxman, 2002; Gopnik & Sobel, 2000; Graham et al., 2004; Nazzi & Gopnik, 2001; Welder & Graham, 2001). Even categories that are assumed to be concrete, such as “bird” or “cat,” can understood as populations of situated, abstract categories that are created in the moment to serve a particular function or goal (e.g., the category “bird” when you are in a pet store is different from “bird” when you are hungry; see Barsalou, 1983). The TCE hypothesizes that words are abstract features that support the learning of emotion categories, which are abstract categories.
Psychological scientists have defined the word “abstract” in several ways, but in this discussion, to “abstract” means to generalize from different patterns of concrete sensory and motor features to the same mental feature, for example, to generalize from sensory patterns and motor movements to “threat” or “value” or “pleasure.” Different patterns of multiple sensory and motor features share the same abstract feature. Many detailed sensory and motor features (patterns of high dimensionality) are summarized into fewer abstract features (of lower dimensionality). 8 The simple act of reading a word, even an emotion word such as “fear” or “joy,” or hearing a word involves patterns of activity that are widely distributed throughout the brain, extending into sensory and motor areas (e.g., Lee & Satpute, 2024; Pulvermüller et al., 2005; Wilson-Mendenhall et al., 2013b, 2019), consistent with this view.
We have provided evidence to justify the utility of the hypothesis that words support emotion category learning (Barrett et al., 2007; Hoemann, Xu, et al., 2019; Hoemann, Wu, et al., 2020; Lindquist, 2017; Lindquist et al., 2015; Satpute & Lindquist, 2021; Satpute et al., 2020; for a discussion, see also Barrett, 2017a), and a growing body of developmental evidence supports it (Nencheva et al., 2024; Nook et al., 2020; Ogren & Sandhofer, 2021, 2022; Plate et al., 2023; Price et al., 2022; Ruba et al., 2021), as well as the usefulness of words for category learning in adults (Chadbourne et al., 2022; Doyle & Lindquist, 2018; Fugate et al., 2010; Gendron et al., 2012; Lindquist, Gendron, et al., 2013; but see Hoemann et al., 2022). 9 We have also discussed the manner in which an emotion word itself can serve as the abstract feature of equivalence for an ad hoc emotion category (Barrett & Lida, 2024; Barrett & Theriault, 2025).
A word helps a person learn that different signal patterns—movements, physiological changes, incoming sensory signals, and so on—are functionally equivalent in a given context because those instances, despite their sensory and motor differences, serve the same goal in a given situation. Relational features, such as “to remove an obstacle” or “unpleasant,” are abstract features that make the different patterns of sensory-motor signals functionally equivalent in a specific situation. Abstract features create similarities out of differences—varied patterns of concrete sensory and motor features are made equivalent (similar) by their shared function in a specific context rather than by their physical similarities. This arrangement allows a person to flexibly enact a function by generalizing across instances that may differ in their concrete spatiotemporal contexts. The specific skeletomotor movements that enact a functional goal (e.g., to remove an obstacle) will depend on the specifics of the situation, the anatomy and the energetic state of the body, and the person’s lived experience. Accordingly, a person might scowl and shout in anger, cry in anger, freeze in anger, or even laugh—whatever action concept the brain has learned to construct to meet the goal in similar situations. The movements of the heart, lungs, and other tissues in the body (i.e., visceromotor movements) that support the movements of the body (i.e., skeletomotor movements) can also vary with the situated goal. And the neural signals that manifest the category, which are distributed across the brain and give rise to these varying features, will themselves vary from situation to situation, instance to instance, a phenomenon we defined earlier as “degeneracy” and is also known as “multiple realizability” (Edelman & Gally, 2001; Marder & Taylor, 2011; for evidence and discussions on how degeneracy pertains to emotion, see Barrett, 2017b, 2022; Doyle et al., 2022; Wang et al., 2024; Westlin et al., 2023). Relational hypotheses predict that different ensembles of physical signals streaming in from the sensory surfaces of the body can have the same relational emotional meaning across situations and persons. In addition, a single physical signal or pattern of signals can have different relational meanings in different situations and persons, in the same way that any sensory input can be conceptually categorized in a flexible, situated manner (as keenly demonstrated by various visual illusions). The power of words, as we conceive it, far outstrips the impact of labeling as van Heijst et al. described it.
Importance of brain structure and ecological niche
Evolutionary hypotheses are often evaluated by comparing observations across species. Research in the basic emotion tradition, guided by the modern synthesis, often compares species that are genetically close to humans, making other great apes, such as the common chimpanzee (Pan troglodytes), an optimal comparison. The evo-devo approach used by the TCE requires that such comparisons also consider the developmental, anatomical, niche, and life-history similarities among the species, however, in addition to their genetic similarity. This is because any phenotypic characteristic, such as an instance of ANGER or FEAR, causally emerges from a complex interaction of these factors and is not merely an adaptation that was shaped by the natural selection of genes.
Degrees of abstraction
Considering niche similarity across species is important for understanding and formulating hypotheses within the TCE in a manner that is largely overlooked by BET. van Heijst et al. mentioned comparisons to great apes, as our closest living genetic relatives, more than a dozen times. But the various species of great apes, including humans, have very different environmental niches that favor different mental capacities. Common chimpanzees, for example, have some capacity to form and learn abstract categories, but they have much better memory for sensory particulars and therefore have better episodic memory (Matsuzawa, 2010) compared with humans, which is critical for survival in their ecological niche: They acquire food by foraging over miles of territory, so memory for concrete details (e.g., this particular tree, with these particular colored leaves and those particular smells, at that particular time of day, is where this kind of fruit can be grabbed with these muscle movements) is much more functional than a higher level of abstraction (e.g., the category “tree”). By implication, we would hypothesize that humans are more likely to construct and use categories of higher abstraction, including more abstract emotion categories, when compared with chimpanzees, who may be more likely to construct and use more situation-specific, concrete categories. 10
BET hypothesizes that common chimpanzees make particular facial configurations to express emotions that are supposedly homologous to human expressions, such as “bared-teeth” faces (construed as fear), “lip-press” faces (construed as anger), and so on (see Kret et al., 2020; Parr et al., 2007). But BARED TEETH and LIP PRESS are human-made categories that, like all categories, abstract away the variable details of muscle movements to create similarities in function. Even humans with extensive experience working with chimps require the assistance of words to learn these abstract categories (Fugate et al., 2010). In all of this research, scientists working in the tradition of BET reduce the complexity of chimp behaviors using their own human categories that may or may not be relevant in the chimps’ niche. The emotion categories the scientists apply to chimps are not even relevant in all human niches (i.e., cultures). Scientists hypothesize, observe, and infer within the bounds of their landscape of categories, and the phenomena they study are relational, not objective and perceiver-independent. An evo-devo approach instructs us to carefully consider whether the categories we use in research are actually homologous with the categories that chimps themselves make and use in everyday life (i.e., the relational reality from the chimps’ perspective; Barrett, 2017a; Bliss-Moreau, 2017).
All animals categorize (Mareschal et al., 2010). Even single-cell organisms can be said to categorize as they generalize from the past to predict and act in the service of allostasis (Freddolino & Tavazoie, 2012). If the TCE is correct, then the world does not come presorted into categories that all animals share. All animal brains can be described as automatically and effortlessly forming ad hoc concepts to guide allostasis and action and thereby categorize incoming signals and create lived experience (whatever that means for the animals in question). We have hypothesized elsewhere that what differs among species, and what may have changed across evolution, is the degree of abstraction that a brain is capable of constructing because of general brain-scaling functions (Workman et al., 2013) and the signals available in an animal’s niche (e.g., Barrett, 2017a; Barrett & Finlay, 2018). For example, the human brain has expanded association cortices in the frontal lobes, parietal cortex, and inferotemporal cortex compared with other primates, including other great apes (e.g., Sherwood et al., 2012, 2017). There are also changes in the metabolic costs of brain function (e.g., Kuzawa et al., 2014), particularly in the upper layers of the cerebral cortex (e.g., Krienen et al., 2016; Sherwood & Gómez-Robles, 2017; Wei et al., 2019) that play a distinct role in the predictive processing aspects of categorization (Barrett, 2017b; cf. Bastos et al., 2020; Brincat et al., 2018; Theriault et al., 2023).
More generally, there are substantial differences in metabolic function between humans and our closest genetic relatives, common chimpanzees (e.g., Clarke & Sokoloff, 1999), and energy regulation, including allostasis, is crucial to understanding evolutionary change (e.g,. Lane, 2014; Pontzer, 2015; Sterling & Laughlin, 2015). These metabolic considerations are an important part of the transdisciplinary basis for the TCE (for discussions, see Barrett, 2017a, 2017b; Barrett & Finlay, 2018; Hutchinson & Barrett, 2019): Brain expansion and metabolic change potentially allow for the increased signal compression and dimensionality reduction afforded in the human brain, creating the opportunity for human emotion categories of greater abstraction. This hypothesis in no way diminishes the importance of survival-related behaviors in human emotion or invalidates the importance of studying survival-related behaviors in animal models for the purposes of understanding part of the biology of human emotion and its disorders. This hypothesis does suggest, however, that solving the puzzle of human emotion requires more than just mapping circuits in the brains of adult animals with little regard for development, anatomy, niche, and life history.
Importance of development
We close our discussion of evolution with a final example of why evo-devo considerations are different from those of the modern synthesis. Scientists working within the tradition of the modern synthesis often make evolutionary inferences by comparing the phenotypes of adult animals because development is thought to be a mere unfolding of preformed genetic programs and epigenetic influences are thought to merely tweak genetic adaptations. But developmental considerations do not merely tweak genetic adaptations—they are fully causal factors, as are niche, life history, and anatomical features of the whole organism. As a consequence, inferences about evolution are best made when looking at how development—from embryo onward—unfolds (for this evo-devo perspective on brain evolution, see, e.g., Finlay & Uchiyama, 2017; Workman et al., 2011, 2013). The genetic, intrinsic (e.g., reproductive mode) and extrinsic (e.g., environmental) sources of epigenetic influences that alter gene expression are referred to as epigenic inheritance—the transfer of influences across generations that are rooted in non-DNA sequences. Epigenic inheritance can vary, sometimes substantially, from moment to moment within an individual, not to mention across species such as rodents and humans, making simple evolutionary inferences untenable when based on genes alone (for a discussion, see Anastasiadi et al., 2021). Again, van Heijst et al.’s emphasis on the modern synthesis ignored these influences.
Both Theories Rest on Irreconcilable Assumptions
van Heijst et al. did not integrate BET and the TCE. They integrated BET with an oversimplification of the TCE. Our actual theory cannot be integrated with BET because the two are incommensurate. The most fundamental assumptions and hypotheses of the two theories are at odds (see Table 3 and Figure 1; for a discussion of incommensurability, see Feyerabend, 1962; Kuhn, 1970; Oberheim & Hoyningen-Huene, 2018). BET is a typological theory that relies on reductionism and mechanistic notions of cause and effect (called “efficient causation”): A specific genotype produces a neural circuit (or few circuits) that produce a type of emotion whose function is stable across instances. Context moderates the triggers and expressions of that type. As a consequence, there is a family of physical signal patterns for each emotion category that is best described as variation around a fixed prototype. Lived experience and culture are important to emotional development but not the evolved nature of emotion; emotions are assumed to be genetically preprogrammed adaptations. The TCE, in contrast, is a population theory that relies on holism and the complexity of causation (Barrett & Theriault, 2025). Many interacting influences, including context, produce a causal impact that is more than the sum of its parts. There is considerable variation in both the function and the form of occurrences within the same emotion category and no necessary family resemblance among them. The so-called “core characteristics” or prototype of a folk emotion category is assumed to be a stereotype (an oversimplified, widely held belief). Lived experience and culture are necessary both for the development and for the evolution of emotion categories.
Comparison of Assumptions and Hypotheses in the TCE and BET

Note: TCE = theory of constructed emotion; BET = basic emotion theory.
Trying to understand the TCE using the assumptions and hypotheses of BET is like trying to understand quantum mechanics using Newtonian mechanics. It works only if you oversimplify quantum mechanics by removing most of its complexity, its relational foundations, and its key hypotheses, producing something that ultimately is not quantum mechanics.
From the perspective of our theory, the emotion types of BET are simplified stereotypes that do not exist in reality. They appear to exist only when contextual relations are ignored or hidden and therefore taken for granted (Barrett & Theriault, 2025). Simplifications are necessary tools in science and in life, and some simplifications are more useful than others, but it is a mistake to believe they are real in a fundamental, perceiver-independent way. Replacing real variation and complexity with human-made simplifications, and then believing those simplifications are real and the variation and complexity are only apparent, is called “surreptitious substitution” (Husserl, 1970) in philosophy.
Furthermore, we understand typological thinking as a case in which an abstraction that occurs in the brains of humans is assumed to be real and immutable in nature, and variation is considered error. This is another example of surreptitious substitution, regardless of whether the abstraction is statistical (e.g., the central tendency of a phenotypic distribution) or conceptual (e.g., the inferred function of the phenotypic characteristic). The hope of the typological thinker is that such abstractions will eventually reveal clear evidence of psychological laws that govern human behavior (see Danziger, 1990, p. 129). The goal of population thinking is to discern meaningful structure in omnipresent variation.
A cautionary example: the so-called James-Lange theory of emotion
Theories, and the experiments they give rise to, are laden with assumptions (Dubova & Goldstone, 2023; Westlin et al., 2023). Failing to appreciate the compatibility of those assumptions, as van Heijst et al. have done, comes with a cost: It can introduce serious errors into the publication record that persist for decades. A case in point is the so-called James-Lange theory of emotion. William James and Carl Lange each described emotions as perceptions of bodily changes, but their assumptions were as different as the TCE and BET, respectively. So how did it come to pass that the two theories were integrated into the James-Lange theory of emotion, a popular view of emotion that has inspired scientists for more than a century? The answer is its author, the eminent philosopher John Dewey, bifurcated emotions into objective physical states versus feelings and ignored the parts of James’s population-thinking theory that did not fit with Lange’s typological thinking. Like van Heijst et al., Dewey himself relied on typological assumptions about emotion. And voila, the “James-Lange” theory of emotion was born. (Dewey proposed a discrete emotion theory in a separate article (Dewey, 1894). 11 )
James’s and Lange’s ideas about emotion are incommensurable. James’s writings on emotion (James, 1884, 1890, 1894/1994) and radical empiricism (James, 1897) are similar to the TCE’s population thinking (Barrett & Theriault, 2025). James was writing about individual occurrences of emotion and emphasizing their variation and contextuality. James observed that a person might tremble in fear in one context, jump in fear in another, attack in fear in yet another, and even laugh in the face of fear if the situation demanded. Each occurrence of fear, according to James, involves a different set of physical changes and sensations. In James’s words, “‘fear’ of getting wet is not the same fear as fear of a bear” (James, 1894/1994, p. 206), and “surely there is no definite affectation of ‘anger’ in an ‘entitative’ sense” (James, 1894/1994, p. 206). Lange’s vasomotor theory of emotion, by contrast, was aimed at describing the unique bodily state that accompanies all instances of a single emotion category (Lange, 1885/1912). Lange’s theory—which was published the same year as James’s “What Is an Emotion?”—used BET’s typological thinking. 12
Dewey’s misleading simplification of James, and its downstream consequences for the impact of James’s actual writings, is one of the great mistakes in modern psychology. It is also a cautionary tale to those who might be tempted to downplay the importance of philosophical assumptions and the hypotheses that they give rise to, particularly incommensurate ones. The James-Lange theory is taught as foundational knowledge in most introductory psychology textbooks and has shaped the thinking and practice of generations of scientists. Yet it is founded on 100-year-old errors that occurred when Dewey ignored James’s ideas about variation and contextuality that challenged assumptions about emotion types. Who knows what the last century of research on emotion would have accomplished by now if these errors had been corrected when they were introduced. Or if they had been avoided in the first place.
Conclusion
Van Heijst et al.’s article is part of a long typological tradition of scientific research on emotion. In a recent survey of experts in the science of emotion, 80% of the respondents endorsed typological beliefs (Ekman, 2016). Typologically inspired research, rooted in the modern evolutionary synthesis, contains certain simplifications that make it easy for scientists to misunderstand the TCE, which rejects typological thinking and focuses on variation, contextuality, and the importance of lived experience as a necessary causal force in creating emotions, on par with genes. Whenever such scientists assimilate our theory into their own tradition, they inadvertently reduce ketchup to sugar and create simplifications that are deeply misleading. When scientists are unaware of their own assumptions, they may even ignore published evidence that precisely calls those assumptions into question. If the history of psychology is any use in predicting its future, the science of emotion will remain mired in a scientific stalemate until this blind spot is resolved.
Any evolutionary analysis begins by specifying the phenomena to be explained (Hogan, 1994). In the science of emotion, this specification contains assumptions about the nature, origin, and function of emotion categories and their relation to other biological and psychological phenomena. The TCE relies on very different assumptions than BET. Emotions are not part of some perceiver-independent reality. Everything you do, including your scientific practices, is possible only given the background of categories that you inherited from the social and cultural practices that you embody. That a group of like-minded scientists experiences a freezing rat as fearful, a chimp with bulging lips as angry, or a crying human as sad does not reveal the objective reality of those emotion categories. It might, instead, reveal the shared set of assumptions that are, themselves, a set of concepts acquired through cultural inheritance.
A scientific research tradition, at its best, crafts a usable system of conceptual tools for thinking about what exists in the world, that is, reality—a system of concepts that works well enough for humans to act in the world, and even add to the world, in ways that interest them. Until it does not. And then it is time to entertain a different conceptual framework with an open mind. This is what the historian and philosopher Thomas Kuhn called a “scientific revolution” in his famous book, The Structure of Scientific Revolutions (Kuhn, 1970). Scientific revolutions are rare, it turns out, but some of Kuhn’s other insights have stood the test of time (so far). “Structure” is, fundamentally, a critique of typological thinking and its attendant assumptions in scientific practice. Scientific knowledge is not discovered—it is created. It is fundamentally relational. It depends on shared categories, goals, values, and agreements about which phenomena exist (i.e., which ontology to use), which questions are admissible, and which methods count as acceptable tools of inquiry (again, see Barrett & Theriault, 2025).
Scientific knowledge exists as collective organized experience (Bogdanov, 2020, p. 113) that emerges as a consensus among the brains of diverse individuals (Longino, 1990). Every way of scientifically knowing the world depends on the actions of a group of humans in agreement. To produce valid scientific knowledge, however, that consensus must be achieved authentically, by engaging with others’ hypotheses as they intended. The scientific community must be self-critical (Longino, 1990), engaging directly and robustly with evidence that is potentially disconfirming. Scientific work must be considered in its historical context, and scientific advances must be given proper consideration, even when they call deeply held beliefs into doubt. Only then can consensus among diverse scientists produce trustworthy, justified knowledge that is useful in the real world.
Footnotes
Acknowledgements
We thank Chan Hong, Kieran McVeigh, and Serife Leman Runyun for their helpful comments on earlier drafts of this manuscript.
Transparency
Action Editor: Kent Berridge
Editor: Interim Editorial Panel
Author Contributions
All authors after L. F. Barrett are listed in alphabetical order. All of the authors approved the final manuscript for submission.