
Abstract
Although chemical signaling is an essential mode of communication in most vertebrates, it has long been viewed as having negligible effects in humans. However, a growing body of evidence shows that the sense of smell affects human behavior in social contexts ranging from affiliation and parenting to disease avoidance and social threat. This article aims to (a) introduce research on human chemical communication in the historical context of the behavioral sciences; (b) provide a balanced overview of recent advances that describe individual differences in the emission of semiochemicals and the neural mechanisms underpinning their perception, that together demonstrate communicative function; and (c) propose directions for future research toward unraveling the molecular principles involved and understanding the variability in the generation, transmission, and reception of chemical signals in increasingly ecologically valid conditions. Achieving these goals will enable us to address some important societal challenges but are within reach only with the aid of genuinely interdisciplinary approaches.
Mammals use smell in almost all domains of their social lives. For instance, newborn rabbits search for nipples by following a mammary pheromone (Schaal et al., 2003), mice avoid the smell of sick conspecifics (Kavaliers et al., 2020), and ring-tailed lemurs engage in social displays known as stink fighting (Evans & Goy, 1968). However, if we were to replace these individual species names with humans, many readers would raise their eyebrows with suspicion or skepticism: Most people do not think of humans as a species that communicates by smell (Doty, 2010). But such a suggestion should not come as a surprise because the human body is richly endowed with apocrine and sebaceous glands that produce hundreds of chemical compounds in individually variable amounts, bestowing each individual with a unique chemical signature (Natsch & Emter, 2020). Many of these compounds are perceived by the human sense of smell, which, contrary to previous belief, is well developed. For example, it has high discriminating power (Gerkin & Castro, 2015) and is capable of detecting odorants at threshold concentrations that are (at least for some substances) comparable with, or even lower than, those of more famously smell-oriented species, such as dogs and mice (Laska, 2017). Furthermore, the high convergence ratio of human olfactory sensory neurons into olfactory bulb glomeruli indicates a sophisticated fine-tuning of olfactory signal processing at the periphery of the system (Maresh et al., 2008), even before cognitive elaboration of odor percepts in higher level brain networks.
What role, then, does the sense of smell and other chemical senses play in the social lives of humans? In many respects, this remains terra incognita, a point stressed previously by including the possible existence of “human pheromones” among a list of outstanding scientific questions to be addressed during this century (Kennedy & Norman, 2005).
However, this situation is rapidly changing, with significant advances in our understanding over recent years. The aim of this article is therefore not only to introduce the state of the art of human chemical communication across variable social contexts but also to demonstrate how this research contributes to general psychological concepts and theories. We highlight challenges of current research and outstanding questions, providing a roadmap for future research, some aspects of which have the capacity for reaping societal benefits. To fulfill these aims, the article is structured along a broad chronological perspective. The first section briefly introduces why olfaction was long considered of little use to interacting humans, before reviewing findings from the second half of the 20th century within the theoretical frameworks of that era, including the quest for human pheromones. The second section reviews the current understanding of chemical communication, focusing on its role in mate choice and perception of emotional and health states while also considering interindividual differences in perception and situating airborne information among inputs from the other senses. Finally, a third section outlines future research avenues, highlighting rapid technological developments that offer new opportunities for progress. We conclude by considering possible benefits of a broader understanding of chemical communication and its implications for our everyday social lives.
Chemical communication in animals has been defined in evolutionary terms. According to this perspective (Leonhardt et al., 2016; Maynard-Smith & Harper, 1995; Wyatt, 2014), which can equally be applied to humans, chemical communication involves at least one individual emitting chemicals (the emitter) that can be received by at least one other individual (the receiver; Fig. 1). Notably, if only a receiver has evolved by natural (or another form of) selection the capacity to receive and extract information from an emitter’s chemical compounds, the medium is called a cue. However, if the emitter also has adaptive benefit from sending the information to the receiver, the medium is considered a signal (whereby the “emitter” is now labeled “sender”). It should be noted that even if we refer to “the emitter/sender” and “the receiver” in singular form, chemical communication potentially involves every individual nearby, and so more than one emitter/sender and receiver can be expected to be present. However, interpersonal distance moderates the likelihood of chemical communication, especially when the medium contains chemical compounds of low volatility, which are perceived predominantly at close range. This implies that some compounds may primarily serve a function in close, dyadic communication, whereas others may reach a broader range of receivers.
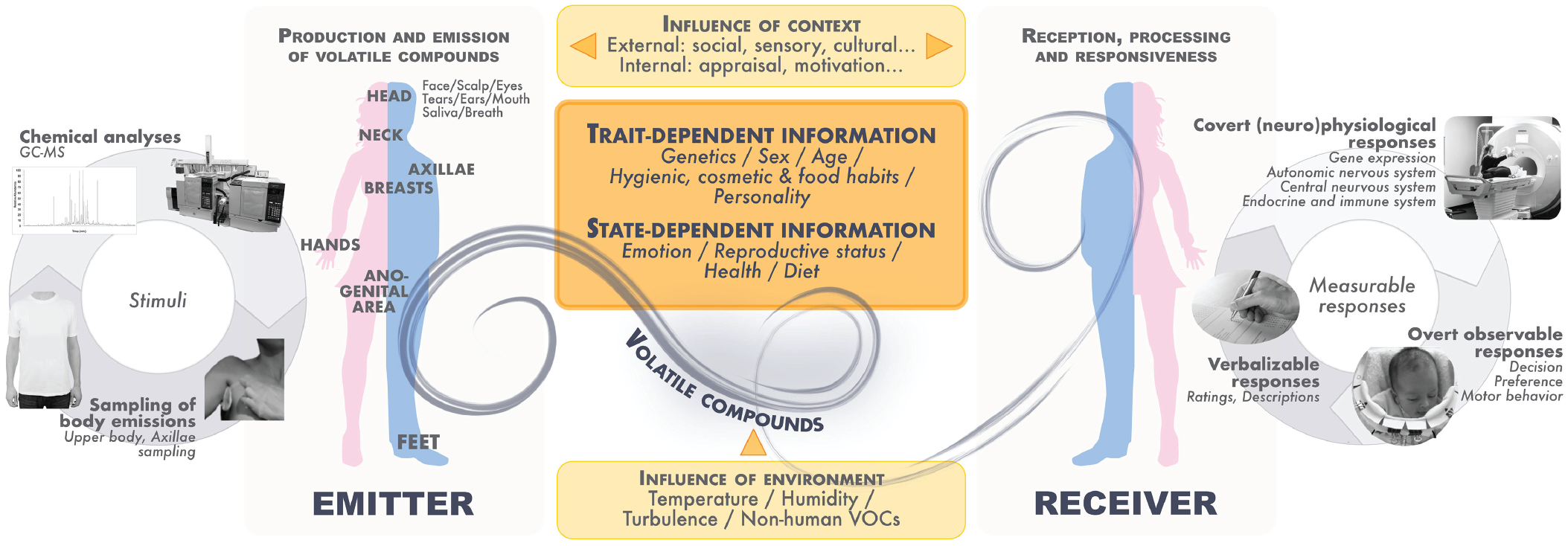
Overview of our current understanding of human airborne chemical communication. Airborne volatile compounds emitted by one or more individual(s) (emitter) reach one or more receiver(s) and are detected and processed, respectively, by chemosensory systems and emotional and social brain networks. Internal and external context influences the processing of trait- and state-dependent information at various levels. It often remains unknown whether these factors predominantly influence the emitter, receiver, or both. Chemical communication ultimately leads to adaptive behavior in the receiver. Contemporary methods used to study chemical communication in humans are illustrated, highlighting sampling and analysis of volatiles to generate/characterize the stimulus (left) and assessment of stimulus effects on verbal, overt, and covert behavior (right). Measurable effects in the receiver relate to different concepts such as attention, orientation, interest, approach/withdrawal, social cognition, social contagion, and decision-making.
Olfactory cues and signals may provide adaptive benefits to receivers by modulating psychological processes (behavior, affect, cognition, and/or physiology) in ways that can be witnessed in various domains and life stages, from avoiding physical danger or diseased conspecifics to promoting social bonds in mother-child interactions, friendship, and romantic relations. Although evolutionary theories often provide us with ultimate (or functional) explanations for certain forms of chemical communication, psychological theories are especially useful in forming testable predictions about proximate causes—a more mechanistic perspective that leaves room for idiosyncrasies and the role of context in shaping behavior. As detailed in our chronological overview, decades passed before psychologists treated chemical communication as more than a scientific curiosity, arguably because of the erroneous assumption that studying invisible molecules, unintentionally produced by emitters and the effects of which escape receivers’ conscious awareness, is unimportant. However, “unintentional and unnoticed” does not equate to ineffective and nonfunctional (think of pathogens, for example), and we showcase here how chemical communication research itself adapted, over time, through the increasing adoption of psychological perspectives.
The Past
Microsmatic humans?
Throughout history, the communicative function of body odor has generally been appraised with reservation compared with the relatively exalted role of olfaction in gastronomy, perfumery, and medical diagnosis (Reinarz, 2014). The Enlightenment gave rise to modern science but did little to illuminate the social nature and value of human body odor. Intellectual figures (with notable exceptions; Le Guérer, 1992) disparaged olfaction as a source of rational knowledge, aiming ultimately to differentiate humans from reeking and snuffling animals. Both producing and detecting smells by sniffing became unwelcome in public settings and discourse (Corbin, 1988), expunging olfaction as a means of communication.
This process of sensory demotion contaminated the nascent domains of psychology, biology, and anthropology. For example, in The Descent of Man, Charles Darwin (1871) opined that to humans “the sense of smell is of extremely slight service, if any.” Olfaction’s academic nadir arrived shortly thereafter, with the grouping of Homo sapiens among animals bearing an involuted olfactory tract, such as birds, cetaceans, and other primates: Humankind was therefore termed “anosmatic” (Broca, 1879), although this was later toned down to “microsmatic” (Turner, 1890). The notion of human microsmaty drifted from Broca’s anatomical definition to a generalized use in functional terms that found fertile ground in the prudish, courteous elites of Western societies. Prejudice against olfaction in general, and social olfaction in particular, pervaded academia, including psychology. For instance, in his psychosexual theory of human development, Freud (1905) considered the sense of smell as being associated with animosity. Freud also conjectured that the evolution of an upright posture led to the “organic repression” and psychological devaluation of the human sense of smell (Freud, 1930/1961). Consequently, research on human olfaction received little attention until the 1950s and 1960s (for notable exceptions, see Henning, 1924; Passy, 1895; von Skramlik, 1926; Zwaardemaker, 1895). It is worth noting that, after the identification of the genes coding for olfactory receptor (OR) proteins by Buck and Axel (1991), the idea of human microsmaty was revived by molecular biologists on the basis of the low endowment of functional OR genes relative to other mammals (Gilad et al., 2003). Later evidence, however, contested the link between olfactory receptor pseudogenization and effective underreliance on olfaction in everyday behavior (Shepherd, 2006), and the measurement of endocranial casts revealed larger olfactory bulbs in modern H. sapiens relative to H. neanderthalensis (Bastir et al., 2011), indicating that human olfactory capability has not decreased at least since our latest common ancestor. Meanwhile, researchers have increasingly objected to the macro/microsmaty division in animals and have progressively invigorated the field of human chemical communication by pulling together research lines from ethology and psychology, neurosciences and physiology, ecology and evolution, chemistry and biochemistry, and more recently cultural history and anthropology. By the end of the 20th century, a step change was in process, with new research centers and teams dedicated to the chemical senses and publishing modern textbooks devoted to the chemosensation with increasing emphasis on related communication in humans (e.g., Müller-Schwarze & Mozell, 1977; see also Brever et al., 2006; Doty, 2015; Engen, 1982; Getchell et al., 1991; Laing et al., 1991; Rouby et al., 2002; Serby & Chobor, 1992; Takagi, 1989; Van Toller & Dodd, 1992; Zucco et al., 2012).
The quest for human pheromones
A first research wave in favor of human chemical communication was conveyed by sensory physiology through Le Magnen’s (1952) work exploring the fluctuations in women’s olfactory sensitivity. He found that the perception of odorants bearing potential reproductive significance is affected by menstrual cycling and gestation, introducing the idea that the communicative impact of body odorants depends on sensory abilities that are aligned with reproductive status and designed to optimize reproductive success. From the 1950s onward, with the rise of ethology and comparative psychobiology, social olfaction was investigated in a wide range of vertebrates, especially mammals, evidencing the fundamental influence of chemosignals in the regulation of reproductive and stress physiology and the organization of social behavior. The concept of “pheromone,” a term coined by Karlson and Lüscher (1959) in their work on insects, was subsequently extended to mammals to account for species-specific signals evolved for communication. The term “pheromones” was originally defined as “substances which are secreted to the outside by an individual and received by a second individual of the same species, in which they release a specific reaction, for example a definite behavior or a developmental process” (Karlson & Lüscher, 1959, p. 55). The chemical composition of insect pheromones was elucidated by the late 1950s and found to be monocomponent (e.g., bombykol; see Butenandt et al., 1959) or multicomponent (e.g., the sex pheromone of Rhyacionia frustrana; see Berisford and Brady, 1973). Depending on the pheromone’s mode of action, it was qualified as a “releaser” (obvious behavioral effect), “primer” (endocrine or developmental effects), or “informer” (attentional or cognitive effects) pheromone (Müller-Schwarze, 1977).
The term was soon applied to other taxa, including mammals, and speculation about the existence of human pheromones quickly followed (Comfort, 1971), with three notable emerging themes. First, findings that odorous steroids (e.g., androstenone, androstenol) excreted in boar saliva or urine trigger the typical precopulatory stance of the sow led to scrutiny of these and similar compounds in humans. They were detected in axillary sweat and saliva, with higher concentrations in men (Gower & Ruparelia, 1993), and their emotional, cognitive, and social effects were widely tested (for a review, see Havlíček et al., 2010). Second, a mixture of five aliphatic acids (so-called copulins) emitted by female rhesus macaques were also found in human vaginal secretions (Michael et al., 1975) and tested for effects on male attraction or physiology, with no positive results (Morris & Udry, 1978). Finally, odor-based physiological responses have been documented extensively in rodents since the 1950s in relation to stress or reproduction (leading to the so-called Lee-Boot, Whitten, Bruce, and Vandenbergh effects). Thus, estrus synchronization evidenced in rodents (McClintock, 1978) stimulated investigation for homologous phenomena and underlying odor-based mechanisms in cohabiting women (Preti et al., 1986; Stern & McClintock, 1998). In the first publication on human menstrual synchrony, McClintock (1971) reported that female students living together tend to synchronize their menstrual cycle (Fig. 2a). McClintock speculated that this might be due to pheromones. Much later, Stern and McClintock (1998) proposed the underlying mechanism, arguing that women’s exposure to axillary compounds produced during the follicular phase shortens their cycle, whereas exposure to compounds produced around ovulation lengthens it (Fig. 2b).
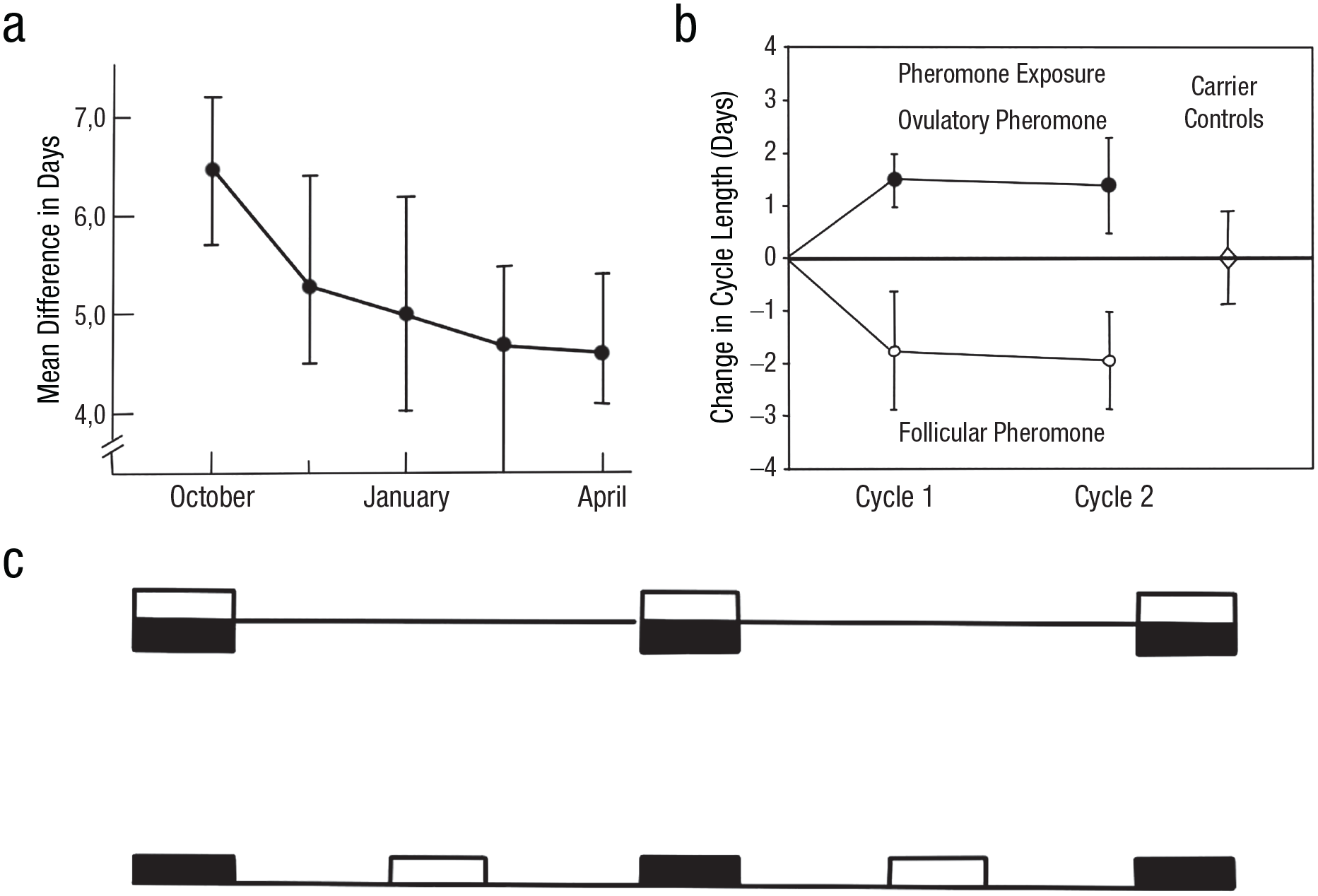
Menstrual synchrony. Top left panel (a) has been adapted from McClintock (1971) and shows the increase in menstrual-onset synchrony in groups of cohabiting female friends over 7 months. On the top right, the increase in cycle length after exposure to body odor from unfamiliar women collected during ovulation and decrease in cycle length after exposure to the odor collected from women during the follicular phase is depicted (panel (b), adapted from Stern and McClintock (1998)). At the bottom, in panel (c), extreme possibilities in menstrual-phase synchrony between two women (white and black box) are shown. The top part shows complete synchronization (the menstrual phases of the two women completely overlap). The bottom part shows maximal desynchronization (the menstrual phases of the two women are constantly half a cycle length apart). Panel (c) is adapted from Strassmann (1997).
Menstrual synchrony is frequently reported as the best-documented candidate of a human pheromonal effect, but there are several criticisms related to the concept of pheromone-induced menstrual synchrony. First, dozens of studies have tried to replicate the first effect (i.e., existence of synchrony), and results are highly inconsistent. Perhaps most noteworthy are two studies from nonindustrial societies, the Dogon of Mali (Strassmann, 1997) and Bedouin of Israel (Weller & Weller, 1997), neither of which found menstrual synchrony. Second, there has been much criticism of how synchrony was computed, and it has been shown that cycles of different length can synchronize and desynchronize by chance (Schank, 2006). Third, most studies compute overlaps of menstrual bleeding, but, as pointed out by Strassmann (1997), the mean difference in any two 28-day cycles is 7 days, and menstrual bleeding lasts on average 5 days (Fig. 2c). Therefore, menstrual bleeding in many female dyads will overlap purely by chance, giving an impression of synchrony. Finally, there is no solid functional explanation for menstrual synchrony (Barash & Lipton, 2009). Taken together, there are still numerous questions regarding the existence of the phenomenon, its possible mechanism, and its function.
Although the three illustrative cases of androstenes, copulins, and menstrual synchrony stimulated creative experimental emulations that have continued unabated, the quest for a human pheromone still remains unfulfilled. No compound(s) in the human volatilome has yet been isolated and ascertained to unconditionally and reliably release stereotyped behaviors or to prime physiological responses (Doty, 2010; Wyatt, 2015). We do not know whether this is because human pheromones do not exist or because we have not yet adopted the correct methodology to find them if they do (Wyatt, 2015). Whichever is correct, the story of the search for human pheromones illustrates that uncritically translating to humans olfaction-based phenomena as they occur in sometimes phylogenetically distant species was not the optimal strategy to guide human chemical communication research. Such extrapolations across species occasionally led research into dead ends, as in the case of vomerolfaction. Chemoreception by the vomeronasal organ plays a central role in the reception of social chemosignals in many vertebrates, and, for a time, the vomeronasal organ was also believed to be a sensory player in human chemical communication (Berliner et al., 1996). However, it now appears to be nonfunctional in adults (Trotier et al., 2000; Witt & Hummel, 2006), and, in any case, studies in other mammals (Dorries et al., 1997; Hudson & Distel, 1986) have found its occurrence unnecessary for the detection of well-characterized specialized chemosignals.
Discoveries in the era of sociobiology
From the 1960s through the 1990s, parallel upsurges in the social sciences and sociobiology rekindled attention on human social odors beyond the sole enthrallment with pheromones. Whereas social psychologists conceive of odorants as another medium of nonverbal communication and a means of impression management (Baron, 1988), behavioral ecologists and evolutionary psychologists view them as having both proximate (physiological/psychological) and ultimate (evolutionary/adaptive) functions. In this light, human chemical communication was studied in the laboratory using explicit stimulation and declarative methods derived from olfactory psychophysics (e.g., rating scales of stimulus properties, such as intensity, hedonic valence, familiarity, or sex typicality). Methods of odorant presentation for implicit perception were also implemented to measure ensuing nonverbal responses, which are not necessarily associated with the allocation of attentional resources (e.g., general arousal, orientation behavior, autonomic nervous system responses).
This research evidenced olfaction-based differentiation of social categories (self, individual, kin, or group identity; sex; age; personality) or processes modeled after the social behavior of other species. Theoretical concepts such as sexual selection, sexual receptivity, filial and parental bonding, and incest avoidance were the dominating topics in animal behavior research of that era. The causes and consequences of social recognition, and preferences between emitting and receiving individuals, thus became central research themes in human research as well. On the basis solely of their body odors, adult partners or child peers were found to be capable of recognizing each other (Hold & Schleidt, 1977; Mallet & Schaal, 1998), whereas mothers can identify their infants (Schaal et al., 1980), and neonate infants orient to their mother’s breast (Russell, 1976; Varendi et al., 1994). Human-specific odorants were conceived as externalized indices of genetic fitness or physiological readiness that guide mate choices or parental responses and, in principle, can drive selective allocation of social investment and optimization of immediate survival, social inclusion, and inclusive fitness. For example, it was found that humans may find the odor of major histocompatibility complex (MHC)-dissimilar individuals more pleasant than MHC-similar individuals (Wedekind et al., 1995), as previously reported in other species; mating with an MHC-dissimilar partner would result in heterozygous offspring with enhanced immunocompetence.
Furthermore, the choice of extrinsic odorants (fragrances) appears complementary, rather than antagonistic, to such processes (Havlíček & Roberts, 2013), and social psychologists have evidenced communication processes involving these odorants in social desirability effects, such as first impression management, display of social distinction, and inclination to interact, help, or aggress (Baron, 1988). More specifically, it has been shown that people tend to select fragrances complementing their own unique body odor (Lenochová et al., 2012; Fig. 3). Furthermore, natural body odor is to some extent dependent on one’s genetic makeup, and populations thus vary in body odor composition. As different fragrances may suit different body odor phenotypes, individuals in different populations may prefer different fragrances, which in turn may shape cultural values. However, such gene-fragrance associations may also work the other way around: Fragrances valued by a particular culture (for reasons other than genetic makeup, such as religion or marketing-constrained mimicry) may not suit some phenotypes and, if such values endure across numerous generations, may ultimately affect population genetic structure. In other words, culture may affect biological evolution and vice versa (e.g., Henrich, 2020).
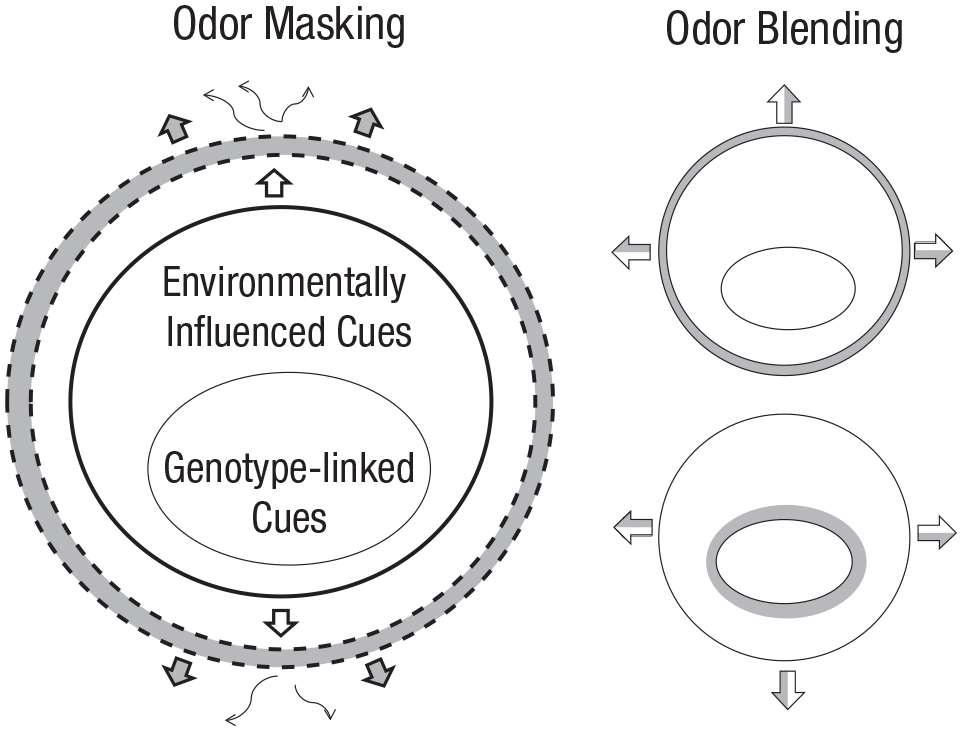
Proposed mechanisms for the interaction between fragrance and body volatiles. In odor masking (left), an applied fragrance forms a perceptual sheath (gray shading) over the naturally occurring volatiles emanating from the body (white arrows). Although some naturally occurring volatiles (thin arrows) may still “leak” through, the dominant smell perceived by others is that of the fragrance (gray arrows). In contrast, in odor blending (right), the odor perceived by others is a blend of the naturally occurring volatiles (white) and those contained in the fragrance (gray). Two possible blending mechanisms are depicted that reflect how a suitable perfume might be chosen for a given individual. Above, the fragrance is chosen to create a blend with the entire chemical profile; below, it is chosen to form a blend specifically with the genetically linked compounds (note these are extreme characterizations of blending mechanisms; in practice it is likely that only a proportion of genetically and environmentally influenced compounds are involved). Adapted from Allen et al. (2019).
In sum, 20th-century research on human chemical communication debunked some tenacious ideas that had dismissed its very existence. First, biological and anthropological findings discredited the prevalent, although vague, notion of human microsmaty (McGann, 2017; Schaal & Porter, 1991; Stoddart, 1990). Second, we gained confirmation that the emission of volatile mixtures from multiple sites of the human body are chemically differentiable along perceptually valid social dimensions such as age, sex, individuality, and health (Doty, 1981). Third, although the most impactful compounds in these chemical signatures still need to be identified, they were shown to be clearly perceivable and learnable and to bear multiple meanings for receivers. Finally, we learned that the generalized human behavior of superimposing extrinsic odorants (fragrances) on their natural odor should not be considered antagonistic to a communicative function (Havlíček & Roberts, 2013) but rather synergistic in a holistic human chemical communication system integrating both biology and culture.
The Present
Work since the turn of this century has provided a still deeper understanding of the range of information available to receivers and the principles of human airborne chemical communication (Fig. 1), facilitated by the emergence of psychological theories (to study proximate causes) complementing evolutionary frameworks (providing information about ultimate causes). In this section, we highlight several key areas of current inquiry.
Chemosensory communication in cooperation and mate selection
Humans recognize not only the body odors of themselves and their kin but also those of friends (Mallet & Schaal, 1998; Olsson et al., 2006). It was recently reported that “click” friends (persons instantaneously feeling a strong bond toward each other) smelled more alike than expected by chance, and similarity in body odor was correlated with better social interaction between strangers in a mirror game (Ravreby et al., 2022). Although it is difficult to prove a causal relationship beyond this observation, it is in line with work demonstrating associations between friendship and similarity in other domains (Selfhout et al., 2009).
Indeed, recognition of friends may be functionally related to kin recognition because friends’ genotypes also tend to be correlated, and there is an unusual degree of homophily in olfactory receptor genes among friends (Christakis & Fowler, 2014). Recognition of kinship might have contributed to fostering kin-directed altruistic impulses and enhanced inclusive fitness (Christakis & Fowler, 2014), but the role of chemical cues in cooperation need not be restricted to kin. Culture-dependent odors may—like visual cues such as hairstyle or dress—also operate as in-group markers and promote trust and reciprocal cooperation. This function could either be served deliberately (e.g., by choice of a given fragrance) or unintentionally (e.g., by body odor compounds related to shared diets).
Perceiving the body odors of kin may help to modulate responsiveness in social interactions. For instance, in childcare, the identification of one’s own child leads to a preference for this odor compared with other children’s odor, with potential consequences for a positive interaction (Schäfer et al., 2020). On the other side of the interaction, 4-month-old babies look longer at their mother’s face compared with an unfamiliar female face when exposed to maternal volatiles (Durand et al., 2020). These bidirectional effects of body odors on social interactions may be one of the sensory foundations of infant-to-parent attachment and parent-to-infant bonding. This is well supported by a brain-to-brain synchrony study using dual-EEG recording during a mother-infant versus stranger-infant face-to-face interaction episode (Endevelt-Shapira et al., 2021). Although interbrain synchrony was expectedly higher in mother-infant dyads relative to stranger-infant dyads, this difference vanished when own-mother’s odor was brought into the scene of the stranger-infant interaction. Thus, mother’s body odor, even in her physical absence, enhances interbrain synchronization with an unfamiliar adult in correlation with an infant’s increased visual attention. Such maternal odor-mediated social engagement appears specific to own-mother’s scent: Although 7-month-olds exposed to a fearful face display an EEG fear response of similar magnitude in control and stranger-mother’s odor conditions, the typical fear response disappears in the context of their own mother’s odor (Jessen, 2020). Own-mother’s body odor thus appears to be an essential facet of an infant’s secure base. In a different context but pertaining to a similar issue, in adults, smelling a partner’s body odor in a stressful separation situation diminishes experienced discomfort and autonomic reactivity (Granqvist et al., 2019).
Chemosensory cues play a role in sexual selection (e.g., initiation and maintenance of romantic relationships; for a review, see Mahmut & Croy, 2019). For instance, men perceive female body odors as most attractive during the fertile phase of the menstrual cycle (Habel et al., 2021; Havlíček et al., 2006), and preference for spousal odor is positively correlated with relationship duration (Sorokowska et al., 2018). Regarding preferences for MHC, it has been suggested that human mate choice is more affected by MHC diversity than by MHC dissimilarity (Winternitz et al., 2016). However, most studies on MHC-related body odor preferences have been conducted in populations with high genetic diversity, although for most of their evolutionary history humans lived in more homogeneous, small-scale societies. It is therefore possible that people tend to avoid individuals with high MHC similarity but show no systematic preference beyond a certain similarity threshold (Croy et al., 2020; Havlíček et al., 2020; Pause et al., 2006).
Chemical cues to emotional state and health status
Receivers exposed to fear-related volatiles assume a fearful facial expression and show concomitant increases in eye-scanning behavior and air intake, which would improve detection and reaction to potential threat (de Groot et al., 2012). These findings are intelligible within a functionalist theory on emotions (Susskind et al., 2008), from which emotional expressions have been argued to primarily have a self-serving (rather than a communicative) function by preparing an individual for adaptive perception and action (already noted by Darwin, 1872/1998). Specifically, a fearful facial expression has a physical sensory gating function: By opening the eyes and nose, a larger amount of visual and chemical information can be acquired to better deal with threat (de Groot et al., 2012; Susskind et al., 2008). Emotional states evoked by chemical cues may be unique among the senses by escaping conscious and linguistic access and by fully relying on automatic emotion processes. As receivers take on the fearful facial expression mimicking the state of the sender, the chemical communication of fear (as an unconditioned response to threat) or anxiety (as a conditioned response to threat-related internal and external stimuli; LeDoux & Pine, 2016) resembles a physiologically ancient form of empathy that results in the contagion of emotions (de Groot et al., 2012; Prehn-Kristensen et al., 2009; for emotional-contagion theory, see Hatfield et al., 1993). In addition, human aggression-related body odorants (sampled from sweat donors in response either to competitive sport contests or to frustration) can provide cues to social status and dominance, as shown by their effects on physiological, behavioral, and neural responses in receivers (Mutic et al., 2017; Pause et al., 2020).
Volatile compounds in sweat can also provide cues of respiratory disease (Sarolidou et al., 2020), as well as viral, bacterial, protozoan, and multicellular parasitic infections (Havlíček et al., 2017). Within hours of experimental activation of the immune system, body odors alter and become less pleasant (Olsson et al., 2014; Regenbogen et al., 2017). Unpleasant odors automatically activate behavioral withdrawal systems; for example, they increase the strength of the startle response. In addition, the activation of behavioral withdrawal systems by odors cannot be as easily reduced by intentional emotion regulation efforts compared with stimuli of other modalities (Adolph & Pause, 2012). Chemical cues can thus provide receivers with a potent long-distance cue about infection status, and, accordingly, olfactory detection and response may be an integral part of the behavioral immune system—a set of behaviors, motivated by disgust, that are thought to have evolved to minimize the risk of contagion (Schaller & Park, 2011). Disgusting odors trigger avoidance in receivers (Croy, Angelo, & Olausson, 2014) via a neural network involving the olfactory- and tactile-processing areas (Croy et al., 2016) as well as adaptive preparatory immune responses (Juran et al., 2023).
Evolutionary theories make a convincing case for the chemical communication of disease and emotions by focusing on their evolutionary origins and the selective pressures shaping a receptive neuroanatomy that would increase receivers’ survival chances. By contrast, psychological theories are especially insightful in providing proximate, mechanistic explanations for behavior caused by human volatile compounds and allow us to predict and test the conditions under which the strongest effects might occur. From a psychological point of view, odor cues to a person’s emotion and health status can be particularly potent in priming and motivating behavior outside conscious awareness (see also Smeets & Dijksterhuis, 2014). Theoretical models such as the situated inference model (Loersch & Payne, 2011) help explain why and when odor primes are effective, namely when people misattribute the chemically primed information to, for example, the (nonolfactory) perception of a particular object (construal priming). Smells are excellent primes because misattribution occurs only when people are unaware of the (effect of the) prime (Loersch & Payne, 2011), a condition that is easily achieved with olfactory cues. The processing of olfactory stimuli frequently bypasses brain regions involved in conscious attention and language processing (e.g., thalamus, inferior frontal gyrus) and directly probes areas related to emotions and associative memory (e.g., amygdala, hippocampus), making olfaction unique among the senses (Olofsson & Gottfried, 2015). Olfactory priming effects are especially pronounced when the odor matches the preparedness of the person. For instance, individuals with high levels of social anxiety show more pronounced withdrawal-related behavior (startle reflex) when exposed to anxiety-related sweat samples than individuals with low levels of social anxiety (Pause et al., 2009).
Variation in perception
The perception of, and responsiveness to, semiochemicals can vary greatly between individuals. Differential effects occur in men and women (de Groot et al., 2014a; Pause et al., 2010, 2020), with women usually outperforming men in processing fear- and anxiety-related volatiles and in responsiveness to them. This may reflect an adaptive sex difference, and such differences can also manifest in other intriguing ways. For example, differential processing of chemosensory anxiety versus emotionally neutral cues (especially increased processing speed of anxiety-related odor) is almost absent in pregnant women compared with nonpregnant women (Lübke et al., 2017), which may diminish the contagion of anxiety and hence reduce potential stress-related neurodevelopmental disorders in the fetus.
Individual differences are also related to personality and mental state. As mentioned above, individuals with high levels of social anxiety (Pause et al., 2009, 2010) and those with a history of childhood maltreatment show an altered responsiveness to anxiety-related sweat samples that can be dampened by intranasal oxytocin application (Maier et al., 2020). Depressed mothers do not exhibit typical preference for their own child’s body odor, and they are less able to recognize their baby’s body odor, even though the hedonic perception of strangers’ babies is unaffected (Croy et al., 2019). Likewise, in individuals with autism, the subliminal presentation of stress-derived volatiles does not lead to the expected reduction of trust toward a robot doll (Endevelt-Shapira et al., 2018). Nevertheless, autistic children profit from maternal olfactory cues in a social synchronization task (Parma et al., 2013).
Multisensory integration
Researchers have begun to investigate how chemosensory inputs are integrated with entries from the other sensory domains. For example, visual attention of infants toward pictures of social (vs. nonsocial) objects is increased in the presence of maternal volatiles (Durand et al., 2013). Furthermore, when exposed to maternal volatiles, 4-month-olds look longer at their mother’s face compared with an unfamiliar female face (Durand et al., 2020) and show enhanced face-related brain responses (Rekow et al., 2020). In adults, emotional cues contained in stress-related sweat (obtained from individuals experiencing stress, fear, or anxiety) facilitate the detection, processing, and categorization of ambiguous or faint visual danger signals, such as fearful or angry faces (Kamiloğlu et al., 2018; Zhou & Chen, 2009a). Weak or ambiguous happy facial expressions, on the other hand, make less of a social safety signal when they appear within a context of stress-related sweat (Pause et al., 2004). Stress-related chemical cues also aid in the detection and decoding of faint visual social signals in general (Rocha et al., 2018), suggesting that they are capable of enhancing vigilance toward otherwise inconspicuous social information, possibly by modulating the perception of incongruent or inconspicuous information (de Groot et al., 2014b). Even when visual scenery provides information that a given conspecific is not in danger, the suffusion of stress-related chemical cues promotes an empathic brain response (Hoenen et al., 2018). Thus, in decoding the social environment, the brain uses information from multiple sensory channels. When visual input is ambiguous or emotionally neutral, information from the chemosensory modality overrules the visual modality. When information from different modalities is congruent, chemosensory cues promote signal sharpening.
Neural underpinnings
Previous research considered social volatile compounds to predominantly activate diencephalic brain structures such as the thalamus and hypothalamus, regulating basic biological functions and hormone release (Savic et al., 2005). In contrast, more recent studies have suggested that social chemosensory information is also processed in networks of the emotional (limbic) social brain (e.g., amygdala, fusiform gyrus, insula, ventromedial prefrontal cortex; de Groot et al., 2021; for reviews, see Lundström & Olsson, 2010; Pause, 2017). In fact, it has been proposed that chemical communication aided in the evolutionary formation of the human social brain (Bastir et al., 2011). The hypothesis that the development of social perceptual and behavioral skills is paralleled by an increase in neocortex size is consistent with Dunbar’s social brain hypothesis (Dunbar & Shultz, 2007). Furthermore, the biological relevance of social signal processing is reflected in the observation that social sensory information is processed in non-modality-specific neural networks of the social brain (Alcalá-López et al., 2018).
Many human body odorants occurring at concentrations well above the perceptual threshold are processed in olfactory brain areas (Schäfer et al., 2019). However, independently of their odorous intensity, emotion-related chemical cues are predominantly processed in social brain areas (Bò et al., 2020; Pause, 2012). For example, chemical anxiety cues activate areas involved in the processing of social emotional stimuli (fusiform gyrus) and regulation of empathic feelings (insula, precuneus, cingulate cortex; Prehn-Kristensen et al., 2009). Human axillary sweat sampled from aggressive individuals activates limbic structures (Mutic et al., 2017), and in women, the dorsomedial prefrontal cortex (Brodmann area 8), priming threat-related physiological responses (Pause et al., 2020). Human disgust-related chemical cues activate the fusiform face area involved in the processing of visual social cues, and the orbitofrontal cortex, being related to value-based decision-making (Zheng et al., 2018). Furthermore, smelling friends activates brain regions known to be involved in processing familiar faces and voices (posterior parts of the retrosplenial cortex; Lundström et al., 2009). Activation of the caudate nucleus, an integral part of the brain’s reward system, is described in primiparous women smelling newborns (Lundström et al., 2013) and in socially open individuals smelling regular human body odorants (Lübke et al., 2014).
In sum, recent research on human chemical communication has developed into a serious research program yielding further insights into its functional value and underlying mechanisms. First, social volatiles affect communication in various domains of human social life, including cooperation, mate choice, parenting, and emotional state. Second, the perception of social volatiles is subject to high inter- and intraindividual variation based on proximate, psychological factors such as personality and actual mental state, in addition to evolutionary mechanisms ultimately affecting all human beings. Third, social chemical cues show complex integration with other sensory modalities, modulating or even overruling inputs from the other modalities. Finally, social volatiles are processed in social and emotional brain areas rather than purely olfactory brain areas, carefully sculpted over the course of evolution but still malleable to present-day psychological factors.
The Future
In this section (see also Fig. 4), we outline and illustrate main avenues in a research roadmap that will further extend our understanding of human chemical communication. This includes greater emphasis on its chemical basis, its impact in more ecological settings that include other-modality communication and the presence of other sources of volatiles and odorants, multidisciplinary collaborations to cover current research gaps, and opportunities for the application of these discoveries.
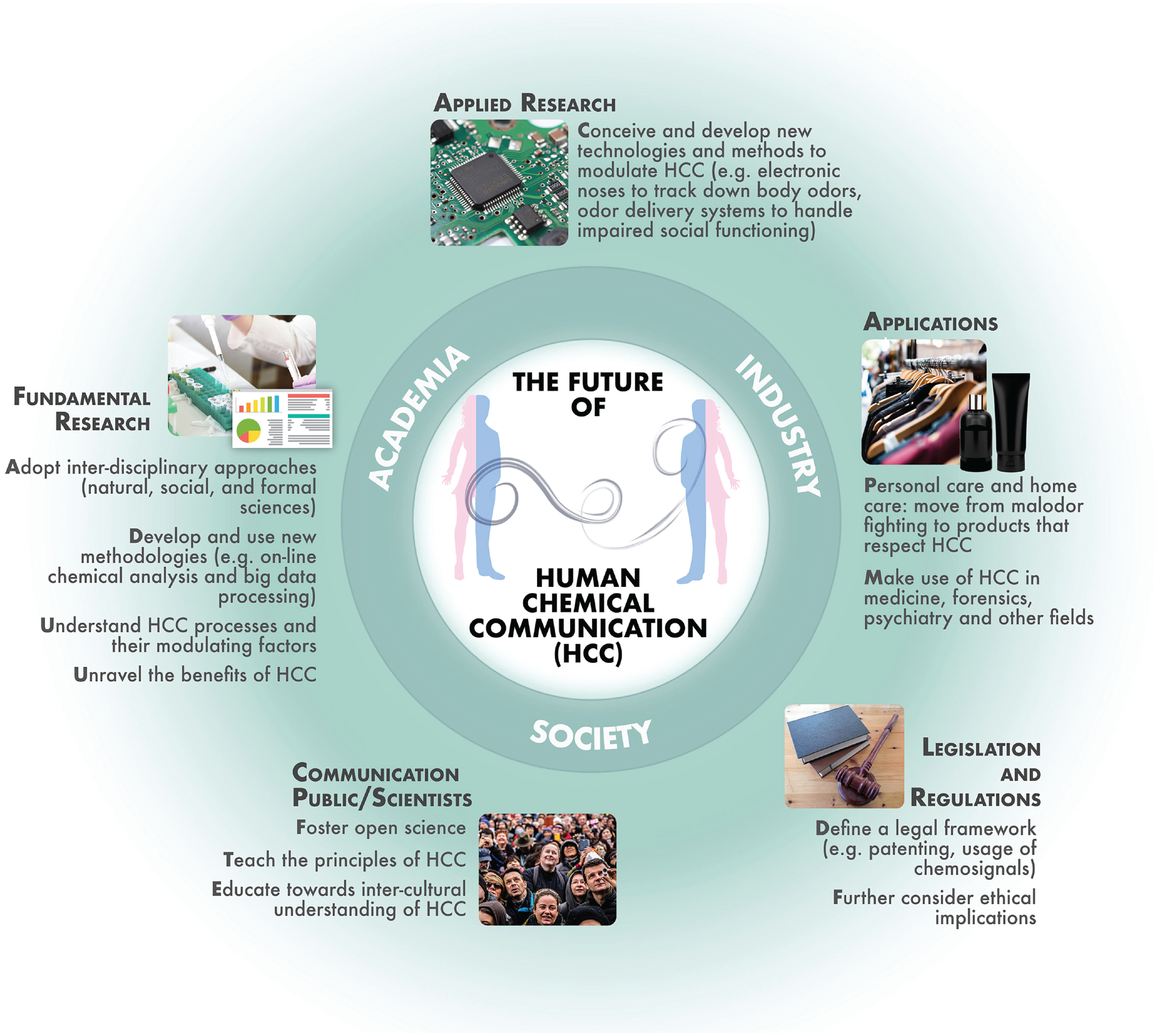
Current ideas of the future of human chemical communication research and its applications considering the interplay between academia, industry, and society. Further insights into the functional significance of human chemical communication and its underlying mechanisms by interdisciplinary fundamental and applied research and communication of these insights to industry and society are prerequisites for recognizing further benefits of chemical communication and conceiving related applications. Future developments may entail the need for further consideration of ethical aspects and development of adequate legal frameworks with respect to the usage of human chemical-communication processes.
Factors modulating chemical communication
Chemical communication research in humans initially focused on unidirectional communication from emitter to receiver. Now that we have seen convincing evidence that supports this form of communication, an important future focus would entail the investigation of adaptive benefits from receiving reciprocal information from the receiver. An example of this is that in the case of fear, the fear-related volatiles could alert others to impending danger, thereby recruiting their attention and help as a benefit to their own survival or rather to warn others so they can find a safe haven, thus providing a benefit to others. This would imply a shift of focus from assessing whether or not the receiver indeed perceived the emitter’s cues (e.g., by measuring facial expressions or autonomic arousal) to measuring ensuing behaviors and establishing any potential benefits. This will help us to distinguish whether the communication involves simply cue perception by receivers or can be classed as true chemosignaling (see Roberts et al., 2022).
Communicative signals and benefits will not be fixed, delivering predetermined outcomes, but will vary depending on the wider social and environmental context in which the communication occurs. Thus, research needs to diversify, going beyond older, deodorized, and desensitized (ODD) people (Roberts, Havlíček, & Schaal, 2020) and, more generally, Western, educated, industrial, rich, and democratic (WEIRD; Henrich et al., 2010) societies by including individuals from different age groups, societies, and ethnicities. Some variations in body odors exist across ethnicities (Prokop-Prigge et al., 2016), across cultural norms (e.g., rules of interpersonal distances; Sorokowska, Sorokowski, et al., 2017), and because of environmental factors (e.g., temperature, humidity, air pollution) or daily practices such as hygiene or the use of fragrance and cosmetic products. Such variations in what may constitute signals need to be considered to understand how chemical communication was shaped by biological and cultural coevolution (Havlíček & Roberts, 2013). Genetic determinants of odorant production (Martin et al., 2010) and olfactory perception (Keller et al., 2007), which may vary in geographical distribution (Wysocki et al., 1991), may also account for variations in chemical communication.
In addition, as noted above, human olfactory perception shows high inter- and intraindividual variability (e.g., related to age, hormonal state, medication) and substantial plasticity in response to contextual factors (de Araujo et al., 2005; Lundström & Olsson, 2005). This suggests that receiver benefits are modulated by social context (who is involved, where, and when), cognitive and cultural factors (expectations, stereotypes), or individual state (health, hormones, emotion). This makes it clear that beneficial behaviors may be far from the systematic and stereotypical (yet still variable) responses induced by some invertebrate pheromones (Doty, 2010). Although we have started to gain insight into such variability (see The Present section), future research is needed to fully understand the complexity of human chemical communication from both the emitter’s and the receiver’s sides and in more ecologically valid settings, including where there is more than one emitter or receiver.
Although the above relates to enhanced external validity, there is also a need for an enhanced focus on internal validity, reliability, and objectivity of the design and target measurements to avoid contributing to what is often called the reproducibility crisis (Nelson et al., 2021). This is particularly important in chemical communication because (a) body odors as stimuli are variable and weak in general; (b) rating scales or behavioral measurements, which are typical targets, are subject to bias and context effects; (c) olfactory presentation and processing need considerably more experimental time than visual or auditory stimuli (Menzel et al., 2019), which places limits on stimulus number and hence statistical power; and (d) participant sample sizes are typically small for similar reasons. Increased power through collaboration and increased standardization through the training of researchers, preregistration, and registered reports, as well as a careful interpretation of the findings (e.g., reflection of the correlational or causal nature of a given relationship), are important means to ensure scientific quality (Baldwin, 2017).
Refining and broadening our knowledge of semiochemicals
Although research has demonstrated the impact of chemical communication on human behavior, the detailed nature of the underlying chemical compounds remains a mystery. Chemical analyses, using a variety of procedures for sampling and analyzing volatiles emitted by the human body and focusing on different substrates such as breath, sweat, skin, milk, or urine (Drabińska et al., 2021; Loos et al., 2019; Penn et al., 2007), have shown different chemical signatures to be associated with gender (Penn et al., 2007), age (Gallagher et al., 2008; Haze et al., 2001), ethnicity (Prokop-Prigge et al., 2016), diet and lifestyle factors (Krilaviciute et al., 2019; Qin et al., 2020), exercise (Finewax et al., 2021), social stress, fear and happiness (Preti et al., 2018; Smeets et al., 2020; Vautz et al., 2020; Williams et al., 2016), and sexual arousal (Wang et al., 2022). Even if results remain inconclusive, evidence about the chemical composition of these substrates in relation to individual traits and states is growing. Combined with appropriate behavioral studies, this will lead to an elucidation of the chemical underpinnings of human chemical communication in the coming years.
In addition to deciphering the qualitative nature of chemical cues, technical developments in the online analysis of exhaled breath (e.g., using online time-of-flight chemical ionization mass spectrometry; Bruderer et al., 2019; Vautz et al., 2020) could also be applied to skin emissions and will enable the tracking of rapid volatilome changes in response to real-life events (e.g., stressful school exams or physical exercises; Roberts, Misztal, & Langford, 2020). Moving away from periodic sampling at single time points, such continuous measurement will fill current knowledge gaps on the time course of signal transmission and receivers’ responses (Roberts, Misztal, & Langford, 2020; Wang et al., 2022).
A fuller understanding of the human volatilome also requires closer harmonization with approaches in microbiology because the microbiome (either on the skin or within the orodigestive and genitourinary tracts) determines the volatiles emitted by the body. A well-known example for the crucial role of the microbiome in body odorant formation relates to axillary emissions, which arise from the biotransformation of naturally secreted nonodorous precursor molecules by bacteria on the skin (Natsch & Emter, 2020). The recent finding of a key odor-forming enzyme conserved in specific armpit strains of haplorhine primates suggests an important role for the enzyme and related volatiles in communication among ancestral primates (Rudden et al., 2020). Microbiology can thus inform psychology by articulating precise hypotheses in the sense that certain axillary odorants may have been preserved for their original function (i.e., communication) or co-opted for a novel function (e.g., defense against colonization by pathogenic skin bacteria), or their original function has been lost, and actual production remains a nonfunctional vestige.
General physiological processes leading to the generation and secretion of semiochemicals and body odorants also remain to be elucidated. Notably, the contribution of genetic and hormonal systems to the formation of volatiles, secretory processes, and their intra- and interindividual variability, need to be understood. Such physiological processes are not only the basis for the generation of semiochemicals but also are important determinants of their detection (Kornbausch et al., 2022) and therefore deserve further research.
Resolving the question of human pheromones
Pheromonal communication in many invertebrate species is frequently independent of previous experience and elicits specific and apparently involuntary behaviors. Expecting the same in humans would seem to be a gross oversimplification, and no single body odorant has yet been shown to elicit a species-typical response. Instead, any pheromonal response, if it existed, might more likely be graded, conditional, and context-dependent, and any resulting behavior would likely include influences from various motivational systems (e.g., sexual, parental, and filial). Furthermore, the significance of learning mechanisms and response variability may vary across domains. For instance, odorant-guided breast-searching behavior in newborns may be relatively more stereotypic (Schaal et al., 2020) than olfactory influences on adult mate choice.
Should the term “pheromone” be therefore abandoned when considering humans? We do not think so. It is becoming clear that pheromonal communication, even in insects, may depend on internal or external conditions (e.g., Lenschow et al., 2018). A clearer working definition may help distinguish putative pheromones from other semiochemicals. Pheromones are signals that evolved for communication purposes sensu Maynard-Smith and Harper (1995). Therefore, one should be able to detect adaptive designs on both the sender and receiver sides. On the sender’s side, these include specialized scent glands, biochemical pathways for chemical production, context-dependent emission, and specific developmental or endocrine status. On the receiver’s side, one may expect specific neural pathways or receptor systems and optimized sensitivity to the signal in the functionally relevant context (specific developmental or endocrine status). It should be noted that the pheromone concept does not depend on a functional vomeronasal organ but can involve the main olfactory system, other chemosensory channels such as the Grüneberg ganglion (Brechbühl et al., 2008), or even direct contact (such as in kissing; see Wyatt, 2017). However, there is currently no evidence for a functional vomeronasal organ and Grüneberg ganglion in humans (Witt & Hummel, 2006).
From this basic theoretical framework, several predictions and research tasks can be derived regarding the chemistry and related behavioral aspects of pheromonal communication. We advocate the systematic framework described by Wyatt (2015, 2017). First, the pheromonal effect should be traced back to one or more identified chemical compound(s). Therefore, researchers should identify the qualitative chemical composition, relative quantities of the active compounds, and their functional threshold in the relevant body emission. Second, the specific compounds should be produced by virtually all individuals in a given state and specific context. For instance, a putative mammary pheromone would be produced by postparturient breastfeeding women but not (or much less) by other adult women or men or prepubertal young. Third, the compounds must be capable of being sensed and exerting their effect at the concentrations in which they are produced by the emitter. Psychophysiological examination of reactivity is required if the compounds are perceived subliminally. Finally, under relevant conditions, their perception should reliably induce a specific response in receivers, such as changing the prevalence of a specific behavior or triggering a physiological or developmental change. Note that we avoid using the term “release behavior” because the eventual behavioral change may depend on numerous other factors such as context, internal state, and previous experience. Nevertheless, a pheromone should systematically change the internal state of the perceiver, such as affective or attentional state, as predicted previously for informer pheromones (Müller-Schwarze, 1977). Researchers should therefore test relevant behavioral and/or psychophysiological responses with the putative pheromone compound and demonstrate they are comparable to those elicited by the original signal. Only if all these conditions are fulfilled should one speak of pheromonal communication for a given compound or mixture of compounds.
In contrast, body odorants may vary across individuals or groups and still have important communicative roles. These would include variation arising from different dietary customs that can influence how one’s body odor is perceived or idiosyncratic effects stemming from context-dependent associations acquired experientially; neither should be labeled pheromones. Likewise, individual-specific odor profiles, so-called signature mixtures, should not be termed pheromones, because by nature they vary greatly between individuals (Wyatt, 2017).
Benefits of chemical communication
One of the important future challenges in research on chemical communication is to understand the benefits it provides to humans, with attention on emitters as well as receivers. How does it enable friendship, mating, bonding, attachment, and adaptive self-regulation? How does it contribute to well-being and affect health-related psychology and behavior? A model of interest for answering these questions can be found in individuals with anosmia and hyposmia, which can affect approximately 20% of the population (Boesveldt, Postma, et al., 2017; Hummel et al., 2017). Indeed, when the sense of smell is absent (anosmia), reduced (hyposmia), or altered (parosmia), social interactions are reduced (Desiato et al., 2021), probably because of modifications in attitudes related to both one’s own and another person’s body odor (Croy, Nordin, & Hummel, 2014). Feelings of insecurity regarding the monitoring of one’s own body odor (sweat, breath), sometimes associated with compulsive personal-hygiene behaviors, are added to the conscious missing of the odor of loved ones. A possible consequence is a disruption of social interactions through a breach in attachment or sexual-attraction processes (Croy, Nordin, & Hummel, 2014).
Such consideration of the effects on social functioning can also extend into individual variation in typical olfactory performance. Two studies reported better olfactory functioning in socially integrated individuals (Boesveldt, Yee, et al., 2017; Zou et al., 2016). It is suggested that the higher social integration might be based on skills related to empathic behavior. Indeed, at least in women, higher olfactory performance scores are related to higher scores in empathic perspective taking (Lübke et al., 2022; Zhou & Chen, 2009b). Other studies showed a higher degree of agreeableness in relation to lower olfactory thresholds (Croy et al., 2011). In line with Bastir et al. (2011), it is suggested that expanded olfactory capabilities might have been engaged in the evolution of social behavior in humans (Lübke et al., 2022).
Recent smell loss as an indicator of COVID-19 infection (Parma et al., 2020) has made the importance of the sense of smell more tangible to people in general, with a clear impact on individual well-being (e.g., Burges Watson et al., 2021; Yom-Tov et al., 2021). Therefore, deploying rehabilitation protocols in individuals with olfactory dysfunction seems a promising way to reduce social-isolation processes that result from olfactory loss (Croy et al., 2012). Indeed, olfactory training (daily exposure to certain odorants over several weeks) has positive effects on olfactory abilities (Sorokowska, Drechsler, et al., 2017) and symptoms of depression (Kamrava et al., 2021) and may thus, in the longer term, have indirect behavioral benefits. Wegener et al. (2018) gave weight to this idea by showing that several weeks of olfactory training in elderly people improves not only olfactory abilities but also cognitive performance and well-being. It remains to be explored whether such an intervention can act on the social sphere and individual well-being.
In addition, there is emerging evidence that the positive emotion of one individual can be evoked in others through chemical cues (Ortegón et al., 2022; Smeets et al., 2020). In the future, principles of chemical communication could thus be used to regulate individual emotional states, such as reducing anxiety and depression by specific olfactory safety cues. The interlinked neural wiring of emotion- and olfaction-processing areas (Schablitzky & Pause, 2014) may give further guidance for depression treatments. Depressed individuals have reduced olfactory functioning and processing (Pause et al., 2003) and potentially top-down attention mechanisms (Laudien et al., 2006), functional state of the olfactory bulb (Pause et al., 2003), and proliferation rates of the olfactory sensory neurons (Croy & Hummel, 2017). Because electrical stimulation of the olfactory epithelium alters the neural interplay in deep brain structures (Weiss et al., 2016), targeted brain-stimulation techniques via the olfactory channel are currently under exploration. Another important question to answer is how to make use of the knowledge regarding chemical communication of physiological and psychological conditions. Examples in this respect are the development of monitoring systems for preventive medicine based on chemical information, including sweat-sensing devices to detect, for example, COVID-19, among other infections (e.g., Snitz et al., 2021; Ibrahim et al., 2022).
Finally, as in many scientific fields, applications can be controversial and are not always seen as positive. For example, body odorants might potentially be accessed to obtain sensitive information about individuals (e.g., on identity, health, drug use) or used to control the behavior of individuals or crowds, or as nonlethal weapons. Such applications could initially be used to achieve aims that are accepted by and of indisputable utility for society, such as the use of volatile fingerprinting in forensic sciences (Cuzuel et al., 2017) or early detection of disease (Regenbogen et al., 2017), but then later abused. Scientists and societies will need to reflect prospectively on the ethics of potential applications and their possible consequences.
Fostering interdisciplinary research strategies and providing theoretical frameworks
Future progress in understanding human chemical communication will be possible only with inputs from different disciplines, including chemistry, biology, psychology, and anthropology. Within neuroscience, special attention is now directed to the impact of chemical cues on overt behavior, such as olfactory sampling via sniffing (Frumin et al., 2015), because the perception of chemical cues is likely to be an implicit rather than explicit process, and individuals might be unaware of the presence or effects of chemosensory stimuli (Degel et al., 2001). Input from the social sciences and other disciplines will help to increase the ecological validity of methods, with behaviors being observed in genuine social interactions (Meister & Pause, 2021) outside the laboratory or by using alternatives such as virtual interactions with avatars in immersive contexts (Kim et al., 2015). Using models of social interactions (Xiao et al., 2016) may highlight both the importance of chemical communication in intergroup relations and understanding how group belonging (e.g., cultural, gender, age groups) influences chemical communication. Concepts used in social cognition research, such as gender stereotypes, expectations regarding body odors (Mutic et al., 2015), and related cultural practices (Majid, 2021), also constitute useful frameworks for studying the variability of human chemical communication. However, developing such interdisciplinary programs is no mean feat (Kimball, 2016). It will require international consortia conjoining disciplines, with a strong focus on building systematic, open-access databases (de Groot et al., 2020; Roberts, Havlíček, & Schaal, 2020) and innovative methods for the analysis of complex datasets, including machine- and deep-learning approaches.
Interdisciplinary efforts also appear to be necessary to further develop theoretical concepts and frameworks of chemical communication. Future research involving theoretical approaches to communication, insights from related areas such as visual or auditory communication, and fundamental biological and chemical research will help to characterize and categorize the different types of chemical interactions between humans, referring to parameters such as the underlying receptive systems, the adaptive value of the chemocommunicative process, or the type and plasticity of the response. Such research may finally lead to more differentiated concepts of chemical communication. In this respect, it may also be helpful to distinguish between communication via odors (e.g., a malodor related to a disease state or a wound, which would be explicit) versus communication via odorless chemicals or chemicals that are below odor thresholds (e.g., below-threshold fear odor, which would be implicit). A major challenge in this respect is that, at the moment, the principles underlying many of the described cases of communication are not yet fully characterized.
So far, human chemical communication research has been dominated by applying approaches from experimental and social psychology to validate in humans what has been traditionally viewed as a form of social communication reserved to the animal kingdom and understood in terms of theories strongly rooted in biology. Close alignment between biological and psychological theories of social communication, with a future scientific roadmap as illustrated above, will enable the scientific community to benefit from either perspective to arrive at an appraisal of the nature, impact, and relevance of chemical communication to humankind.
Changing societal views on human body odor
The nature and functions of human body odors is a topic that fascinates not only scientists but also both the general public and industries that manufacture personal and home-care products, including cosmetics and fragrances. However, awareness of the communicatory function of smell, and the opportunities this affords, remains low. The scientific community might join forces with manufacturers to move beyond their central aim of eradicating malodor, if we can better understand the interactions between fragranced cosmetics and naturally produced body odor that forms personal odor (Havlíček & Roberts, 2013; Lenochová et al., 2012). Researchers need to engage to better explain the functioning of human chemical communication and dispel misleading claims of proven pheromones (Wessel, 2017). They might also engage the public in citizen science via questionnaires and systematic recording of attention to body odor in their daily lives (Ferdenzi et al., 2008; Li et al., 2022; Schaal, 1996), which will likely present new and interesting perspectives.
Finally, we believe that, based on cumulative evidence from research over the past decades into the human capability for chemical communication, we have arrived at a point at which it has earned its place in mainstream psychology, where it can be fully integrated in comprehensive social communication theories that appreciate the contributions of all sensory channels. Consequently, we are now able to understand that lack of chemosensory input can affect our social experiences and well-being in ways that psychologists have not always considered. This may be especially important in a world in which our social interactions are increasingly online.
Conclusions
Here we described possible historical reasons for the scientific neglect of the human sense of smell in general, and chemical communication in particular. This view has started to slowly change, partly because of recent empirical findings on the role of chemical cues in various social interactions. In particular, there is now robust evidence that chemical cues play a critical role in mother-infant interactions (Schaal et al., 2020), affect mate choice (Ferdenzi et al., 2019) and romantic relationship functioning (Mahmut & Croy, 2019), and are involved in communication of affective states (Lübke & Pause, 2015).
Our aim was not only to review current evidence for human chemical communication but also to promote better theoretical foundations for psychology and beyond. For instance, a more structured understanding was long precluded by unrealistic expectations about human pheromones. The idea of a chemical compound triggering a stereotypic reaction, irrespective of contextual factors, might attract major media attention but seems unlikely (at least in adults) given what we currently know about human cognitive and behavioral functioning.
An important limitation of current research is a lack of concrete knowledge about the chemical compounds responsible for observed biopsychological effects. The human body emits hundreds of compounds, some highly variable over time, and detecting and assessing the concentrations responsible for eliciting key behavioral effects is not an easy task. Nevertheless, major breakthroughs may be forthcoming because of the increasing availability of advanced techniques that enable the measurement of dynamic changes in volatiles and the identification of active compounds in normally behaving individuals. Another limitation of current research is its dependence on laboratory work on WEIRD and especially ODD samples; our understanding of social odors in other societies is extremely limited (Roberts, Havlíček, & Schaal, 2020). There is a pressing need to conduct fieldwork in non-ODD societies to understand the uses of social odors in different cultural settings rather than making sweeping claims about humankind on the basis of only small samples from a single society (Henrich et al., 2010). Clearly, psychologists have a central role to play in these efforts, but bridging remaining knowledge gaps will only be possible through collaborative efforts with scholars from different fields, including analytical chemistry, microbiology, behavioral sciences, medicine, and anthropology. We hope that the roadmap we propose here will facilitate such research in the near future.
Glossary
Terms related to physicochemistry
Terms related to chemosensation
Terms related to communication
Footnotes
Acknowledgements
We would like to express our gratitude to Arnaud Fournel and Dagmar Schwambergová for creating the figures.
Transparency
Action Editor: David A. Sbarra
Editor: Interim Editorial Panel
Author Contributions
H. M. Loos, B. Schaal, B. M. Pause, M. A. M. Smeets, C. Ferdenzi, S. C. Roberts, J. de Groot, K. T. Lübke, I. Croy, J. Freiherr, and J. Havlíček drafted the manuscript. H. M. Loos, B. S Schaal, B. M. Pause, M. A. M. Smeets, and J. Havlíček coordinated the writing of individual sections and the overall text. All authors contributed to the conception and outline of the manuscript, were involved in the review and editing, and approved the final manuscript for submission.