
Abstract
Trigeminal neuralgia (TN) is a debilitating neuropathic facial pain disorder characterized by sudden, severe paroxysmal pain that markedly impairs quality of life. Although current pharmacological treatments provide pain relief, their adverse effects highlight the need for safer alternative therapies. Curcumin, a natural polyphenol, exhibits anti-inflammatory and neuroprotective properties with minimal side effects. In this study, we examined the impact of intraperitoneal curcumin administration on the trigeminal ganglia in a TN rat model induced by unilateral infraorbital nerve constriction (IONC). Behavioral tests revealed that curcumin significantly reduced mechanical hypersensitivity compared with vehicle-treated controls. Immunohistochemical analysis showed decreased expression of glial fibrillary acidic protein (GFAP) and activating transcription factor 3 (ATF3), established markers of satellite glial cell (SGC) activation and neuronal stress in the trigeminal ganglia. Furthermore, curcumin administration reduced tumor necrosis factor-α (TNF-α) expression and modulated phosphorylated signal transducer and activator of transcription 3 (pSTAT3) activity. These findings suggest that curcumin alleviates neuropathic pain by suppressing glial activation and pSTAT3-related pro-inflammatory signaling. Curcumin may therefore represent a promising potential therapeutic candidate for TN, warranting further investigation into its clinical applicability and underlying mechanisms.
Keywords
Introduction
Neuropathic pain, as defined by the International Association for the Study of Pain, is pain caused by a lesion or disease affecting the somatosensory nervous system, encompassing syndromes such as trigeminal neuralgia (TN), one of the most debilitating facial pain disorders.1,2 The trigeminal ganglion (TG) contains sensory neurons of the trigeminal nerve, which are essential for transmitting sensory information from the orofacial region and play a critical role in the pathophysiology of TN.3,4 Clinically, TN is characterized by sudden, intense, and electric shock-like pain conditions in one or more branches of the trigeminal nerve, most commonly resulting from neurovascular compression at the trigeminal root entry zone, leading to demyelination and dysregulation of voltage-gated sodium channels.5,6
Recent research highlights the importance of glial fibrillary acidic protein (GFAP) as a marker of activated satellite glial cells (SGCs) within the TG. 7 Peripheral nerve damage, such as chronic infraorbital nerve constriction, initially induces the overexpression of GFAP in astrocytes of the trigeminal nucleus caudalis (TNC), a second-order relay center in the brainstem, that corresponds with mechanical allodynia and thalamic hyperexcitability. 8 This glial activation facilitates intercellular communication among neurons in TG, reflecting a systemic response to localized injury. 9 The key mediators in the pathogenesis of TN include proinflammatory cytokines, particularly tumor necrosis factor-α (TNF-α) and interleukin-6 (IL-6). 10 Activated microglia and astrocytes in the spinal trigeminal nucleus simultaneously secreted IL-6 and TNF-α, thereby sustaining neuroinflammation and central sensitization. 11 GFAP-positive glial cells and cytokines interact in a pathological feed-forward loop, in which glial activation promotes cytokine release, further aggravating neural hyperexcitability and enhancing glial reactivity.8,11,12
ATF3 and GFAP are essential molecular markers that reflect distinct yet interrelated responses to nerve injury in the TG, significantly contributing to the pathophysiology of neuropathic pain.13,14 ATF3 is specifically upregulated in the nuclei of injured sensory neurons following peripheral or central nerve injury, serving as reliable indicator of neuronal stress and dysfunction. Simultaneously, GFAP is markedly elevated in SGCs adjacent to these neurons, indicating glial activation and the initiation of neuroinflammatory processes. 14 The concurrent upregulation of ATF3 and GFAP indicates a synergistic neuron-glia interaction that contributes to the maintenance of trigeminal pain. The pathological condition is further exacerbated by the release of proinflammatory cytokines, including TNF-α, which intensify the inflammatory response and enhance neuronal sensitization. This interaction establishes a feed-forward loop that amplifies nociceptive signaling.14,15 ATF3 and GFAP thus serve as critical markers of neuronal and glial activation, respectively, and represent key components of the neuroimmune axis implicated in the chronicity of TN.
First-line pharmaceutical therapies, such as carbamazepine and oxcarbazepine, aim to modulate neuronal activity by targeting voltage-gated sodium channels, whereas in refractory cases, surgical interventions, such as microvascular decompression may be required.6,16 However, both carbamazepine and oxcarbazepine are associated with prevalent side effects, including drowsiness, dizziness, and nausea, along with more severe risks affecting systemic conditions.17,18 Cognitive impairment and memory deficits are also frequently reported. 18 A deeper understanding of the molecular and cellular mechanisms underlying TN, including peripheral and central sensitization in TG, is essential for enhancing current therapeutic approaches.3,16 Thus, exploring and developing alternative treatments for TN that provide comparable efficacy with improved safety and tolerability for TN profile is needed.
Curcumin, a polyphenolic substance derived from the rhizome of Curcuma longa, has attracted considerable scientific attention owing to its potent anti-inflammatory effects and extensive medical potential.19,20 Extensive research, including both preclinical and clinical investigations, has demonstrated that curcumin exerts its anti-inflammatory benefits via multiple molecular mechanisms. 21 These include downregulating proinflammatory cytokines, including TNF-α and IL-6, through several pathways and one of which is STAT3 pathway. 22 Comprehensive reviews and research articles have demonstrated that phosphorylated signal transducer and activator of transcription 3 (pSTAT3) is significantly upregulated in various models of neuropathic pain, including chronic constriction injury through this pathway.23,24 Numerous animal studies and clinical trials have reported that curcumin supplementation reduces inflammatory mediators and improves disease outcomes, suggesting its potential as an adjuvant or alternative therapy for managing chronic inflammation.25,26 Despite its favorable pharmacological profile, the clinical application of curcumin is limited by its low bioavailability, highlighting the need for continued research into novel formulations and structural analogs to enhance its therapeutic efficacy. 27
The growing research emphasizes curcumin’s versatility as a potent anti-inflammatory agent with significant potential in the management of various inflammatory diseases.28–30 Therefore, the present study was designed to investigate the effect of curcumin on specific molecular changes in the TG of TN animal model.
Materials and methods
Animals
Four-week-old healthy male Sprague Dawley rats (weighing 80–100 g) were obtained from CLEA Japan, Osaka, Japan, and housed in polycarbonate cages with shredded wood tip bedding to ensure comfort and well-being. To replicate natural conditions, the animal facility was maintained at a regulated temperature of 20–24°C with a 12 h light/dark cycle. The rats were provided unrestricted access to a standard laboratory diet and fresh water. All experimental procedures were conducted in accordance with the institutional guidelines for animal care and use. This study is reported in accordance with the ARRIVE guidelines 31 and adheres to the ethical recommendations for pain research in animals published by the International Association for the Study of Pain (IASP). At the end of the experiment, rats were deeply anesthetized with isoflurane until loss of reflexes. Euthanasia was then performed by decapitation using a sharpened guillotine. This method was selected to obtain TG tissue uncontaminated by chemical anesthetics. The animal protocols were approved by Tokushima University (T2020-108, T2023-90).
TN model
Four experimental groups – naïve, IONC, IONC + vehicle, and IONC + curcumin – were assigned randomly to rats. For rats undergoing IONC, unilateral infraorbital nerve constriction was performed via an intraoral approach targeting the infraorbital branch of the trigeminal nerve (maxillary nerve). The rats were deeply anesthetized using a combination of 0.375 mg/kg medetomidine hydrochloride (Nippon Zenyaku Kogyo Co., Ltd., Japan), midazolam (Sandoz K.K., Yamagata, Japan) 2 mg/kg, and 2.5 mg/kg butorphanol (Meiji Seika Pharma Co., Ltd., Japan). A precise intraoral incision was made through the hard palate, extending about 1 cm anteriorly toward the incisors. The nerve was carefully exposed, and two loose ligations (4-0 silk suture) were placed around it to induce constriction. Following the procedure, the incisions were securely closed with mucosal adhesive (3M™ Vetbond Tissue Adhesive, USA).
Mechanical hypersensitivity measurement
The mechanical hypersensitivity measurement was done as previously described.14,32 One day before the behavior test, the bilateral orofacial area of each rat was shaved using clippers. Behavioral assessments were conducted by an examiner blinded to the experimental conditions to minimize bias. During testing, each rat was placed inside a holder (Durham Holders, 37100, Ugo Basile, Verase, Italy) with its snouts protruding through a semicircular small opening, allowing free movement of the head. Mechanical stimulation was delivered to the designated whisker pad area using an electronic von Frey aesthesiometer (Model 1601C, IITC Instruments, Woodland Hills, CA, USA) equipped with a rigid plastic tip (0.8-mm diameter), capable of measuring forces up to 800 g. The center of the rat’s snout was the stimulation point within the infraorbital nerve’s territory. Stimuli were applied until the rat showed signs of withdrawal response. Each rat underwent stimulation on both the ipsilateral and contralateral sides with five times per side at 1-min intervals. Three median force values were averaged after excluding the highest and lowest measurements. The applied force (g) was recorded at the time of withdrawal. The behavior assessments were conducted on the day before IONC surgery, 7 days post-operative, and in every 2 h after drug administration.
Drug administration
Freshly prepared curcumin (C1386, Sigma-Aldrich, USA) was dissolved in a vehicle of 100 µl of 0.5 M sodium hydroxide (NaOH) in PBS and diluted again in PBS until 1.5 ml final volume, yielding a final pH of 8. A dose of 45 mg/kg of curcumin was given to each rat. Intraperitoneal administration of curcumin was administered in IONC + curcumin group. Meanwhile, in the IONC + vehicle group, only 0.5 M NaOH in PBS was administered. The drug administration was conducted on the seventh day post-IONC, while the naïve and IONC groups did not receive any intervention on that day.
Immunofluorescence staining
The extracted TGs were collected on the seventh day at 4 h after drug administration and then preserved in 4% paraformaldehyde (PFA). After fixation, the tissues underwent sequential cryoprotection in 10% and 20% sucrose solutions. The tissues were embedded in Tissue-Tek O.C.T Compound (Sakura Finetek USA Inc, Torrance, CA, USA) and stored at −20°C until cryosectioning. The TGs were sliced horizontally into 5–10 µm thickness along the long axis. The slices were air-dried at room temperature and subsequently washed in 0.25% Triton-X 100 in PBS two times for 5 min each. The sections were blocked with 5% Block One (03953-95, Nacalai, Japan) for 30 min to prevent non-specific binding. This was followed by overnight incubation at 4°C with anti-GFAP polyclonal antibody (1:500; Bioss Antibody, Boston, MA, USA), anti-ATF3 monoclonal antibody (1:500; Santa Cruz, CA, USA) and anti-phospho-STAT3 monoclonal antibody (1:100; Cell Signaling Technology, Denver, MA, USA). The sections were incubated for 2 h at room temperature the following day using Alexa Fluor 488 anti-rabbit IgG (1:200; Thermo Fisher Scientific, Waltham, MA, USA). Followed a thorough rinse, the sections were mounted using antifade mountant ProLong™ Diamond Antifade Mountant with DAPI (P36981, Thermo Fisher Scientific, USA). Imaging was performed using BX 800 fluorescence microscope (Keyence, Osaka, Japan). To examine the intensity and quantify the immunoreactivity cells in the TG, three nonadjacent TG sections were randomly selected and measured with ImageJ.
Quantitative real-time polymerase chain reaction (qRT-PCR)
TGs were collected on 4 h after drug administration on seventh day after IONC and promptly preserved in RNAlater (Sigma Aldrich, St. Louis, MO, USA) according to the manufacturer’s instructions. Naïve rats were used as controls. A tissue lyser was used to homogenize the tissues samples, and total RNA was extracted from the TG using trizol (Invitrogen, Carlsbad, CA, USA). The extracted RNA was then reverse transcribed to cDNA using a high-capacity RNA-to-cDNA kit (Applied Biosystems, Foster City, CA, USA) following the manufacturer’s instructions. qRT-PCR was performed using gene-specific primers. The sequences were as follows: β-actin (forward 5′-AAGTCCCTCACCCCTCCCAAAG-3′, reverse 5′-AAGCAATGCTGTCACCTTCCC-3′), 33 TNF-α (forward 5′-CCCCGACTATGTGCCCTCAC-3′, reverse 5′-AGGGCTCTTGATGGCGGA-3′), 34 IL-6 (forward 5′-GGGACTGATGTTGTTGACAGCC-3′, reverse 5′-CATATGTAATTAAGCCTCCGACTTGTG-3′), 34 and GFAP (forward 5′-AGTGGTATCGGTCCAAGTTTGC-3′, reverse 5′-TGGCGGCGATAGTCATTAGC-3′) 35 using Fast SYBR Green Master Mix (4309155, Applied Biosystems, USA) in Quantstudio 3 system (Applied Biosystems, Foster City, CA, USA). The amplification was conducted using QuantStudio 3 RT-PCR System (A28567, Thermo Fisher Scientific, USA), and relative gene expression was normalized to β-actin using the 2−ΔΔCt method.
Cytokine assay
Cytokine expression in TG was analyzed using the Proteome Profiler™ Rat Cytokine Array Panel A (R&D Systems, Minneapolis, USA), following the manufacturer’s protocol. In this study, TGs from all three division were collected 4 h after intraperitoneal injection of either curcumin or vehicle on day 7 following IONC. Excised TGs were washed with cold PBS, finely minced, and homogenized in a lysis buffer composed of 80 μl distilled water, 10 μl RIPA buffer, and 10 μl sodium dodecyl sulfate. Lysates were kept on ice for 30–60 min and then centrifuged at 10,000 ×g for 10 min at 4°C to remove cellular debris. The supernatant was collected, and total protein concentration was measured using a BCA protein assay (23227, Thermo Fisher Scientific, USA). Equal amounts of protein from each sample were incubated with a biotinylated detection antibody cocktail and applied to pre-spotted nitrocellulose membranes. After overnight incubation at 4°C, membranes were washed and incubated with streptavidin–horseradish peroxidase, followed by chemiluminescent substrate. Chemiluminescent signals were captured with the ImageQuant™ system (GE Healthcare) and analyzed densitometrically using ImageQuant TL software. Spot intensities were quantified with the same software. Signals were background-subtracted and normalized to internal reference spots. Results were expressed as relative fold changes in curcumin-treated compared to vehicle-treated TGs.
Statistical analysis
Data are expressed as means ± standard errors of the mean (SEM). Statistical analyses were conducted using IBM SPSS Statistics for MacOS, Version 27.0 (IBM Corp., Armonk, NY, USA). A 2-h interval behavior test was measured with repeated two-way ANOVA. The 4 h behavior test on all groups, qRT-PCR and immunofluorescence staining result were measured with one-way ANOVA. A p-value of <0.05 was considered statistically significant for all group comparisons.
Results
The behavior changes after curcumin administration
Pain behavior, as reflected by mechanical threshold in rats subjected to IONC, was significantly reduced on the seventh post-operative day compared with their baseline values, indicating that the IONC procedure effectively established and simulated TN. On day 7, rats received either curcumin or vehicle, and behavioral changes were monitored every 2 h (h). As shown in Figure 1, the IONC + curcumin group exhibited significantly higher threshold than the IONC + vehicle group at 2 h (p = 0.024), 4 h (p = 0.007), and 6 h (p = 0.001) post-injection, with the peak effect observed at 4 h.

Reduction of pain sensitivity after curcumin administration. Baseline values were taken before IONC surgery. Seven days after the surgery, the pain threshold in both groups decreased compared with their baseline value. Injections were performed following 0 h of behavior measurements.
Effects of curcumin on mechanical hypersensitivity in TN
Behavioral tests were assessed every 2 h following drug administration. A peak effect was observed at 4 h post-injection across the experimental groups. At this time point, the IONC + curcumin exhibited a significantly higher pain threshold compared with its pre-injection value (p < 0.001), the IONC only group (p < 0.001), and IONC + vehicle group (p < 0.001). The significant increase in threshold demonstrates that curcumin markedly alleviated pain hypersensitivity in rats with IONC-induced neuropathic injury (Figure 2).
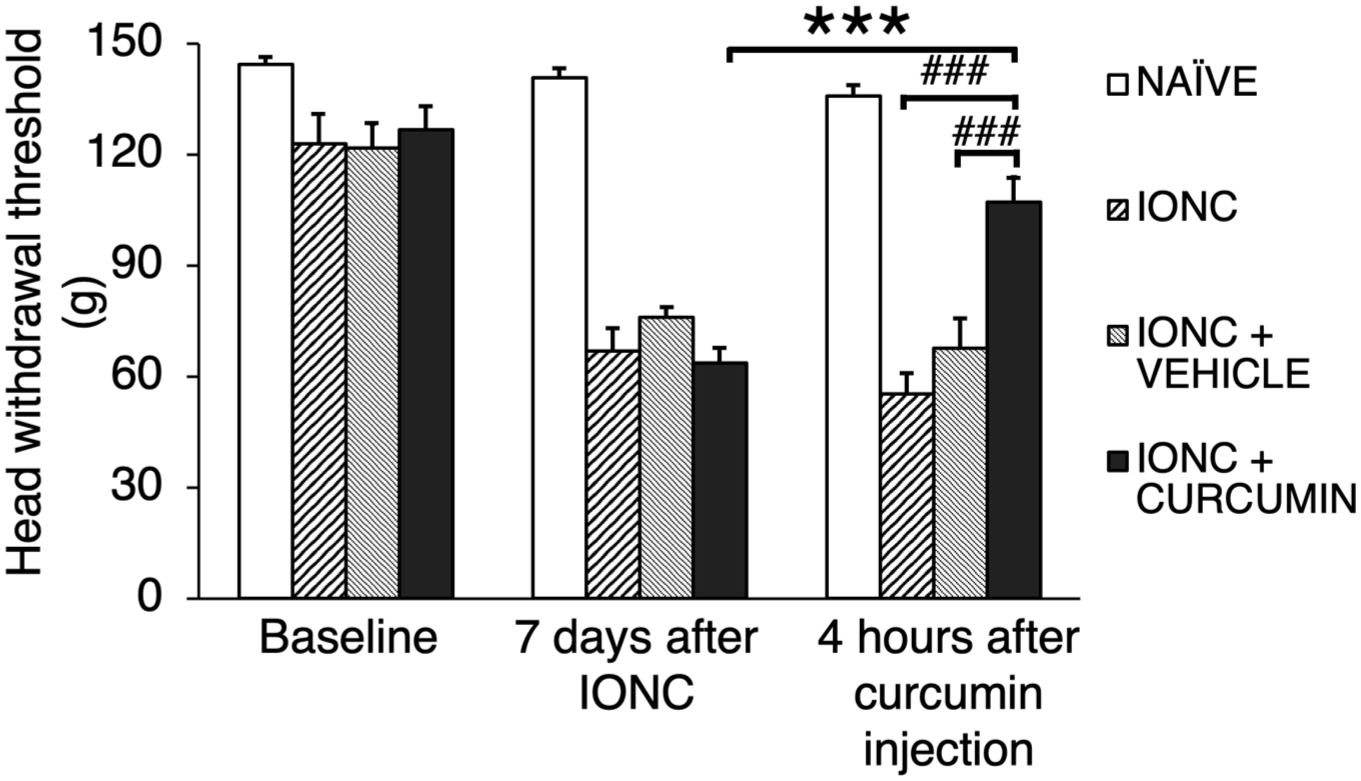
Mechanical threshold analysis across all groups. Baseline value were taken before IONC surgery. Seven days after the surgery, the pain threshold in all IONC-treated groups decreased compared with their baseline value. Curcumin was administered after performing a behavior test 7 days post-IONC. Curcumin treatment noticeably had a significant higher pain threshold compared to its pre-injection value and the other IONC-treated groups 4 h after injection.
Curcumin reduced GFAP expression in the TG of TN rats
GFAP is a well-established marker of glial cell activation. 36 SGCs express GFAP at low levels under resting conditions; however, its expression is markedly upregulated in response to injury. 37 In the present experiment, the pictures were taken from V2 area and showed glial cell activation led to changes in GFAP protein (Figure 3(a) and (b)) and mRNA expression 4 h post-injection (Figure 3(c)). Immunohistology analysis using corrected total cell fluorescence (CTCF) revealed significantly higher GFAP-positive cell intensity in TG of IONC (p < 0.001) and IONC + vehicle (p < 0.001) rats compared to the IONC + curcumin group 4 h post-injection (Figure 3(b)). Additionally, while changes in mRNA expression were detectable, they did not reach statistical significance (vs IONC, p = 0.123; vs IONC + vehicle, p = 0.106; Figure 3(c)).

GFAP expression across all groups. (a) The immunoreactivity of GFAP in IONC + curcumin group is decreased in the rat trigeminal ganglia after injection of curcumin (GFAP: green, DAPI: blue). (b) The intensity of GFAP is measured using corrected total cell fluorescence and showed a significantly lower intensity in IONC + curcumin group compared to IONC and IONC + vehicle groups. (c) mRNA expression of GFAP decreases after curcumin administration.
Curcumin reduced ATF3 expression in the TG of TN rats
ATF3, an established marker of neuronal activation, is usually undetectable in normal, healthy neurons (Supplemental Figure S1). 38 By contrast, ATF3 transcription is rapidly upregulated and localized within the neuronal nucleus following peripheral nerve injury, as in the IONC model, reflecting a cellular stress response. 39 In this study, ATF3 expression was markedly increased in TG neurons after IONC induction, consistent with previous investigations (Figure 4(a)). Importantly, curcumin administration significantly decreased the proportion of ATF3-positive neurons (vs IONC, p = 0.002; vs IONC + vehicle, p = 0.007; Figure 4(b)). This reduction in ATF3 immunoreactivity indicates that curcumin attenuated neuronal stress signaling, consistent with the observed improvement in pain behaviors in IONC-injured rats.

ATF3 expression across all groups. (a) The immunoreactivity of ATF3 in IONC + curcumin group is decreased in the rat trigeminal ganglia after injection of curcumin (ATF3: red, DAPI: blue). (b) The number of positive cells compared to total cells shows that the ATF3 expression is decreased on curcumin injection group.
Curcumin reduced TNF-α but not IL-6 expression
Quantitative real-time PCR (qRT-PCR) of TG tissue extracts was performed for each of experimental groups to assess relative mRNA levels of the pro-inflammatory cytokines Tnf-α and Il-6. Tnf-α expression was significantly decreased in the IONC + curcumin group compared to IONC (p = 0.016) and IONC + vehicle groups (p = 0.031), indicating that curcumin attenuates the injury-induced increase in Tnf-α (Figure 5(a)). In contrast, Il-6 expression did not differ significantly among the IONC (p > 0.05) and IONC + vehicle (p > 0.05) groups compared with the IONC + curcumin group (Figure 5(b)).

Gene expression of Tnf-α and Il-6. (a) Tnf-α expression in TG of TN rats receiving curcumin was significantly lower compared to IONC and IONC + vehicle group. (b) The Il-6 expression remained unchanged in all groups.
Curcumin differentially regulated cytokines after nerve injury in the TG
An antibody array including 29 different cytokines and chemokines revealed that curcumin administration in the chronic constriction injury model suppressed the expression of several inflammatory mediators. Four hours after curcumin treatment, multiple pro-inflammatory factors were reduced compared with the vehicle-treated therapy (Supplemental Figure S2). In particular, a substantial decline in the chemokine LIX (CXCL5) was observed, suggesting that this signaling molecule was rapidly modulated. These results demonstrate that curcumin rapidly alerts the inflammatory milieu in the injured TG by reducing CXCL5 level, thereby potentially targeting the chemokine-driven pain pathway early after nerve injury (Table 1).
Average fold change in the level of cytokine release in TG on TN rats after curcumin injection compared to vehicle injection.

The level of some cytokines was less than 1 fold, with the lowest being LIX (CXCL5).
PSTAT3 subcellular localization in TG neurons and SGCs
Immunofluorescence labeling for pSTAT3 was performed to examine the underlying signaling processes. As shown in Figure 6, the naïve group exhibited negligible pSTAT3 expression, whereas the IONC and IONC + vehicle groups displayed markedly elevated pSTAT3 levels, indicating activation of STAT3 pathway. In contrast, the IONC + curcumin group showed reduced pSTAT3 fluorescence signals, suggesting suppression of this pathway. These findings are consistent with previous reports implicating pSTAT3 activation in neuropathic pain models. 40 Given the involvement of STAT3 signaling in both peripheral and central nervous systems, curcumin-mediated suppression of this pathway may also contribute central pain processing.

pSTAT3 expression across all groups. The immunoreactivity of pSTAT3 in IONC +curcumin group is less compared to IONC and IONC + vehicle group (pSTAT3: green, DAPI: blue). Scale bar 50 µm.
Discussion
This study demonstrates that systemic administration of curcumin alleviates mechanical hypersensitivity in rats with IONC-induced TN. Following a single intraperitoneal injection, withdrawal thresholds were significantly increased at 2 h, and remained elevated until 6 h post-injection (Figure 1). The rapid onset and sustained effect suggest that curcumin modulates early pain-processing pathways, likely through suppression of inflammatory signaling within the TG. Compared with both untreated IONC and vehicle-treated groups, curcumin produced a more robust antinociceptive response, supporting its therapeutic potential. These behavioral results are consistent with previous reports highlighting the contribution of early neuroimmune interactions to the initiation and persistence of neuropathic pain, and reinforce the concept that targeting inflammatory mediators immediately after nerve injury may be an effective treatment strategy. 41
At the cellular level, curcumin reduced both GFAP and ATF3 – established markers of SGC activation and neuronal stress, respectively – in TG tissue 4 h after treatment compared with vehicle controls. GFAP, typically upregulated in reactive SGCs, and ATF3, expressed in neuronal nuclei under stress, both showed early suppression. This indicates that curcumin dampens glial activation and indirectly reduces neuronal excitability by disrupting neuron-glia signaling. These results align with evidence that SGC-derived proinflammatory mediators enhance nociceptive transmission in chronic pain models, underscoring glial reactivity as a therapeutic intervention.40,42
Curcumin also correlates with downregulation of Tnf-α mRNA expression without affecting Il-6 levels. This specificity suggests a targeted inhibitory effect on TNF-α mediated signaling rather than broad cytokine suppression. In the TG, Tnf-α functions as a crucial mediator of bidirectional communication between sensory neurons and adjacent glial cells, stimulating excitatory neurotransmitter release and promoting glial proliferation. 43 Curcumin selectively reduced Tnf-α expression, thereby disrupting pathological feedback loops that drive neuronal hyperexcitability and glial activation. 44 These findings are consistent with previous studies reporting that curcumin attenuated neuroinflammation by decreasing TNF-α levels in the hippocampal region of neuropathic pain model. 45 By contrast, Il-6 may contribute at later stages or act through distinct cellular targets, such as infiltrating macrophages or endothelial cells. 46
Protein-level profiling using cytokine arrays further confirmed curcumin’s anti-inflammatory effects. Among the 29 cytokines assessed, CXCL5 exhibited the most pronounced downregulation. CXCL5 has been reported to amplify pain-related immune responses by promoting leukocyte recruitment and upregulating pSTAT3 through cytokine-driven JAK/STAT signaling. 47 Consistent with prior report, 48 our results demonstrate that curcumin reduces CXCL5 expression in neuropathic pain, suggesting that the behavioral effects observed here are closely linked to local cytokine suppression within the TG rather than systemic immune modulation alone.
The molecular profile observed in this study implicated the STAT3 pathway, a central regulator of proinflammatory signaling. STAT3 activation has been linked to TNF-α production in chronic inflammatory disorders such as rheumatoid arthritis and atopic dermatitis,49,50 and is also involved in renal pathology, cancer, and cardiovascular responses.51–53 Importantly, curcumin has been shown to inhibit STAT3 signaling in neurological injury models, where it reduces neuroinflammation and tissue damage. 54 In line with these findings, our data indicate an association between curcumin treatment, reduced pSTAT3 levels in the trigeminal system, and lower neuropathic pain behaviors, along with decreased TNF-α-driven neuroimmune interactions.
Curcumin is unlikely to directly eliminate pre-existing GFAP or ATF3 proteins or TNF-α mRNA within 4 h in the trigeminal ganglion; instead, the observed changes are more plausibly explained by curcumin’s rapid suppression of upstream STAT3-dependent inflammatory signaling, which is well documented in neural injury and neuropathic pain models.40,55,56 In the spinal cord, curcumin inhibits JAK2-mediated STAT3 phosphorylation, a pathway known to drive reactive astrogliosis and GFAP transcription, suggesting that reduced STAT3 activation can quickly diminish ongoing GFAP synthesis.40,55 Curcumin also modulates STAT3-linked stress and survival pathways in neural and tumor systems. 52 Importantly, curcumin has also been shown to down-regulate elevated ATF3 expression in a chronic endoplasmic-reticulum–stress mouse model by attenuating upstream stress signaling, indicating that reductions in ATF3 can occur rapidly when the sustaining stress pathways are suppressed. 57 Taken together, these data support the interpretation that the 4-hour reduction in GFAP, ATF3, and TNF-α expression observed in our model most likely reflects a rapid decline in upstream transcriptional drive – particularly within STAT3-related pathways – rather than direct clearance of previously elevated proteins or mRNA.
The behavioral and molecular results support a model in which curcumin interrupts the STAT3-dependent neuroimmune feed-forward loop that sustains peripheral sensitization. The selective suppression of TNF-α and CXCL5, but not IL-6, highlights pathway specificity that may be therapeutically advantageous, potentially minimizing systemic immunosuppression while retaining efficacy. 58
This study has several limitations. Molecular investigations were limited to a single time point (4 h), corresponding to the peak of behavioral recovery, which restricts interpretation of the temporal profile of curcumin’s effects. The cytokine array provided only relative quantification, and further validation using quantitative methods such as western blotting will be required. In addition, the relatively small sample sizes may limit the robustness of the statistical analyses; therefore, the present findings should be interpreted with appropriate caution.
Importantly, while the present study focused on peripheral mechanisms within the trigeminal ganglion, neuropathic pain is recognized to involve central sensitization processes in higher-order brain regions. In particular, the anterior cingulate cortex (ACC) and insular cortex (IC) have been implicated in the affective and cognitive dimensions of pain processing. Previous studies have demonstrated that synaptic plasticity within the ACC plays a critical role in chronic pain perception,59–61 while neuronal activation and circuit-level modulation in these cortical regions contribute to the integration of sensory and affective components of pain. 62 Furthermore, trigeminal nerve injury has been reported to induce cortical hyperexcitability in these regions. 60 In this context, it is plausible that the analgesic effects of curcumin observed in this study may also be associated with modulation of central neural activity. However, cortical neuronal activation was not directly assessed in the present study. Future studies evaluating markers of neuronal activation and plasticity, such as c-Fos or CREB in the ACC and IC, will be important to further elucidate the central mechanisms underlying the analgesic effects of curcumin.
Despite these limitations, the present findings identify curcumin as a promising therapeutic candidate for TN. By rapidly suppressing glial activation and selectively downregulating STAT3-related cytokines, curcumin disrupts early neuroimmune signaling cascades within the TG. Further studies should investigate its pharmacokinetics, long-term efficacy, repeated dosing, and broader molecular targets to establish its potential as a therapeutic agent for orofacial neuropathic pain.
Supplemental Material
sj-docx-1-mpx-10.1177_17448069261445281 – Supplemental material for Systemic curcumin administration mitigates neuropathic pain in a rat model of trigeminal neuralgia by suppressing TNF-α and pSTAT3 signaling
Supplemental material, sj-docx-1-mpx-10.1177_17448069261445281 for Systemic curcumin administration mitigates neuropathic pain in a rat model of trigeminal neuralgia by suppressing TNF-α and pSTAT3 signaling by Dara Sari Arini, Lutfi Putra Perdana, Jaime Fabillar, Mayu Miyagi, Daisuke Ikutame, Kazuyuki Koike, Shino Suma, Misaki Ichinomiya, Yoshizo Matsuka and Kosuke Kataoka in Molecular Pain
Footnotes
Acknowledgements
We would like to thank Professor Keiko Miyoshi from the Department of Frontiers in Dental Education and Science, Graduate School of Biomedical Science, Tokushima University for supporting our research.
Author contributions
DSA, MM, KK, and YM participated in the design and conceptualized the experiment. DSA, LPP, and JFJ carried out animal model and behavior test. DSA and LPP also performed qRT-PCR, immunofluorescence staining, and cytokine array. KK and YM conceived the project, ensured the funding and supervised the experiment. DSA analyzed the data and wrote the original manuscript. JFJ contributed to the language editing of the manuscript. LPP, JFJ, MM, DI, KK, SS, MI, YM, and KK edited the draft manuscript. All authors reviewed the final manuscript.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by grants from the Ministry of Education, Culture, Sports, Science, and Technology of Japan (No. 25K13127 for MM and No. 24K02659 for KK).
ORCID iDs
Data availability
The datasets generated during the current study are available from the corresponding author upon reasonable request.*
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.