
Abstract
Introduction:
It has well been documented that sex-related difference in the prevalence of migraine is widely accepted with more commonly seen in female patients. Although trigeminal ganglion (TG) neurons are the key players in the etiology of migraine, the underlying pathophysiology remains under debate so far.
Methods:
Myelinated Ah-type TG neurons were identified by the waveform characters of action potential (AP) conjugated with pharmacological validation using whole-cell patch techniques.
Results:
The results showed that AP duration and current derivative for repolarization were markedly increased by 3 µM Yoda1 along with the increased firing frequency of repetitive discharge that could be abolished by 3 µM GsMTx4. Although Yoda1 concentration-dependently increased the peaks of Ca2+ currents, the blocker for Ca2+ channel ω-Conotoxin did not alter the AP waveform characters and discharge profiles, whereas Yoda1-mediated changes in AP waveform trajectory and repetitive discharge could be completely reversed by 1.0 µM Iberiotoxin, a selective KCa1.1 blocker, suggesting that Piezo1-induced Ca2+ influx can activate KCa1.1 via presumably channel coupling. Additionally, Ah-type TG neurons functionally expressed Nav1.9/Nav1.8 in all tested neurons and their peaks were significantly increased by 3 µM Yoda1 and completely blocked by 3 µM GsMTx4.
Conclusion:
These datasets have demonstrated Piezo1-mediated neuroexcitation of female-specific subpopulation of myelinated Ah-type TG neurons due at least to the coupling between N-type Ca2+ channel and KCa1.1 and functional upregulation of Nav1.9, which uncovers an additional insights for higher incidence of migraine in woman.
Introduction
Increasing evidences from both bench and bedside have pointed out that pathophysiological and clinical insights are closely associated with gender-related incidence of migraine being commonly seen in the females.1–3 However, the developments and etiologies of migraine are extremely complicated and being debate. From neuroanatomical point of view, the evidence has been reported that there is a sub-population of myelinated Ah-type trigeminal ganglion (TG) neurons specifically identified from adult female rats,4,5 which is consistent well with the literature reports from other sensory neurons6,7 and sheds new light to clarify the etiological mechanism for migraine and answers the key question why is more common in female patients. The fact is that any reason causing the neuroexcitation of TG neurons would be considered as key to trigger the migraine. Based on this therapy, multiple ion channels fall into our scope of investigation and serve pleiotropic functions often being found in complexes, 8 for instance, Nav1.9/Nav1.8, 9 N-type Ca2+/KCa1.1, 10 and recently being discovered mechanosensitive Piezo1 with TRPM4 11 that aliens with the molecular evidence. 12 Therefore, the aim of current investigation is to understand how these ion channels collaborating together to influence the neuroexcitability of female-specific sub-population of myelinated Ah-type TG neurons isolated from adult female rats using whole-cell current- and voltage-clamp techniques. The major findings of our datasets demonstrated that firing frequency of Ah-types could be significantly increased by Piezo1 activation through not only the functional up-regulation of N-type Ca2+ channel currents and consequently KCa1.1 activation (indirectly), but also Nav1.9 (directly) as well Nav1.8 to facilitate the neurotransmission that may be responsible for or impact on the pathophysiology and etiology of migraine.
Materials and methods
Animals
Adult female Sprague–Dawley rats (weighting 200–220 g) were ordered from the SPF Experimental Animal Center at Dalian Medical University in a pathogen-free environment under controlled temperature and humidity with 12-h light cycle and free access of rodent food and tapped water. All experimental protocols were pre-approved by the Institutional Animal Care and Use Committee of the School of Medical Science at Dalian Medical University, which are in accordance with the recommendations of the Panel on Euthanasia of the American Veterinary Medical Association and the National Institutes of Health publication “Guide for the Care and Use of Laboratory Animals” (http://www.nap.edu/readingroom/books/labrats/).
Ethical considerations
The experiments on rats were approved by the Animal Use and Care Committee of Dalian Medical University (No. AEE24094, the Ethics Committee of Dalian Medical University). The experiments on rat conformed all methods were performed in accordance with the relevant guidelines and regulations.
Chemicals
Yoda1 (#HY-18723), GsMTx4 (#HY-P1410), Iberiotoxin (IbTX, #HY-P0190), ω-Conotoxin GIVA (ω-CTX, #HY-P0189), and Capsaicin (Cap, #HY-10448) were all purchased from MedChem Express (Monmouth Junction, NJ, USA). Tetrodotoxin (TTX, #1078) was ordered from Tocris (Ellisville, MO, USA). All chemicals were dissolved in a suitable solvent as stock, kept at −80°C, and diluted by recording solution prior to use. All other chemicals for neuron isolation and recording solutions were from standard routine commercial sources.
Neuron isolation
The rats were anesthetized with CO2 and subsequently decapitated and trigeminal ganglia (TG) were dissected from female adult rats. 13 For isolation, the whole ganglia were initially washed for 3 times with ice-cold Hank’s Balanced Salt Solution (Life Tech, Frederick, MD, USA) and digested using Papain (10 units/mL) for 20 min at 37°C, followed by a digestion cocktail made up of 1.0 mg/mL Collagenase type-II (Worthington Biochem. Corp. Lakewood, NJ, USA) and 2.5 mg/mL Dispase (Roche, USA) for additional 30 min at 37°C. The ganglion tissue was then washed twice in Dulbecco’s Modified Eagle Medium (HyClone, Logan, UT, USA), re-suspended in DMEM with 10% fetal bovine serum (HyClone), 10% heat-inactivated horse serum (Invitrogen, Grand Island, NY, USA), and 1% L-glutamine (Sigma, St Louis, MO, USA), and gently titrated with an aspiration pipette and cell suspension was placed onto poly-D-lysine (Sigma, St Louis, MO, USA) coated coverslips (10 × 10 mm diameter, Bellco Glass, Inc, Vineland, NJ, USA) for at least 4 h of incubation at 37°C routine cultured environment before patch recording.
Afferent fiber-type identification
In current-clamp recordings, Ah-type of TG neurons were identified based on the AP waveform characters including AP firing threshold (APFT), the maximal upstroke velocity (UVMAX) of depolarization, AP duration (APD50), and a repolarization hump. However, in voltage-clamp recording of Na+ currents, simply because the intracellular solution was not formulated for AP recording, so, it was impossible to get AP recorded for afferent fiber-type identification before Na+ current observation. In this case, Ah-type TG neurons was screened by its morphological characters initially 14 and confirmed at the end of recording using TTX conjugated with 100 nM Cap to eliminate A-types and C-types. For Ca2+ currents recordings, Ah-types could also be identified by their morphological characters and confirmed by changing the extracellular solution for Na+ at the end of experiment and checked with both blockers.
Electrophysiology
All electrophysiological and pharmacological studies were carried out at room temperature with isolated TG neurons that had been in culture for at least 4 but not more than 10 h. The whole cell current-clamp technique was carried out with a 700B patch-clamp amplifier (Axon Instruments, CA, USA). The pipettes were pulled from borosilicate glass (Sutter Instrument, Novato, CA) and heat polished (MF-830, Narishige, Tokyo, Japan) down to a resistance of 1–2 MΩ in normal recording solution (see below). After offset adjustment and formation of a Giga-ohm (GΩ) seal the pipette capacitance was compensated. On going to the whole-cell the neuronal capacitance (40–50 pF) and electrode access resistance were also compensated (60%–80%). Single APs were elicited with a 500-s current pulse, and repetitive AP discharge was evoked with a current step, both with magnitudes 1.5 that required for threshold discharge unless otherwise specified. Raw data were low-pass filtered to 10 kHz and digitized at 50 kHz. All experiments were conducted and visualized under Axioskop routine microscope (Zeiss, Germany) at room temperature (22 ± 1°C). Experimental protocols, data collection, and preliminary analysis were carried out with pCLAMP 10 and the Digidata 1440A (Axon Instruments, CA, USA) operating on a PC platform.
Recording solutions
The bath/extracellular recording solution contains (in mM): (1) for AP recordings: 137.0 NaCl, 5.4 KCl, 1.0 MgCl2, 10.0 glucose, and 10.0 HEPES; (2) for Ca2+ current recordings: 140.0 TEA-Cl, 5.0 4-aminopyridine, 15.0 HEPES, 20.0 glucose, and 2.0 CaCl2; (3) for Na+ current recordings: 50.0 NaCl, 10.0 MgCl2, 10.0 glucose, and 25.0 HEPES. The pipette/intracellular solution contains (in mM): (1) for AP recordings: 6.0 NaCl, 50.0 KCl, 50.0 K2SO4, 5.0 MgCl2, and 10 HEPES; (2) for Ca2+ current recordings: 124.0 CsCl, 5.0 MgCl2, 10.0 glucose, and 10.0 HEPES; (3) for Na+ current recordings: 7.0 NaF, 195.2 NMDG, 165.7 TEA-Cl, 2.0 MgCl2, and 10.0 HEPES. The pH for extracellular or intracellular solutions were adjusted to 7.30 and 7.20, respectively, using 1 N NaOH or I N KOH. The osmolarity of extracellular and intracellular solutions were adjusted to 310 and 290, respectively, using D-mannitol (Sigma). For providing a physiological intracellular environment, the final [Ca2+]i for all intracellular solutions were adjusted to 100 nM by adding 4.0 mM Bapta-K (Sigma), 0.25 mM CaCl2, and 2.0 mM each of Na-ATP (Sigma) and Na-GTP (Sigma) to the pipette solution just before the experiments.
Data acquisition and statistical analysis
For voltage-clamp recordings, the current values were quantified by directly measured peaks due to a similar whole-cell capacitance (WCC/pF) of all tested Ah-type TG neurons in our current observation. All electrophysiological recordings were analyzed with Clampfit (v10, Axon Instruments, CA, USA), and pooled statistics were calculated with Excel or Origin (Microsoft, CA, USA). Average data are expressed as means ± SD, with significance assessed by paired or unpaired Student’s t-test, or one-way ANOVA with post-hoc Tukey test where appropriate. A p value of <0.05 was considered significantly different.
Results
Piezo1 channel activation mediates a neuroexcitability in identified Ah-type trigeminal neurons
Increase in the neuroexcitation by Piezo1 channel activation has been reported in cardiomyocytes, 15 smooth muscle cells, 16 bone cells 17 and also sensory neurons including nodose ganglion (NG),6,18 dorsal root ganglion (DRG), 19 and trigeminal ganglion neurons (TG). 20 Here, this investigation was focused on the mechanism insight into the gender-related difference of migraine, so, adult female rats were used for isolated neurons preparation and a female-specific subpopulation of myelinated Ah-type TRG was identified based on its electrophysiological characters,4,5 such as the action potential (AP) waveform/current derivative (SFigure 1A) and phase plot (SFigure 1B). To test if Piezo1 activation affects the neuroexcitability, the step current depolarization was used to elicit repetitive discharge in identified Ah-type TG before and after 3 µM Yoda1. The results showed that, under identical current steps, the firing frequency was markedly increased from the control level, and this enhanced excitability could be completely reversed by 3 µM GsMTx4 (Figure 1) on top of Yoda1.

Piezo1 activation-mediated changes in action potential (AP) waveform trajectory in identified Ah-type trigeminal ganglion (TG) neurons isolated from adult female rats. Single AP was elicited by a brief current pulse and Ah-type neuron was identified by its waveform characters such as AP firing threshold below −40 mV, maximal up-stroke velocity (UVMAX) over 250 mV/ms, brief APD50 within 2 ms, and possible hump over the time course of repolarization. (a–b) Representative APs and current derivatives before and after 3 µM Yoda1; (c–d) Averaged data for APD50 and current derivative regarding the repolarization (negative/downward portion). Averaged data were presented as mean ± SD, n = 10 complete recordings from at least four rats/preparations. Scale bar in (a) also applied for (b).
Changes in waveform trajectory of Ah-type TN neurons
AP is the basic unit of neuronal function and sensory inputs are encoded into the frequency train, so, any detectable changes in the trajectory of AP is always meaningful to indicate the ion channel current alternation behind the membrane voltage. To this regard, AP and its current derivative obtained before and after treatment were superimposed and this analysis showed that ADP50 was shortened (Figure 2(a) and (c)) and total outward current of current derivative plot was increased (Figure 2(b) and (d)) by Yoda1 treatment, highly suggesting that Ca2+-activated K+ channel (KCa1.1 or BK-type) 21 is activated through the coupling mechanism 22 with N-type Ca2+ channel.

Piezo1 activation-mediated changes in the frequency of repetitive discharge profiles in identified Ah-type trigeminal ganglion (TG) neurons isolated from adult female rats. The repetitive discharge of identified Ah-type TG neurons was elicited by a fixed step current depolarization before and after treatments. (a) Representative recording of repetitive discharges under 100, 200, and 300 pA current intensity before Yoda1; (b–c) Representative recordings in the presence of 3 µM Yoda1 and 3 µM GsMTx4 on top of Yoda1; (d) Summarized frequency changes of repetitive discharge with or without treatments. Averaged data were presented as mean ± SD, n = 10 complete recordings from at least four rats/preparations. Scale bar in (a) applied for all.
Piezo1 activation-mediated peak current of N-type Ca2+ increased
It is well known that Piezo1 channel is permeable to cation ions without selectivity to Na+ and Ca2+, therefore, the involvement of N-type Ca2+ channel in Yoda1-induced AP trajectory change is worthy to be tested at first under voltage-clamp configuration with holding potential of −100 mV (SFigure 2) and the results showed that the peak of Ca2+ currents measured at 0 mV were obviously increased by Yoda1 in a concentration (0.1–3 µM)-dependent manner (Figure 3) without any detectable changes in the voltage-dependent profiles (data not shown) and the effect of Yoda1 on Ca2+ currents was abolished by 3 µM GsMTx4. Owing to the similar size of tested neurons, the current density was not obtained by divided the peak with the whole-cell capacitance. Recent literature has reports that Expression of Piezo1 is mainly detected in the myelinated afferent fiber/neurons of TG, 12 for double check, 100 nM Capsaicin was added in the recording chamber at the end of recording to make sure that tested neurons is not Cap-positive.

Poezi1 activation induced functional upregulation of Na-type Ca2+ channel currents in identified Ah-type TG neurons in the presence. Upper panel: Effects of Piezo1 channel activation and inhibition on the peaks of Ca2+ currents before and after Yoda1 or GsMTx4 treatments; Lower panel: Summarized results. Averaged data were expressed as mean ± SD, n = 3–11 complete recordings from at least three rats/preparations upon the tested concentration.
Modulated effect of KCa1.1 blocker on Yoda1-mediated AP trajectory change
It is an important fact that Ca2+ currents are indeed enhanced by Piezo1 activation, however, unlike the cardiomyocyte and skeletal muscle cells, Ca2+ currents are not directly modulate the AP waveform trajectory (SFigure 3) obtained before and after 1 µM Conotoxin GVIA (ω-CTX), but it might be indirectly via KCa1.1 through the co-localization or coupling mechanism with N-type Ca2+ channels. As our expected, Yoda1-mediated changes in AP trajectory and firing frequency (Figure 4(a) and (b)) were reversed in the presence of 1 µM Iberiotoxin (IbTX) in identified Ah-type TG neurons (Table 1). These datasets confirm that enhanced Ca2+ influx via N-type Ca2+ channels does not directly participate the modulation on AP trajectory and firing frequency but can activate KCa1.1 and indirectly upgrades neuroexcitation through both alternation of AP waveform trajectory and profiles of repetitive discharge.

Potential impact of KCa1.1 involvement in Poez1 activation-mediated changes in AP trajectory and frequency of repetitive discharge. (a–b) Single AP and current derivatives were elicited by brief current pulse and step current depolarization were collected from identified Ah-type TG neurons before after Yoda1 and Iberiotoxin (IbTX) on top of Yoda1, scale bar in (b) also applied for (a); (c) Repetitive discharge profiles before and after Yoda1 and IbTX on top of Yoda1. Summarized results were listed in the Table 1.
Changes in parameters of action potential (AP) waveform trajectory and firing frequency of repetitive discharge before after treatment with Yoda1 and Iberiotoxin (IbTX) on top of Yoda1.

APD50: AP duration at 50% height of AP/ms; APFF: AP firing frequency/Hz; APFT: AP firing threshold/mV; DVMAX: the maximal down-stroke velocity/mV/ms; PeakAP: the peak of AP/mV; RMP: resting membrane paternal/mV; UVMAX: the maximal up-stroke velocity/mV/ms.
Averaged data were presented as mean ± SD, n = 4 complete recordings from at least three rats/preparations.
p < 0.05 and **p < 0.01 versus control; #p < 0.05 and ##p < 0.01 versus Yoda1.
Functional upregulated current peak of Nav1.9 in the presence of Yoda1
Increasing evidences have shown that Nav1.9 is a key player in retaining the higher neuroexcitability in the sensory neurons and closely associated with pathophysiological pain processing7,23,24 and Nav1.9 is also functionally expressed in a female-specific subpopulation of Ah-types TG neurons.4,5 Based on these facts, we have strong reason to speculate that Nav1.9 might be involved in Piezo1 induced neuroexcitation. By using whole-cell voltage-clamp protocol (SFigure 4), persistent TTX-resistant Na+ currents (Nav1.9) were recorded at holding potential between −60 and −25 mV in identified Ah-type TG neurons (Figure 5, n = 11) in the presence of 100 nM TTX, rather than those tested C-types with functional expression of Nav1.8 only (SFigure 5). Strikingly, the peak currents were dramatically increased by 3 µM Yoda1 and this effects of Yoda1 on Nav1.9 could be completely reversed back to the control level (Figure 5(a) and (b)), meanwhile, TTX-resistant Na+ currents (Nav1.8) obtained in these Ah-type TG neurons were also upregulated simultaneously (Figure 6), suggesting that Piezo1 activation induces neuroexcitation co-interplayed by both KCa1.1/N-type Ca2+ and Nav1.8/Nav1.9 in these Ah-type TG neurons.

Alternation of peaks of Nav1.9 current before and after Piezo1 activation or inhibition in identified Ah-type TG neurons. Nav1.9 was recorded in the presence of 100 nM Tetrodotoxin (TTX) to block Nav1.7’s potential contamination on the current peaks. (a) Representative peak currents before and after Piezo1 activation or inhibition; (b) Summarized results of peak current change. Averaged data were expressed as mean ± SD, n = 10 complete recordings from at least six rats/preparations. Both paired Student’s t-test and one-way ANOVA were applied for the statistical analysis.
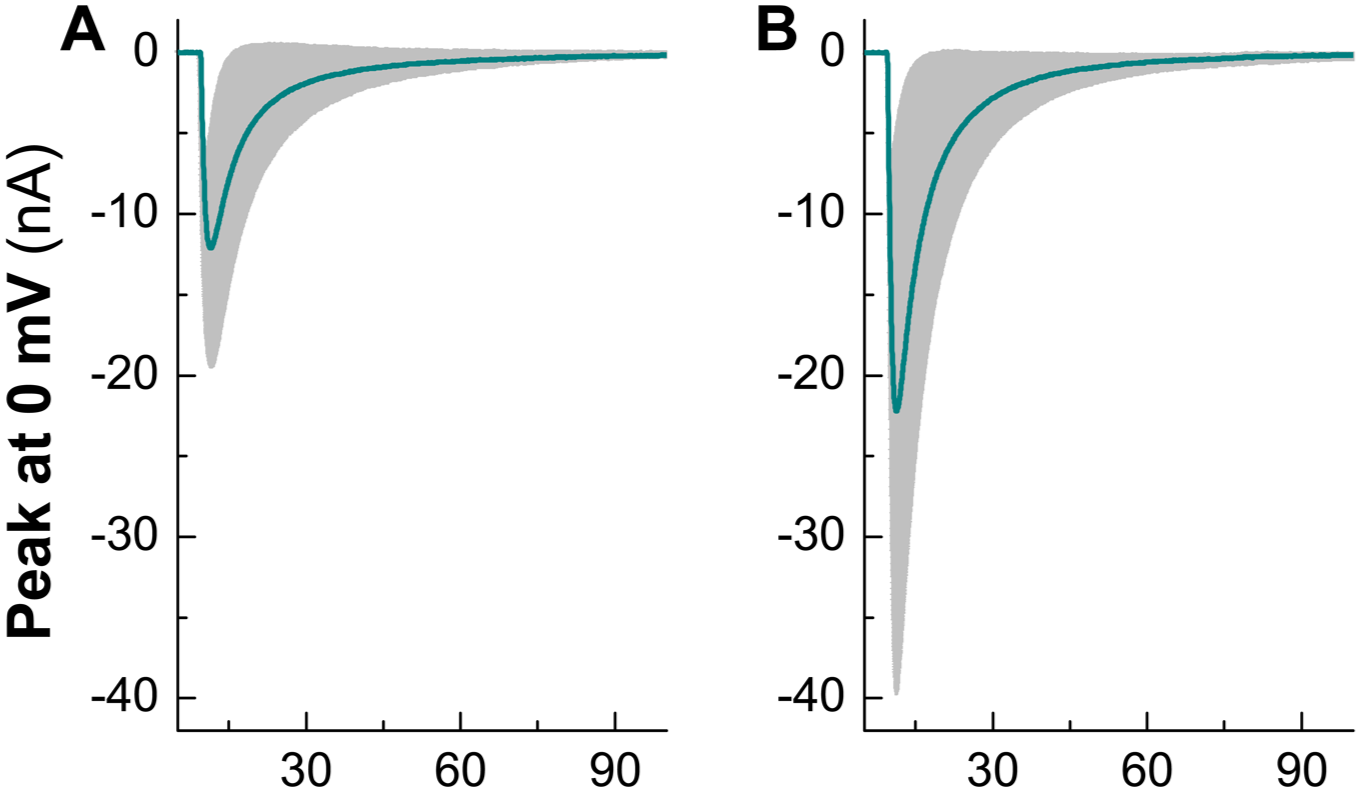
The peak currents changes of Nav1.8 in the presence of Yoda1. (a and b) Representative recordings of Nav1.8 before and after Yoda1. n = 5 complete recordings from at least three rats/preparations. 100 nM Tetrodotoxin (TTX) was included in the recording solution to eliminated the potential contamination of Nav1.7.
Discussion
Migraine is a chronic neurological disorder characterized by recurrent moderate-to-severe headaches with approximately nearly 20% of the population is affected by migraines at some point in life globally and more commonly seen in female patients than that in age-matched males in adulthood.1,2 The pathophysiology of migraines is believed to be attributable to involve complex interactions between genetic, neurological, and environmental factors. 25 Circulating hormone levels may also play a crucial role in the etiology of the disorder.26–28 The precise mechanisms of migraine are still under discussion and debate unknown. Nonetheless, it is believed to be a neurovascular disorder. 29 Increasing evidences imply that the primary theory of migraine is related to increased excitability and abnormal control of pain neurons in the trigeminal nucleus of the brainstem.30,31 More excitingly, a female-specific sub-population of myelinated Ah-type neurons has recently been identified from sensory nerve system including nodose ganglia6,7 and TG4,5 of adult female rats, and their neuroexcitability are controlled by estrogen and closely associated with functional expression of multiple ionic channels such as KCa1.1, 10 Nav1.9,4,7 and Piezo1. 6 Although the insights into the mechanisms underlying the pathophysiology of migraine remain not fully understood, the evidences aforementioned lead us into the current pass through which collaborative/synergistic role of multiple ion channels in neuroexcitation of Ah-type TG neurons are explored. Piezo1 as mechanosensitive channel has been demonstrated that its activation could initiate a host of intracellular signaling cascades through conversion Ca2+ mobilization to biomechanical signals32–34 and other channels modulation11,35,36 in many tissue and organ systems, so, the current investigation has explored whether or not Piezo1 activation triggers the functional up-regulation of AP and repetitive discharge profiles in identified Ah-type TG neurons through KCa1.1 and Nav1.9 modulation using electrophysiological approach. Apparently, Piezo1 activation-mediated significant increase in the neuroexcitation of Ah-type TG neurons is fully supported by the notion that the repolarization process is accelerated by narrowing the APD50 and increasing DVMAX, and this is attributed to functional up-regulation of KCa1.1 followed by the enhancement of Ca2+ currents through Piezo1 channel due presumably to the coupling/colocalization between Piezo1 and KCa1.1, which is a common mechanisms being recognized within other channels.11,22,37 Although the current observation do not examine if these channels are co-localized using immunostaining or molecular approaches, their existence in the TG neurons has been detected.12,38 Interestingly, the fact of expression of Piezo1 detected only in the myelinated TG neurons highly supports our observation because of its myelinated feature of Ah-type TG.
Nav1.9 as a persistent TTX-resistant Na+ channels plays a critical role in controlling the nociceptive process in sensory nerve system 24 and dominantly expresses in Ah-type TG neurons4,5 with higher neuroexcitability. Even though there is no existing evidence showing the relation of its activity being modulated by Piezo1 activation, it is worthy of notice if Ca2+ influx through Piezo1 is an important trigger for Nav1.9. Strikingly, the peak currents of Nav1.9 can be elevated dramatically by Yoda1 and reversed by GsMTx4, suggesting the functional connection between these channels that could not be explained and warranted for future investigation.
One of technical limitation is that, in the case of Ca2+ current recording under voltage-clamp mode, identification of afferent-fiber type of isolated TG neurons could only be confirmed based upon their morphological characters conjugated with or without inward current in the presence of 100 nM Cap tested at the end of recording.
For outline the functional role of all types of TG neurons, an identical observation using myelinated A- and unmyelinated C-types isolated from adult male and age-matched female rats were also conducted under the similar experimental condition, which will be included in our ongoing article.
Taken all these data together, we could conclude that the neuroexcitation of female-specific distributed myelinated Ah-type TG neurons could be significantly increased by Piezo1 activation through not only the functional up-regulation of KCa1.1 activation due most likely to the coupling, but also Nav1.9 facilitation that may be responsible for or impact on the pathophysiology and etiology of migraine. Future research should focus on validating channel co-localization, elucidating estrogen-dependent modulation, and exploring downstream signaling pathways to clarify the mechanistic link between mechanosensation and migraine. These findings highlight the therapeutic potential of targeting the Piezo1-KCa1.1-Nav1.9 axis for developing gender-specific treatments, such as novel channel modulators or non-invasive neuromodulation strategies, ultimately advancing precision medicine for migraine and related chronic pain disorders.
Supplemental Material
sj-docx-1-mpx-10.1177_17448069251410754 – Supplemental material for Piezo1-mediated neuroexcitation via collaboration with KCa1.1 and Nav1.9 currents in myelinated Ah-type of trigeminal ganglion neurons in rats: Mechanistic insights with sex-specific effects
Supplemental material, sj-docx-1-mpx-10.1177_17448069251410754 for Piezo1-mediated neuroexcitation via collaboration with KCa1.1 and Nav1.9 currents in myelinated Ah-type of trigeminal ganglion neurons in rats: Mechanistic insights with sex-specific effects by Yang Liu, Yao Song, Ximeng Ren, Meng Li, Shuangshuang Liu, Zixuan Li, Dapeng Ding and Xiaolong Lu in Molecular Pain
Footnotes
Acknowledgements
We acknowledge the Dalian Medical University and the First Affiliated Hospital of Dalian Medical University for their financial support.
Author contributions
Conceptualization: YS; Methodology: ML; Data collection: XMR, SSL; Statistical analyses: ZXL; Writing – original draft preparation: YL; Writing – review and editing: DPD, XLL; All authors have approved the final version of the manuscript and agree to be accountable for all aspects of the work.
Data availability statement
Data available on request from the authors.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by grants from Youth Talent Cultivation Fund Project of Dalian Medical University to Xiaolong Lu; the Natural Science Foundation of Liaoning Province (No.2023-MS-262 to Yang Liu), the Foundation of Liaoning Province Education Administration (No. LJKMZ20221276 for Dapeng Ding) and Key Program of the Medical-Engineering Joint Innovation Fund of the First Affiliated Hospital of Dalian Medical University and Dalian Institute of Chemical Physics, Chinese Academy of Sciences (DMU-1&DICP UN202304 to Dapeng Ding).
ARRIVE guidelines statement
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.