
Abstract
Keywords
Introduction
The chronic complications of diabetes attack multiple organs, causing disability and high mortality. 1 When the upper gastrointestinal tract is involved, patients often have a variety of clinical symptoms, including upper abdominal pain or discomfort, nausea and vomiting, acid reflux and postprandial fullness. Studies reported that 40–80% of diabetic patients have varying degrees of upper gastrointestinal symptoms, which seriously affects the quality of life.2,3 At present, the pathogenesis of the disease has not yet been elucidated. Several lines of evidence suggest that the discomfort and pain of gastroparesis is associated with altered function of visceral afferent pathways. We have recently reported that promoted interaction of nuclear factor-κB (NF-κB) with gene of endogenous hydrogen sulfide producing enzyme cystathionine β-synthetase (CBS) played an important role in gastric hypersensitivity. 4 However, the precise mechanism of NF-κB activation in gastric-specific primary sensory neurons remains unknown.
Toll-like receptors (TLRs) play important roles in natural and adaptive immunity, inflammatory response, and autoimmunity. 5 Recent studies suggest that continuous activation or dysregulation of TLR signaling may contribute to chronic pain and is expected to be an emerging therapeutic target for persistent pain.6,7 TLR4 can activate NF-κB through MyD88-dependent and -independent signaling pathways, and regulate the expressions of many downstream inflammatory factors. 7 We speculate that TLR4 may also be involved in diabetic gastric hypersensitivity.
MicroRNA (miRNA) is a kind of non-coding single-stranded small RNA with a length of about 20–24 bp. It plays a negative or positive role in regulating gene expression by complementary binding with the 3′-UTR of the target mRNA. Growing studies have shown that miRNAs were involved in chronic pain and visceral pain.8–11 TLR4 has many endogenous ligands, which can affect TLR4 expression and function. 12 We wonder whether miRNAs can regulate TLR4 expression.
Therefore, we hypothesized that the loss of miRNA could regulate the expression of TLR4, which activating NF-κB, and induce enhanced excitability of DRG neurons, thus contributing to diabetic gastric hypersensitivity. To verify the hypothesis, we chose a suitable and stable animal model of diabetes. Our findings suggest that miR181a/TLR4/NF-κB signaling might be a novel mechanism underlying the diabetic gastric hypersensitivity. This and future studies might shed light on the clinical mechanisms of gastric pain in patients with diabetes.
Materials and methods
Generation of streptozotocin-induced diabetes
All animal experiments were approved by the Institutional Animal Care and Use Committee at the Soochow University and were in accordance with the guidelines of the International Association for the Study of Pain. Adult female Sprague-Dawley (SD) rats (weighing 180–200 g) were housed 4 per cage in a temperature-controlled (25 ± 1°C) and 12 h/12 h light/dark cycle room. All experimental rats were allowed access to tap water and standard laboratory chow ad libitum.
Diabetes mellitus (DM) model was induced by a single intraperitoneal injection of streptozotocin (STZ, 65 mg/kg, Sigma Chemicals, St. Louis, MO, USA), which was freshly dissolved in citrate buffer (10 mmol/L, Na citrate, pH = 4.3–4.4), as described previously.4,13 The control (CON) rats only received an equivalent volume of citrate buffer. One week later, fasting blood glucose concentration obtained from the tail vein was measured by glucometer (Johnson & Johnson, New Brunswick, NJ, USA). Only rats with blood glucose concentrations higher than 15.0 mmol/L (270 mg/dl) were further used in the study and data analysis.
Implantation of gastric distension balloon and electrodes
All behavioral studies were performed under blind conditions and a total of 89 SD rats were used for electromyographic (EMG) recording in the study. All surgical procedures were performed under sterile conditions. The process for the implantation of the gastric distension (GD) balloon was as described previously 14 with some modifications. In brief, a 2.5-cm-long balloon made from latex condoms was attached to a catheter (PE-90). An incision was made in left lateral abdomen to expose the stomach, and the balloon was placed properly in the stomach through a small hole made at the tip of the fundus. Then the hole was securely tied to avoid leakage of gastric fluid. The EMG electrode leads and the polyethylene tubing, which is used for air inflation of the gastric balloon, were exteriorized at the back of the neck. After the surgery, the incisions of abdominal cavity and skin were sutured.
To record EMG activities, 40-gauge teflon-insulated stainless steel wires (Cooner Wire) were implanted in the acromiotrapezius (a superficial neck muscle) by aseptic techniques, and the incision was closed with a 4-0 silk suture. The electrode leads were externalized at the back of the head for easy access during experiments.
EMG recording
One week after abdominal surgery, rats were intraperitoneally injected with STZ (65 mg/kg, n = 71) or the same volume of citrate buffer (n = 18). After 2 weeks, six rats from each group were detected once a week for EMG responses to GD in graded 60–120 mmHg distention pressure. Response to GD was defined as the increase in EMG activity relative to baseline during the 20s GD period. Data were recorded as the area under curve (AUC) of the integrated EMG after baseline subtraction and standardized to the AUC rate. A number of the DM rats (n = 41) and CON rats (n = 12) were used to study the effect of the TLR4-selective inhibitor CLI-095 and LV-p65 siRNA (Lentiviruses, LV) on behavior. 4 weeks after STZ injection, 41 DM rats were divided into 6 groups and received an intrathecal injection of normal saline (NS, i.t., n = 6), CLI-095 15 μg/kg (i.t., n = 6), CLI-095 50 μg/kg (i.t., n = 6), or CLI-095 150 μg/kg (i.t., n = 6), LV-NC siRNA (1 × 109 TU/ml, i.t., n = 8), and LV-p65 siRNA (1 × 109 TU/ml, i.t., n = 9). In addition, age- and sex-matched CON rats received CLI-095 injection (50 μg/kg, i.t., n = 6) and LV-p65 siRNA injection (1 × 109TU/ml, i.t., n = 6). EMGs were recorded 0.5, 2, 4, 8, 12, and 24 h after CLI-095 treatment. 15 The rest DM rats (n = 24) were used to study the effect of miR-181a agomir on behavior. 3 weeks after STZ injection, 24 rats were divided into four groups and received intrathecal injections of NC agomir (single intrathecal injection, n = 6), miR-181a agomir (single intrathecal injection, n = 6), NC agomir (intrathecal injection once a day for seven consecutive days, n = 6), miR-181a agomir (intrathecal injection once a day for seven consecutive days, n = 6).
Cell retrograde labeling
As described previously,4,13 the origin of the primary afferent innervating the stomach was labeled by retrograde tracing with DiI (Invitrogen, New York, NY, USA). Experiments were performed on 22 female SD rats. Food was withheld for 24 h before DiI injection surgery. The animals were anesthetized with a combination of Ketamine (100 mg/kg) and Xylazine (25 mg/kg). After the stomach was exposed completely by midline abdominal incision, DiI (25 mg in 0.5 mL methanol) was injected into the ventral (ten sites) and dorsal (ten sites) walls of the corpus, pyloric antrum, or fundus with a Hamilton microsyringe. Each site was injected with 1 μl DiI. After 1 week, 22 rats were injected intraperitoneally with STZ. Among them, eight DM rats were divided into two groups (n = 4 for each group), receiving LV-NC siRNA or LV-p65 siRNA intrathecal injection, respectively. The other eight rats were injected intrathecally with NS or CLI-095 at 50 μg/kg once a day for seven consecutive days at 3 weeks after STZ injection (n = 4 for each group). The rest 6 DM rats were divided into two groups (n = 3 for each group), and injected intrathecally with NC agomir or miR-181a agomir once a day for seven consecutive days at 3 weeks after STZ injection, respectively. After the above treatments, the bilateral T7-T10 DRGs were dissected to study the excitability of gastric-specific neurons.
Dissociation of DRG neurons and patch-clamp recording
Neuronal excitability was detected by whole-cell patch-clamp recording techniques as described previously.4,16 DiI-labeled neurons were identified by fluorescence microscope (Olympus IX71, Japan). For the patch-clamp recordings, small and medium sized DRG neurons were chosen in our study, because they were considered to be responsible for pain sensation.17,18 Resting potential (RPs) and action potentials (APs) were recorded. The voltage was clamped at −60 mV. Whole-cell current and voltage were recorded with a HEKA EPC10 patch-clamp amplifier. The data were acquired and stored on a computer and analyzed by Fit Master from HEKA. The above experiments were performed at room temperature (22°C).
Real-time PCR
The total RNA was extracted by applying Trizol reagent (ambion) according to the manufacturer’s protocol. Quantitative PCR analysis was performed to detect the expression of microRNA in T7-T10 DRGs from CON and DM rats. The sequences of primers are shown in Supplementary Table S1. U6 small nuclear RNA was used as endogenous control to normalize differences for microRNA detection, and analyzed with the 2−ΔΔCt method.
Western blotting analysis
TLR4, TRAF6, and p65 expressions in T7-T10 DRGs from CON and DM rats were detected using Western blotting analysis. The primary antibodies used are listed below: anti-TLR4 (1:1000; Abcam, USA), anti-TRAF6 (1:1000; Abcam, USA), anti-p65 (1:200; Santa Cruz, USA), and anti-GAPDH (1:1000; Goodhere, China). Immunoreactive proteins were detected by enhanced chemiluminescence (ECL kit; GE Healthcare Pharmacia Biotech). The densities of protein bands were analyzed with NIH Image software.
Lentivirus vector generation and intrathecal injection
As described previously, 19 the siRNA targeting the cDNA sequence of rat p65 (GenBank Accession #NM_199,267) was used, with the sequences as follows: 5′-GCAGUUCGAUGCUGAUGAAUU-3′. An additional scrambled sequence was also designed as a negative control (NC). The cDNAs corresponding to the p65 siRNA and NC were subcloned into the vector LV pFU-GW-RNAi-GFP (Shanghai Gene Chem Co. Ltd.). The resulting recombinant LVs were designated as LV-p65 siRNA or LV-NC. The titer of LV-p65 siRNA and LV-NC was determined by a hole-by dilution titer assay and the final titer was 1 × 109TU/mL. A pilot study with LV-GFP was performed firstly to determine the level of infection efficiency of virus constructs. The lentiviruses were then injected into rats intrathecally. Behavioral and electrophysiological experiments were performed 4 weeks later.
Dual-luciferase reporter assay
The fragment of the rat TLR4 3′-UTR (481 nt) containing potential miR-181a and miR- 7a target sites (nucleotides 370–376 and 455–460 of the TLR4 3′-UTR, UGAAUGU and UCUUCC) and a fragment containing a mutated TLR4 3′-UTR (481 nt) without miR-181a and miR-7a target sites (AUCAUACA and GAGAAAU) within the seed region were inserted into the luciferase reporter plasmid (GP-miRGLO). PC12 cells were seeded into 24-well plates and co-transfected with 200 ng of the luciferase reporter plasmid and 20 nM of the miR-181a mimic (AACAUUCAACGCUGUCGGUGAGU) or miR-7a (UGGAAGACUAGUGAUUUUGUUGU) using Lipofectamine 2000 (Invitrogen). After 24 h, the firefly and Renilla luciferase activity were measured with the dual-luciferase reporter assay system (Promega) according to the manufacturer’s instructions. Firefly luciferase activity was normalized to Renilla luciferase activity for each transfected well. The experiments were performed in triplicate.
Immunofluorescence study
The rats were anesthetized deeply and perfused transcardially with 200 mL PBS (Thermo, United States), followed by 400 mL ice-cold 4% paraformaldehyde (Sinopharm Chemical Reagent Co. Ltd., China) in PBS 1 week after the DiI injection. Sections were simultaneously incubated with two antibodies: TLR4 (1:200; Santa Cruz) and NF-κB (1:200; Santa Cruz), and then incubated with Alexa Fluor 355 and 488 (1:500, Life Technologies Inc., Gaithersburg, MD, United States). After being covered with cover slips, images were captured and analyzed using Metaview software.
Fluorescence in situ hybridization
IsHybridIn Hybridization (ISH) Kit (Cat. #K2191050-5, Biochain) was used to detect the distribution of miRNA-181a and TLR4. Experiments were conducted using ISH Kit based on the manufacturer’s manual with minor modifications. Briefly, T7-T10 DRGs were removed, post fixed and dehydrated. DRGs were cut into 15 μm sections. Sections were incubated in 3% pepsin diluted in fresh citrate buffer at 37°C for 30 min and then prehybridized in prehybridization solution for 2 h at 37°C and then hybridized overnight at 37°C with a digoxigenin-labeled miRCURY LNA miRNA detection probe for miR-181a (Univ-Bio) at 3 ng/μL in hybridization Samples were then washed with a graded series of SSC buffer, blocked with 3% BSA, and incubated with alkaline phosphatase-conjugated sheep 6 anti-DIG Fab fragments. The probe for miR-181a was 5′-AACAUUCAACGCUGUCGGUGAGU-3′. The sections were mounted and images were captured and analyzed using Metaview software.
Targeted miRNA prediction
miRNA target prediction and functional annotations were examined by several online databases.20,21 Using the overlapping results obtained from the miRanda, TargetScan, and miRTarBase databases, assessed using a Venn diagram; Predicted miRNAs that target TLR4 were identified.
Statistical analysis
No DRG neuron with an RP more depolarized than −40 mV was included in the data analysis. All data are expressed as means ± sem. Statistical analysis was conducted using GraphPad Prism. Normality was first checked for all data before analysis. Significance was determined using a two-sample t-test, two-way ANOVA followed by Sidak’s post hoc test and Dunn’s post hoc test, as appropriate. Results were considered statistically significant when the p value was less than 0.05.
Results
STZ injection induced gastric hypersensitivity
STZ has been widely used to induce type 1 diabetes in rodents for the study of diabetic gastrointestinal complications4,22 and diabetic neuropathic pain.13,23 Consistent with our previous studies,
24
blood glucose elevated 1 week after a single injection of STZ and maintained for at least another 4 weeks. To determine whether STZ induced gastric hypersensitivity of diabetic rats, we recorded EMGs of the acromiotrapezius muscle responding to gastric distention at 2 and 4 weeks after STZ injection or CON. The results showed that AUC rate of DM rats increased significantly at distention pressures of 60, 80, 100, and 120 mmHg at 2 and 4 weeks after STZ injection (Figure 1(a)–(d)), particularly at 4 weeks. In agreement with our previous reports,
4
diabetic rats were more sensitive to graded gastric balloon distension as evident by the AUC. STZ injection induced gastric hypersensitivity and upregulation of TLR4 at 2 weeks and 4 weeks. (a) Representative EMG recordings from CON and diabetic rats responding to graded GD at 60, 80, 100 and 120 mmHg 2 weeks after vehicle (CON) or STZ injection (DM). (b) Representative EMG recordings from CON and DM rats responding to graded GD at 60–120 mmHg 4 weeks after vehicle or STZ injection. (c) AUC rate of EMG recordings from the DM group was significantly higher than CON at distention pressures of 60–120 mmHg at 2 weeks (CON, n = 6; DM, n = 10, *p < 0.05, **p < 0.01, ***p < 0.001, compared with CON, two-way ANOVA followed by Sidak’s post hoc test). (d) AUC rate of EMG recordings from the DM group was significantly higher than CON at distention pressures of 60–120 mmHg at 4 weeks (CON, n = 6; DM, n = 10, **p < 0.01, ***p < 0.001, compared with CON, two-way ANOVA followed by Sidak’s post hoc test). (e) TLR4 protein expression was greatly enhanced in T7-T10 DRG of diabetic rats compared with CON rats 2 weeks after injection (CON, n = 3; DM, n = 4, *p < 0.05, compared with CON, two-sample t-test). (f) TLR4 protein expression was greatly enhanced in diabetic rats compared with CON rats 4 weeks after injection (CON, n = 5; DM, n = 6, **p < 0.01, compared with CON, two-sample t-test).
TLR4 inhibitor CLI-095 treatment attenuated STZ-induced gastric hypersensitivity
Our previous studies reported that TLR4 was involved in chronic visceral hypersensitivity.
25
To determine whether TLR4 is involved in the development of gastric hypersensitivity of DM rats, the western blotting was performed to detect the expression of TLR4 in T7-T10 DRGs. TLR4 expression was upregulated at 2 and 4 weeks after STZ-induced diabetes (Figure 1(e) and (f)). Then we observed the effect of CLI-095, a selective inhibitor of TLR4, on EMG amplitude in response to GD pressure at 4 weeks after STZ treatment, a total of 24 diabetic rats with intragastric balloons were administrated CLI-095 in different doses (15, 50, and 150 μg/kg, i.t.). EMG recordings were recorded from diabetic rats in response to graded GD 30 min after CLI-095 treatment. NS and CLI-095 at the lowest dose (15 μg/kg) had no significant effect on EMG amplitude. However, CLI-095 at 50 and 150 μg/kg resulted in a dramatic reduction in EMG responses at 60–120 mmHg GD pressures (Figure 2(a) and (b)). More importantly, the TLR4 inhibitor CLI-095 significantly decreased STZ-induced gastric hypersensitivity in a dose-dependent manner. The optimized dose of CLI-095 that produced the maximal effect was 50 μg/kg body weight in the study. We used this dose to determine the time course of CLI-095 effects. As shown in Figure 2(c) and (d), the analgesic effect of CLI-095 lasted for approximately 4 h, and maximal inhibition occurred at 0.5 h. In addition, CLI-095 at 50 μg/kg did not produce a significant effect on AUC of EMG in age- and sex- matched CON rats (data were not shown). These data indicate that TLR4 is necessary for the STZ-induced gastric hypersensitivity. TLR4 inhibitor CLI-095 treatment attenuated STZ-induced gastric hypersensitivity. (a) Representative EMG recordings from diabetic rats treated with NS (normal saline) and CLI-095 (15, 50 and 150 μg/kg) responding to graded GD at 60, 80, 100 and 120 mmHg 4 weeks after STZ injection. (b) AUC rate of EMG recordings from the 50 and 150 μg/kg CLI-095 group resulted in a dramatic reduction in EMG responses at 60–120 mmHg GD pressures compared with NS and 15 μg/kg CLI-095 group (n = 6 for each group, *p < 0.05, ***p < 0.001, compared with NS at each pressure, Dunn’s post hoc test after two-way ANOVA). (c) Representative EMG recordings from diabetic rats treated with 50 μg/kg CLI-095 responding to graded GD at 60–120 mmHg 4 weeks after STZ injection. (d) AUC rate of EMG recordings from the diabetic rats treated with 50 μg/kg CLI-095 was significantly reduced till 4 h at distention pressures of 60–120 mmHg 4 weeks after injection (n = 11 for each group, *p < 0.05, **p < 0.01, ***p < 0.001, compared with before treatment, Dunn’s post hoc test after two-way ANOVA).
Inhibition of TLR4 reduced the hyper-excitability of gastric-specific DRG neurons
To further confirm the function of TLR4 in gastric hypersensitivity of DM rats, the patch-clamp recording was performed to detect the excitability of gastric-specific DRG neurons of DM rats after intrathecal injection of CLI-095. We observed a significant hyperpolarization of the RP in DRG neurons from CLI095-treated rats. The results showed that RPs were −47.0 ± 0.8 mV and −50.7 ± 1.0 mV for NS- and CLI-095-treated rats, respectively (Figure 3(a)). CLI-095 treatment markedly increased the AP threshold and rheobase. The results showed that AP threshold were −31.3 ± 0.9 mV and −28.7 ± 0.7 mV for NS- and CLI-095-treated rats, respectively (Figure 3(b)). The rheobase measurement was 42.80 ± 2.2 pA and 67.4 ± 3.1 pA for NS- and CLI-095-treated rats, respectively (Figure 3(c)). In addition, CLI-095 treatment greatly reduced the number of APs evoked by 2 × and 3 × rheobase current stimulation. The number of APs evoked by 2 × rheobase current stimulation were 4.1 ± 0.4 and 2.8 ± 0.4 for the NS- and CLI-095-treated groups, respectively. The number of APs evoked by 3 × rheobase current stimulation were 6.6 ± 0.5 and 4.8 ± 0.5 for the NS- and CLI-095-treated groups, respectively (Figure 3(d)–(f)). CLI-095 treatment greatly decreased the number of APs evoked by 100, 300 and 500 pA current ramps. The number of APs evoked by a 100 pA current ramp was 9.1 ± 1.0 and 3.9 ± 0.6 for NS- and CLI-095-treated rats, respectively. The number of APs evoked by a 300 pA current ramp was 19.0 ± 1.5 and 11.8 ± 1.2 for NS- and CLI-095-treated rats, respectively. The number of APs evoked by 500 pA current ramp was 28.4 ± 1.9 and 18.9 ± 1.4 for NS- and CLI-095-treated rats, respectively (Figure 3(g)–(j)). These data indicate that the upregulation of TLR4 enhanced the excitability of gastric-specific DRG neurons. Inhibition of TLR4 reduced the hyper-excitability of gastric-specific DRG neurons. (a) Compared with NS group, CLI-095 injection hyperpolarized the RP in DiI-labeled DRG neurons of diabetic rats (NS, n = 18; CLI-095, n = 21, **p < 0.01, two-sample t-test). (b) Compared with NS group, CLI-095 treatment markedly increased the AP threshold (NS, n = 18; CLI-095, n = 21, *p < 0.05, two-sample t-test). (c) CLI-095 injection caused apparently increase of rheobase compared with NS group on diabetic rats (NS, n = 18; CLI-095, n = 21, ***p < 0.001, two-sample t-test). (d and e) Image involved the examples of AP trances evoked by 2 × and 3 × rheobase current stimulation from NS- (top) and CLI-095-injected diabetic rats (bottom). (f) CLI-095 treatment greatly reduced the number of APs evoked by 2 × and 3 × rheobase current stimulation (2 × rheobase: NS, n = 18; CLI-095, n = 21; 3 × rheobase: NS, n = 18; CLI-095, n = 21; *p < 0.05, two-sample t-test). (g–i) Image indicated the examples of APs by 100, 300 and 500 pA ramp current injection from NS- and CLI-095-injected diabetic rats. (j) CLI-095 treatment greatly decreased the number of APs evoked by 100, 300 and 500 pA current ramps (100 pA: NS, n = 18; CLI-095, n = 21, ***p < 0.001, two-sample t-test; 300 pA: NS, n = 18; CLI-095, n = 21, ***p < 0.001, two-sample t-test; 500 pA: NS, n = 18; CLI-095, n = 21, ***p < 0.001, two-sample t-test).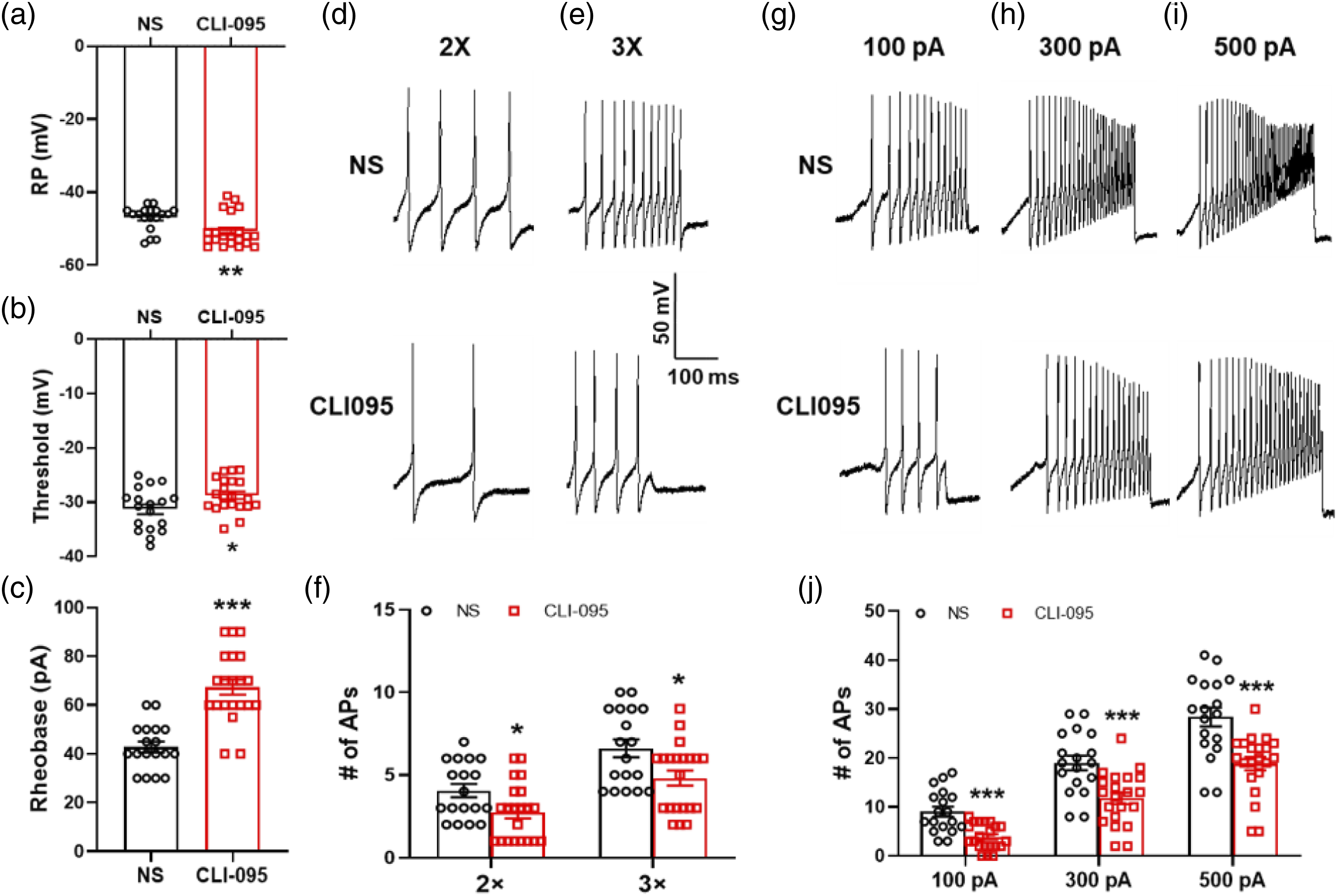
The expression of miRNA-181a was decreased in T7-T10 DRGs and had targeting regulation roles on TLR4
We have initially demonstrated that TLR4 participates in diabetic gastric hypersensitivity. But what is the mechanism of TLR4 up-regulation? Based on the existing research basis of the laboratory, we focus on the perspective of epigenetic regulation. Two microRNAs are predicted and chosen with the forecasting websites and software: TargetScan, miRTarBase, and miRanda, which were miR-181a and miR-7a. To determine whether these microRNAs are involved in diabetic gastric hypersensitivity, QPCR was used to measure their expression in T7-T10 DRGs from DM and CON rats at 4 weeks. The results showed that both miR-181a and miR-7a were significantly decreased compared with CON rats at 4 weeks (Figure 4(a) and (b)). The expression of two microRNAs was decreased in gastric-related DRGs in diabetic rats, suggesting that they may be involved in pain. Meanwhile, by Dual-luciferase reporter gene assay, we found that miR-181a had a direct targeting regulation with TLR4 (Figure 4(c)), while miR-7a did not (Figure 4(d)). The reduction of miR-181a up-regulated TLR4 expression in T7-T10 DRG of diabetic rats. (a) The expression of miR-181a greatly decreased in diabetic rats 4 weeks after STZ injection (n = 6 for each group, **p < 0.01, compared with CON, two-sample t-test). (b) The expression of miR-7a greatly decreased in diabetic rats 4 weeks after STZ injection (n = 6 for each group, ***p < 0.001, compared with CON, two-sample t-test). (c) Dual-luciferase reporter gene assay showed miR-181a had targeting regulation roles on TLR4 (Luc-TLR4-m3′-UTR, n = 4 for each group; Luc-TLR4-3′-UTR, n = 3 for each group, *p < 0.05, compared with NC, two-sample t-test). (d) Dual-luciferase reporter gene assay showed miR-7a had no targeting regulation roles on TLR4 (Luc-TLR4-m3′-UTR, n = 4 for each group; Luc-TLR4-3′-UTR, n = 4 for each group, p > 0.05, compared with NC, two-sample t-test). (e) MiR-181a positive neurons are shown in red. TLR4 positive neurons are shown in green. Merge of double labeling of miR-181a and TLR4. Scale bar = 50 μm. (f) TLR4 protein expression was greatly decreased in miR-181a agomir-treated diabetic rats compared with NC-treated diabetic rats (n = 5 for each group, **p < 0.01, compared with NC agomir, two-sample t-test).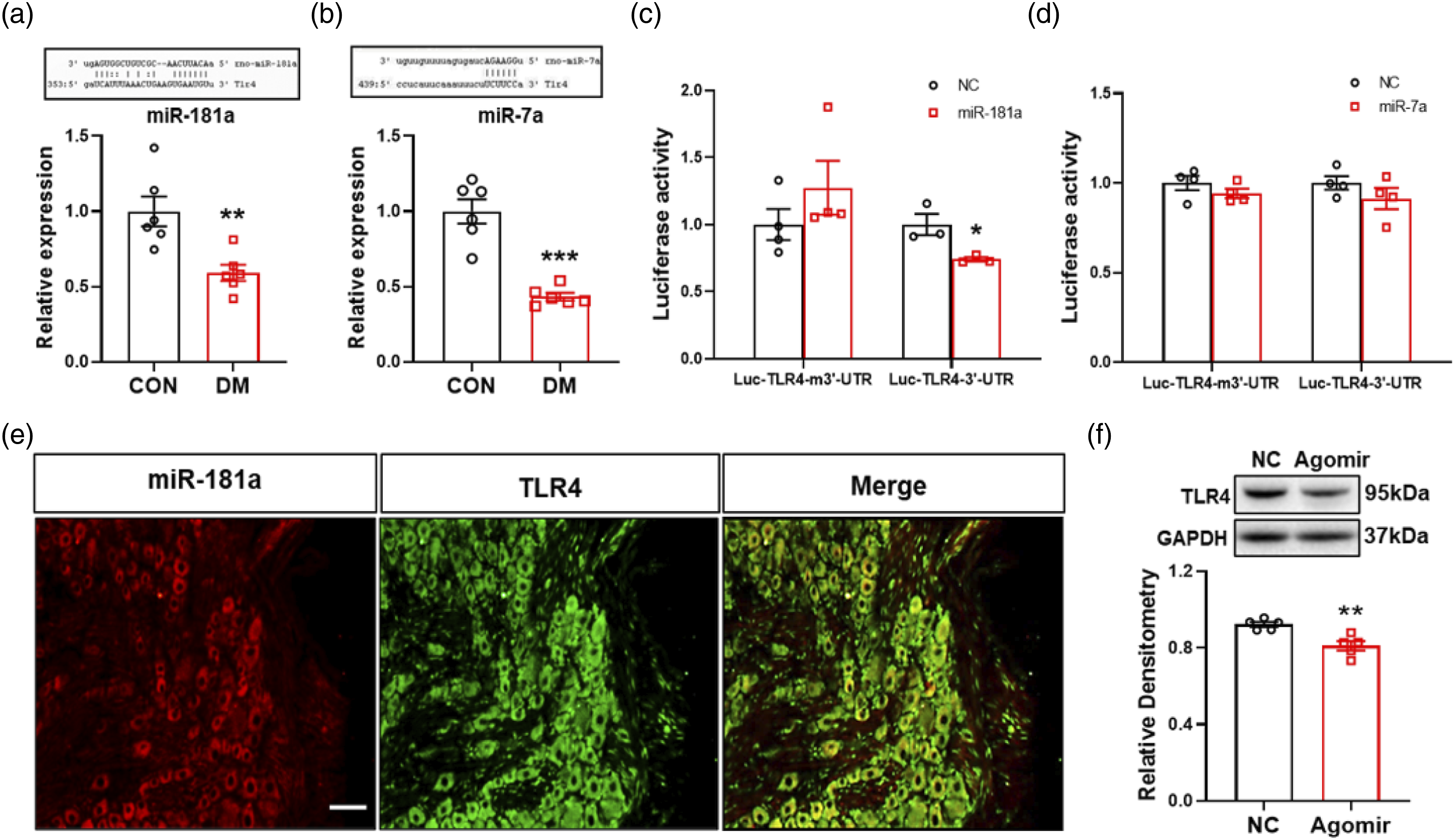
Reduction of miR-181a upregulated TLR4 expression in diabetic rats
We have confirmed that miR-181a could regulate TLR4 directly. Based on these results, we tried to investigate whether miR-181a could influence TLR4 expression in diabetic rats. Firstly, FISH was applied to detect the expression of miR-181a and TLR4 in DRGs of CON rats. The figures showed that miR-181a was co-expressed with TLR4-labeled neurons (Figure 4(e)). Next, we treated diabetic rats with miR-181a agomir and NC agomir, and the western blotting was performed to detect the expression of TLR4 in T7-T10 DRGs. We found that the expression of TLR4 was decreased significantly after injection of miR-181a agomir, compared with age- and sex-matched NC agomir injected rats (Figure 4(f)).
MiR-181a agomir treatment attenuated STZ-induced gastric hypersensitivity
We have already confirmed that loss of miR-181a up-regulated TLR4 expression, which suggested that miR-181a may be involved in the regulation of gastric hyperalgesia in diabetic rats. To prove it, miR-181a agomir was intrathecally injected into diabetic rats. In comparison to the agomir NC group, there was a significant decrease in EMG responses at GD pressures of 60 to 120 mmHg from 0.5 to 2 h, with maximal inhibition occurring at 1 h after the agomir injection (Figure 5(a) and (b). Besides, we intrathecally injected miR-181a agomir for seven consecutive days and tested behavior changes. As shown in Figure 5(c) and (d), the analgesic effect of miR-181a agomir lasted for approximately 48 h, and maximal inhibition occurred at 8 h. MiR-181a agomir treatment attenuated STZ-induced gastric hypersensitivity. (a) Representative EMG recordings from diabetic rats treated with miR-181a agomir responding to graded GD at 60, 80, 100 and 120 mmHg 4 weeks after STZ injection. (b) AUC rate of EMG recordings from the diabetic rats treated with miR-181a agomir was significantly reduced till 2 h at distention pressures of 60–120 mmHg 4 weeks after injection, and baseline normalized to 1 is the NC group (n = 6 for each group, *p < 0.05, ***p < 0.001, compared with agomir NC group, Sidak’s post hoc test following two-way ANOVA). (c) Representative EMG recordings from diabetic rats treated withmiR-181a agomir for 7 consecutive days responding to graded GD at 60–120 mmHg 4 weeks after STZ injection. (d) AUC rate of EMG recordings from the diabetic rats treated with miR-181a agomir for 7 consecutive days was significantly reduced till 48 h at distention pressures of 60–120 mmHg 4 weeks after injection, and baseline normalized to 1 is the NC group (n = 6 for each group, **p < 0.01, ***p < 0.001, compared with agomir NC group, Sidak’s post hoc test following two-way ANOVA).
MiR-181a agomir reduced the hyper-excitability of gastric-specific DRG neurons
Because miR-181a agomir inhibited EMG responses, we next investigated whether miR-181a agomir affects the excitability of gastric-specific DRG neurons of diabetic rats. We observed a significant hyperpolarization of the RP in DRG neurons from miR-181a agomir-treated rats. RPs were −49.0 ± 0.9 mV and −52.7 ± 1.6 mV for NC agomir- and miR-181a agomir-treated rats, respectively (Figure 6(a)). MiR-181a agomir treatment significantly increased the AP threshold. AP thresholds were −29.0 ± 0.6 mV and −27.2 ± 0.4 mV for NC agomir- and miR-181a agomir-treated rats, respectively (Figure 6(b)). MiR-181a agomir treatment also increased rheobase of DM rats. The rheobase measurements were 14.2 ± 1.5 pA and 34.3 ± 4.4 pA for gastric projecting DRG neurons isolated from NC agomir- and miR-181a agomir-treated rats, respectively (Figure 6(c)). Meanwhile, miR-181a agomir treatment greatly reduced the number of APs evoked by 2 × and 3 × rheobase current stimulation. The numbers of APs evoked by 2 × rheobase current stimulation were 2.9 ± 0.4 and 1.8 ± 0.1 for the NC agomir- and miR-181a agomir-treated groups, respectively. The numbers of APs evoked by 3 × rheobase current stimulation were 4.5 ± 0.5 and 2.6 ± 0.3 for the NC agomir- and miR-181a agomir- treated groups, respectively (Figure 6(d)–(f)). MiR-181a agomir treatment greatly decreased the number of APs evoked by 100, 300 and 500 pA current ramps. The numbers of APs evoked by a 100 pA current ramp were 14.9 ± 1.3 and 3.14 ± 0.73 for NC agomir- and miR-181a agomir-treated rats, respectively. The numbers of APs evoked by a 300 pA current ramp were 21.7 ± 1.8 and 10.4 ± 1.4 for NC agomir- and miR-181a agomir-treated rats, respectively. The numbers of APs evoked by a 500 pA current ramp were 26.1 ± 2.2 and15.5 ± 1.9 for NC agomir- and miR-181a agomir-treated rats, respectively (Figure 6(g)–(j)). miR-181a agomir treatment significantly decreased the number of Aps evoked by current ramp stimulation. These data indicate that the down-regulation of miR-181a enhanced the excitability of gastric-specific DRG neurons from diabetic rats. MiR-181a agomir reduced the hyper-excitability of gastric-specific DRG neurons. (a) Compared with NC agomir group, miR-181a agomir injection hyperpolarized the RP in DiI-labeled DRG neurons of diabetic rats (NC agomir, n = 15; miR-181a agomir, n = 14, **p < 0.01, two-sample t-test). (b) Compared with NC agomir group, miR-181a agomir treatment markedly increased the AP threshold (NC agomir, n = 15; miR-181a agomir, n = 14, *p < 0.05, two-sample t-test). (c) MiR-181a agomir injection caused apparently increase of rheobase compared with NC agomir group on diabetic rats (NC agomir, n = 15; miR-181a agomir, n = 14, ***p < 0.001, two-sample t-test). (d and e) Image involved the examples of AP trances evoked by 2× and 3× rheobase current stimulation from NC agomir- (top) and miR-181a agomir-injected diabetic rats (bottom). (f) MiR-181a agomir treatment has greatly reduced the number of APs evoked by 2 × and 3 × rheobase current stimulation (2 × rheobase: NC agomir, n = 15; miR-181a agomir, n = 14; 3 × rheobase: NC agomir, n = 15; miR-181a agomir, n = 14; *p < 0.05, **p < 0.01, two-sample t-test). (g–i) Image indicated the examples of APs by 100, 300 and 500 pA ramp current injection from NC agomir- and miR-181a agomir-injected diabetic rats. (j) MiR-181a agomir treatment greatly decreased the number of APs evoked by 100, 300 and 500 pA current ramps (100 pA: NC agomir, n = 19; miR-181a agomir, n = 16, ***p < 0.001, two-sample t-test; 300 pA: NC agomir, n = 15; miR-181a agomir, n = 14, ***p < 0.001, two-sample t-test; 500 pA: NC agomir, n = 15; miR-181a agomir, n = 14, **p < 0.01, two-sample t-test).
Inhibition of p65 attenuated STZ-induced gastric hypersensitivity
The upstream mechanism of TLR4 had been discussed above, but how does TLR4 up-regulation cause gastric hypersensitivity in DM rats? TLR4 can activate NF-κB through MyD88-dependent and -independent signaling pathways.
26
The inhibition of p65 signaling by the NF-κB inhibitor pyrrolidine dithiocarbamate or recombinant lentiviral vector-p65 small interfering RNA (LV-p65 siRNA) remarkably attenuated diabetic neuropathic pain.
19
Our previous study reported that the expression of p65 (a subunit of NF-κB) was increased significantly in gastric-specific DRG neurons of DM rats, compared with age- and sex-matched CON rats.
4
Firstly, the localization of p65 and TLR4 in CON rats was detected by immunofluorescence analysis. We injected Dil into the gastric area. TLR4 co-expressed with neuron which labeled by Dil. p65 co-expressed with TLR4+ positive neurons (Figure 7(a)–(d)). To further confirm the function of p65, diabetic rats was intrathecal injected with LV-p65 siRNA or LV-NC siRNA, and the EMG responses was recorded. The results showed that AUC rate of LV-p65 siRNA group was markedly decreased at 60–120 mmHg GD pressures at 2 and 4 weeks (Figure 7(e) and (f)). Besides, LV-p65 siRNA had no significant effect on AUC of EMG in age- and sex- matched healthy CON rats (data were not shown). Inhibition of p65 attenuated STZ-induced gastric hypersensitivity. (a) Dil-labled neurons are shown in red. Scale bar = 50 μm. (b) p65-positive neurons are shown in blue. (c) TLR4-positive neurons are shown in green. (d) Merge of three labeling of Dil, p65 and TLR4. (e) Representative EMG recordings from diabetic rats treated with LV-NC siRNA and LV-p65 siRNA responding to graded GD at 60–120 mmHg 4 weeks after STZ injection. (f) AUC rate of EMG recordings from the LV-p65 siRNA group was significantly lower than LV-NC siRNA group at distention pressures of 60–120 mmHg at 4 weeks (LV-NC siRNA, n = 8; LV-p65 siRNA, n = 9, ***p < 0.001, compared with LV-NC siRNA at each pressure, Sidak’s post hoc test after two-way ANOVA).
Inhibition of p65 reduced enhanced excitability of gastric projection DRG neurons
Because LV-p65 siRNA alleviated gastric hypersensitivity in diabetic rats, we next investigated whether LV-p65 siRNA affected the excitability of gastric-specific DRG neurons. RPs were −44.8 ± 0.3 mV and −49.0 ± 0.7 mV for gastric projection DRG neurons isolated from LV-NC siRNA- and LV-p65 siRNA-treated rats, respectively. Therefore, intrathecal injection of LV-p65 siRNA significantly hyperpolarized RP (Figure 8(a)). LV-p65 siRNA treatment also markedly increased rheobase. Rheobase was 43.0 ± 1.4 pA and 71.2 ± 3.4 pA for gastric projection DRG neurons isolated from LV-NC siRNA- and LV-p65 siRNA- treated rats, respectively (Figure 8(b)). The treatment of LV-p65 siRNA greatly decreased the number of APs evoked by 100, 300 and 500 pA ramp current stimulation, compared with LV-NC siRNA. The number of APs evoked by a 100 pA ramp current stimulation was 10.8 ± 0.9 and 5.2 ± 1.0 for LV-NC siRNA- and LV-p65 siRNA-treated groups, respectively. The number of APs evoked by a 300 pA ramp current stimulation was 22.7 ± 1.7 and 15.6 ± 0.8 for LV-NC siRNA- and LV-p65 siRNA-treated rats, respectively. The number of APs evoked by 500 pA ramp current stimulation was 32.1 ± 2.2 and 23.7 ± 1.1 for LV-NC siRNA- and LV-p65 siRNA- treated rats, respectively (Figure 8(c)–(h)). These results showed that inhibition of p65 reduced enhanced excitability of gastric-specific DRG neurons. Inhibition of p65 reduced enhanced excitability of gastric-specific DRG neurons. (a) Compared with LV-NC siRNA group, LV-p65 siRNA injection hyperpolarized the resting membrane potential (RP) in DiI-labeled DRG neurons of diabetic rats (n = 20 for each group, ***p < 0.001, two-sample t-test). (b) LV-p65 siRNA injection caused apparently increase of rheobase compared with LV-NC siRNA group on diabetic rats (n = 20 for each group, ***p < 0.001, two-sample t-test). (c, e and g) Image indicated the examples of APs by 100, 300 and 500 pA ramp current injection from LV-NC siRNA- and LV-p65 siRNA-injected diabetic rats. (d, f and h) LV-p65 siRNA treatment greatly decreased the number of APs evoked by 100, 300 and 500 pA current ramps (n = 20 for each group, 100 pA: ***p < 0.001, two-sample t-test; 300 pA: ***p < 0.001, two-sample t-test; 500 pA: **p < 0.01, two-sample t-test).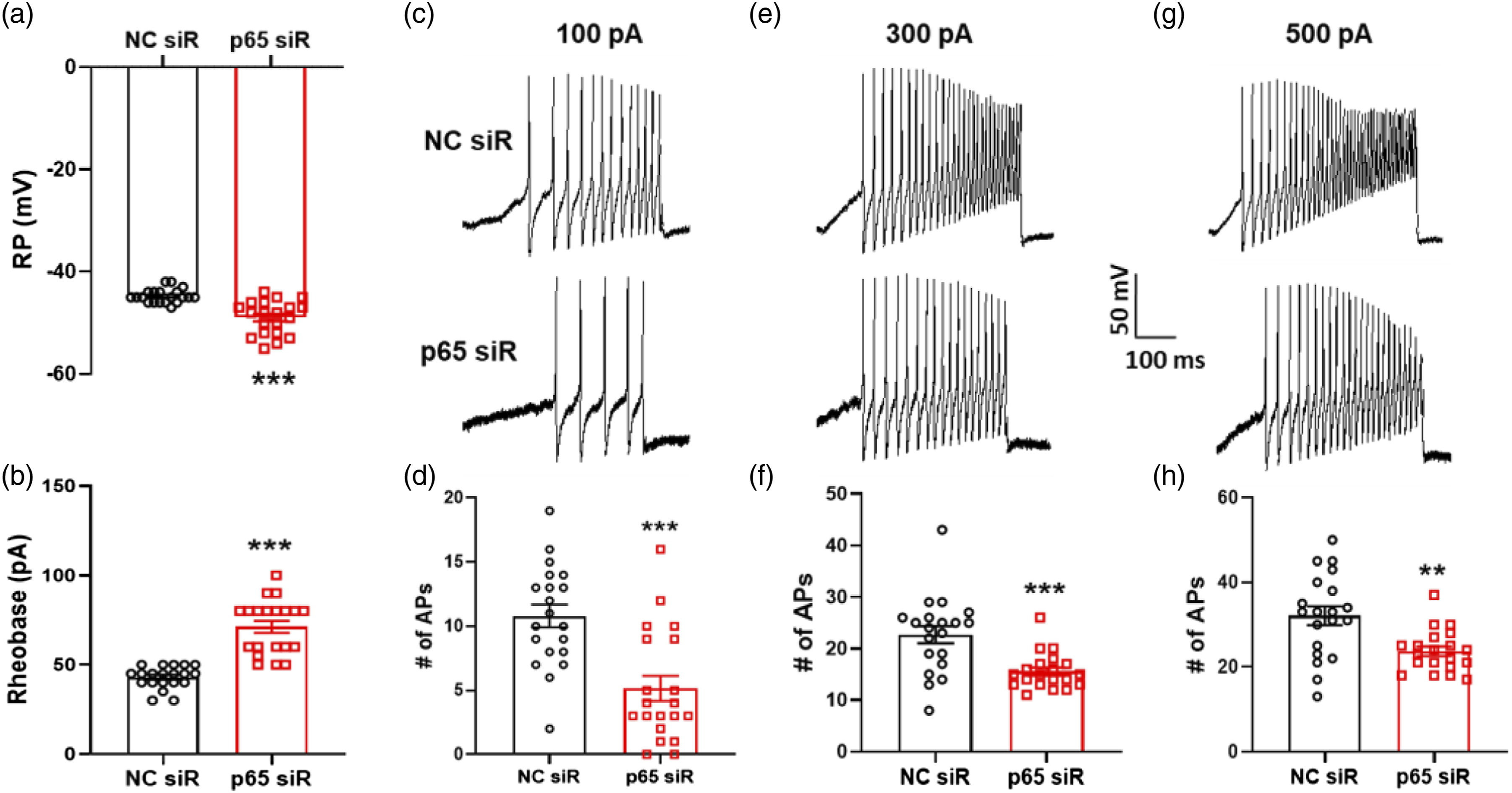
Inhibition of TLR4 reduced p65 and TRAF6 expressions in T7-T10 DRGs
A recent study showed that upregulation of interleukin-1 receptor activated kinase (IRAK1) and tumor necrosis factor receptor-associated factor 6 (TRAF6), two adaptor proteins for the TLR signaling pathway, in DRG neurons was associated with activation of NF-κB in diabetic peripheral neuropathy.
27
Western blotting was performed to detect the expression of TRAF6 in the T7-T10 DRGs. The results showed that the expression of TRAF6 was increased significantly in T7-T10 DRGs of diabetic rats (Figure 9(a)). To determine whether upregulation of p65 resulted from TLR4 activation, western blotting was used to detect the expression of p65 after CLI-095 (50 μg/kg) treatment. CLI-095 treatment significantly reduced the expression of p65 in gastric DRGs isolated from DM rats, and had no effect on TLR4 expression (Figure 9(b)). To further verify whether TRAF6 took part in TLR4’s involvement in gastric hypersensitivity of diabetic rats, CLI-095 was used in this study as described above, which markedly reduced the expression of TRAF6, compared with NS treatment (Figure 9(c)). Thus, TLR4 may upregulate the expression of p65 through TRAF6 signaling pathway in diabetic rats. The expressions of TRAF6 were increased in T7-T10 DRGs, and inhibition of TLR4 reduced p65 and TRAF6 expressions. (a) TRAF6 protein expression was greatly enhanced in diabetic rats compared with CON rats 4 weeks after injection (CON, n = 5; DM, n = 6, *p < 0.05, compared with CON, two-sample t-test). (b) TLR4 protein expression had no changes in CLI-095-treated diabetic rats compared with age- and sex-matched NS- treated diabetic rats (n = 4 for each group, p > 0.05, compared with NS, two-sample t-test). (c) TRAF6 protein expression was greatly decreased in CLI-095- treated diabetic rats compared with NS- treated diabetic rats (NS, n = 3; CLI-095, n = 4, *p < 0.05, compared with NS, two-sample t-test). (d) NF-κB protein expression was greatly decreased in CLI-095- treated diabetic rats compared with NS- treated diabetic rats (NS, n = 4; CLI-095, n = 5, *p < 0.05, compared with NS, two-sample t-test).
Discussion
Upper abdominal pain or discomfort, one of the cardinal symptoms of patients with long-term diabetes, has been a challenge for gastroenterologists to treat effectively. Gastrointestinal symptoms of diabetic patients not only cause blood sugar fluctuations, which in turn triggers and exacerbates other acute and chronic complications, but also leads to severe malnutrition and endangers patients’ lives when the condition worsens. However, the pathogenesis of the disease has not been fully elucidated, and the clinical treatment methods are very limited. Kumar et al. 27 performed intragastric balloon dilation tests on 13 normal people and 18 diabetic patients with obvious upper gastrointestinal symptoms. The researchers found that the pressure threshold of balloon distension-induced abdominal discomfort in diabetes patients was significantly lower than that in normal people, that is, gastric hypersensitivity is present in diabetic patients. Therefore, it is very important to further investigate its pathogenesis and find new potential therapeutic targets.
Because it is difficult to quantitatively evaluate gastric sensation, the research on diabetic gastric hypersensitivity is rare. In our previous study, intragastric balloon dilation technology combined with electromyographic recording technology could be used to successfully detect streptozotocin (STZ) -induced gastric hypersensitivity in type 1 diabetic rats. Our study also found that the STZ-induced diabetes model is stable, 24 with obvious gastric hypersensitivity and small differences among individuals. 4 Therefore, in this study, we continued to use STZ-induced type 1 diabetic rats as animal models.
The primary sensory neuron is located in the dorsal root of the spinal cord. It first responds to surrounding environmental stimuli, and then continuously transmits signals to the central nervous system, causing the entire system to be continuously sensitized. The sensory afferent nerve of the stomach is mainly composed of visceral sensory fibers, and its soma (that is the primary sensory neuron) is located in T7-T10 dorsal root ganglion (DRG). We found in previous studies that NF-κB activates and promotes the endogenous hydrogen sulfide synthase cystathionine-β-synthetase (CBS) expression, then up-regulates CBS-H2S signaling, and induces diabetic gastric hypersensitivity by enhancing the excitability of DRG neurons. 4 Wang et al. 28 showed that interleukin-1 receptor-activated kinase (IRAK1) and tumor necrosis factor receptor-associated factor 6 (TRAF6) are up-regulated in primary sensory neurons and activate NF-κB. IRAK1 and TRAF6 both are the important intermediate molecules in the process of NF-κB activation by Toll-like receptors 4 (TLR4) through myeloid differentiation factor 88 (MyD88) -dependent signaling pathways. We speculate that TLR4 is involved in the activation of NF-κB in T7-T10 DRG neurons. We first examined the increased expression of TLR4 and TRAF6 and then observed the effects of TLR4’s antagonist CLI-095 on gastric hypersensitivity and neuronal excitability of T7-T10 DRG neurons in diabetic rats. The results showed that the CLI-095 could alleviate diabetic gastric pain in rats and reduce the excitability of T7-T10 DRG neurons. The results suggested that the upregulation of TLR4 was involved in gastric hypersensitivity in diabetic rats.
To further confirm the role of TLR4/NF-κB signaling in diabetic visceral pain, we firstly applied the immunofluorescence technique to observe the distribution of TLR4 and NF-κB, and the results showed that TLR4 and NF-κB were co-expressed on gastric-specific DRG neurons. We developed a highly efficient method of lentivirus-mediated delivery of small interfering RNA (LV-siRNA) targeting p65 for gene silencing as described previously, and then intrathecal injection of LV-p65 siRNA (1 × 107 TU) in diabetic rats, observing its effect on diabetic gastric hypersensitivity and DRG neuronal excitability. The results showed that the LV-p65 siRNA could alleviate diabetic gastric pain in rats and reduce the excitability of T7-T10 DRG neurons. The studies suggest that NF-κB not only promotes neuroinflammation,29,30 but also that NF-κB activation in the neuronal body can increase neuronal excitability and is one of the initiating factors that induce diabetic gastric hypersensitivity. Meanwhile, the treatment of CLI-095 could reduce the NF-κB and TRAF6 expression significantly. These data means that the activation of TLR4/NF-κB signaling contributes to diabetic gastric hypersensitivity. Our previous studies have shown that specific activation of TLR4 is associated with the development of diabetic neuropathic pain. 15 Xu et al. found that inhibition of TLR4/NF-κB signaling pathway in spinal cord might help alleviate chronic neuropathic pain in CCI rat models. 31 In visceral pain, enhanced TLR4 expression and microglial activation in prefrontal cortex and hippocampus are involved in visceral pain induced by a high-fat diet. 32 Zhang et al. found that activation of spinal microglial TLR4/MyD88/NF-κB signaling facilitated the development of visceral hypersensitivity in a rat model developed by neonatal and adult colorectal distension. 33 However, to date, no related studies have reported the role of TLR4 in diabetic visceral pain. Our study increases evidences of the role of TLR4/NF-κB signaling pathway in gastric hypersensitivity in diabetic rats.
In addition to LPS, there are many endogenous ligands that can activate TLR4 and its downstream pathways.12,34 Growing studies show that microRNAs can regulate gene expression after transcription, including TLR4. 35 MicroRNA-6869-5p acts as a tumor suppressor by targeting TLR4/NF-κB signaling pathway in colorectal cancer. 36 We first predicted that miRNA181a, miR-7a, miR-218, and miR-23a could complement bind with TLR4 mRNA at its 3′-UTRd through bioinformatics including TargetScan and miRTarBase databases.20,21 Then RT-PCR was used to detect the expression of the above miRNAs in gastric-specific DRGs of diabetic rats (Supplementary Figure S2). The results showed that the expressions of miR-181a and miR-7a were significantly reduced. In order to further study the possible regulatory effect of microRNAs on TLR4, we constructed a dual-luciferase reporter gene system for TLR4 and its mutants, and administered mimics of miR-181a and miR-7a, respectively. The results found that only miR-181a mimics could reduce TLR4 expression. We then applied miR-181a agomir intrathecally and found that it could down-regulate TLR4 expression and partially reduce gastric hypersensitivity and excitability of gastric-specific DRG neurons in diabetic rats. At present, studies showed that miR-181a/TLR4 increased the expressions of inflammatory cytokines, such as IL-6, IL-1β, and TNF-α, and contributed to atherosclerosis 29 and acute lung injury. 37 However, the role of miR-181a/TLR4 in the nervous system is rarely reported. We report for the first time that miR-181a/TLR4 is upregulated in the DRGs of diabetic rats and exerts effects on neuronal excitability and diabetic visceral pain. We expect that our studies may shed light on epigenetic mechanisms of chronic visceral pain in organic gastrointestinal disorders. This added knowledge would provide new strategies for treatment of chronic visceral pain in patients with diabetes.
Limitation
In the present study, a single injection of miR-181a agomir could alleviate diabetic gastric hypersensitivity, suggesting that miRNAs may have a direct regulating effect on neuronal excitability, which needs to be confirmed by further research. In addition, whether miR-181a agomir affects p65 expression or whether miR-181a only affects p65 by regulating TLR4 remains to be further clarified. Furthermore, DRG sensory neurons are only one of the components in the peripheral sensory afferent pathway, and the expression of miR-181a in the vagus nerve is unknown and needs further exploration.
Conclusion
The decreased microRNA-181a in gastric-specific DRGs could up-regulate TLR4 expression. TLR4 activated NF-κB through MyD88-dependent signaling pathway, increased excitability of DRG neurons, and contributed to gastric hypersensitivity in rats with diabetes. The schematic drawing is shown in Supplementary Figure S1. These findings might provide potential targets for clinical treatment.
Supplemental Material
Supplemental Material - microRNA-181a contributes to gastric hypersensitivity in rats with diabetes by regulating TLR4 expression
Supplemental Material for microRNA-181a contributes to gastric hypersensitivity in rats with diabetes by regulating TLR4 expression by Qian Sun, Shiyu Zhang, Bing-Yu Zhang, Yilian Zhang, Lijun Yao, Ji Hu and Hong-Hong Zhang in Molecular Pain.
Footnotes
Author contributions
Qian Sun, Shiyu Zhang and Bing-Yu Zhang performed the experiments, analyzed the data, and prepared figures and the manuscript. These authors contributed to this work equally. Yilian Zhang and Lijun Yao performed the experiments and analyzed the data. Ji Hu revised the manuscript. Hong-Hong Zhang designed and supervised the experiments and finalized the manuscript. All authors contributed to the article and approved the submitted version.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by grants from National Natural Science Foundation of China (82071234 and 31400947 to H-HZ and 82170836 to JH). This work was also supported by grant from the Jiangsu Youth Medical Talents Project (QNRC2016874 to H-HZ) and the project of the clinical research center of neurological disease of the Second Affiliated Hospital of Soochow University (ND2022B01 to H-HZ).
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.