
Abstract
Keywords
Introduction
Opioid-induced hyperalgesia (OIH) is defined as a paradoxical condition of nociceptive sensitization in patients who use opioids to treat acute and chronic pain.1,2 Clinical studies have confirmed that intraoperative use of high-dose opioid agonists, such as morphine and remifentanil, was associated with increased acute pain incidence after surgery, inducing perplexities for the clinical application of opioids.3–5 The mechanisms of OIH are complex, involving both central mechanisms and peripheral mechanisms.6–8 The latest research indicated that peripheral sensitization mainly leads to OIH during the early opioid post-infusion period, while central mechanisms might account for hyperalgesia during the late opioid post-infusion period, indicating that remifentanil might sensitize peripheral and central nociceptive signal to promote chronologically distinctive hyperalgesia. 9 Among these clinical opioids, remifentanil was able to elevate postoperative pain scores featured by remifentanil-induced postoperative hyperalgesia (RIH) and increase opioid consumption. Although numerous contributory molecular and cellular mechanisms had been described in previous studies, the underlying molecular mechanism of OIH/RIH still needs further investigation.1,6
Recent studies have indicated that Toll-like receptor 4 (TLR4) might participate in the development of OIH.10,11 TLR4 is a pattern recognition receptor that initiates the innate immune response to protect the body from invasive pathogens. 12 In addition, the process of TLR4 challenged by various endogenous or exogenous ligands, both in the sensory neuron and dorsal horn glias, would induce pain. 13 Studies have reported that TLR4 and its downstream signals, such as NF-κB, play an important role in acute and chronic pain.14–16 Moreover, opioids could directly activate spinal microglial TLR4, promoting the release of neuroexcitatory immune mediators and following glial “priming”, exacerbating neuropathic pain after opioid treatment. 17 Increasing researches revealed that initiating central sensitization in OIH depended on TLR4 signaling while inhibiting TLR4 or Tlr4 knockout prevented mechanical allodynia in OIH.10,11,17 However, the molecular underlying TLR4-mediated OIH/RIH remains unclear, especially focusing on peripheral sensitization mediated by sensory neurons.
The importance of transient receptor potential (TRP) ion channels in pain sensation has been emphasized by many studies.18,19 The transient receptor potential vanilloid 1 (TRPV1) channel and transient receptor potential ankyrin 1 (TRPA1) channel, which are mainly present in peripheral nociceptors, are well-known nociceptive TRP family members. TRPV1 and TRPA1 served as transducers and amplifiers for pain signals, mediating mechanical allodynia and thermal hyperpathia in neuropathic and inflammatory pain.20–22 Detailedly, TLR4 signaling might cause neuropathic and inflammatory pain by upregulating and sensitizing TRPV1 and TRPA1 in sensory neurons.22,23 The role of sensory neuron TRPV1 in RIH has been well established.24,25 However, whether TRPA1 participated in RIH remains unclear.
In this study, we proposed that TLR4 signaling pathway in sensory neurons modulated the development of RIH via upregulating the TRPA1 channel. To address this hypothesis, we first established a rat RIH model and assessed the expression changes of TLR4 signaling and TRPA1 in DRG neurons. Furtherly, pharmacological inhibition of TLR4 and TRPA1 was applied to test the role of sensory neuron TLR4-TRPA1 axis in RIH. This study might supply potential therapeutic strategies for the pain symptom in RIH and promote appropriate clinical applications of associated opioids.
Materials and methods
Animals
Male Sprague-Dawley (SD) rats aged 5–6 weeks, weighing 240–250 g, were purchased from Beijing Huafukang Bioscience Co. Ltd (Beijing, China). The rats were given water and food ad libitum and were housed in conditions of constant temperature (22 ± 2°C) and humidity (50 ± 15%) with a 12-h light/dark cycle. All animals were adapted to the laboratory environment for at least 3 days before the experiment. This study was approved by the Animal Welfare and Ethics Committee of China-Japan Friendship Hospital (No. zyrhyy21-20-01-4). All experimental procedures were performed in accordance with the guidelines of the Animal Welfare and Ethics Committee of China-Japan Friendship Hospital and the International Association for the Study of Pain.
Experiment protocol
This study consisted of two experiments. A schematic diagram of the protocol is shown in Figure 1. In experiment 1 (Figure 1(a)), 24 rats were randomly divided into four groups (n = 6 for each group): group C (control), group R (rats with remifentanil infusion), group I (rats with plantar incision), and group RI (rats with remifentanil infusion and incision). According to previously reported procedures,
26
rats received remifentanil infusion and a plantar incision to induce a model of RIH. Plantar incision was performed 10 min after remifentanil infusion. After the behavioral tests, the expressions of TLR4, NF-κB (phosphorylated p65 subunit), and TRPA1 were measured by using western blot analysis (n = 4 for each group) or immunofluorescence staining (n = 4 for each group). TRPA1 and TLR4 mRNA levels were detected by RT-PCR (n = 4 for each group). Schematic diagram of the experiment protocol. 
In experiment 2 (Figure 1(b)), 24 rats were randomly divided into four groups (n = 6 for each group): group C (control group), group RI (rats with RIH model), group TAK (i.p. 3 mg/kg TAK-242 in rats with RIH model), and group HC (i.p. 300 mg/kg HC-030031 in rats with RIH model). After the behavioral tests, the expressions of TLR4, NF-κB (phosphorylated p65 subunit), and TRPA1 were measured by western blot analysis (n = 4 for each group) or immunofluorescence staining (n = 4 for each group). Trpa1 and Tlr4 mRNA levels were detected by RT-PCR (n = 4 for each group).
Drugs preparation
TAK-242 (Selleck Chemicals, Shanghai, China) and HC-030031 (Selleck Chemicals, Shanghai, China) were dissolved in dimethyl sulfoxide (DMSO; Sigma Aldrich, USA) and diluted with saline. Rats in group TAK and group H were intraperitoneally (i.p.) injected with TAK-242 and HC-030031 1 h before remifentanil infusion, respectively.
Remifentanil (Yichang Renfu Pharmaceutical Co., Hubei, China) at a dose of 60 μg/kg was dissolved in 0.9% normal saline (NS) to a volume of 0.6 mL. After being anesthetized with 2% sevoflurane (Hengrui Pharmaceutical CO., Shanghai, China) using a nose mask, a 24-gauge cannula was inserted into the caudal vein and flushed with heparinized saline. The rats in groups R, RI, TAK, and HC were infused intravenously with remifentanil at a rate of 1.0 μg·kg−1·min−1 for 60 min 26 using an infusion pump (Harvard Pump 11, Harvard Apparatus, USA). Intravenous saline was infused at a rate of 0.01 mL·min−1 for 60 min as control exposure.
Plantar incision
After being anesthetized with sevoflurane, rats in groups I, RI, TAK, and HC received plantar incisions. According to a previously described procedure, 26 a 1-cm longitudinal incision through the skin and fascia of the right hind paw plantar, starting 0.5 cm from the edge of the heel and extending to the toes, was performed with a number 11 blade after disinfection with 75% alcohol. Then the plantar muscles and tendons were exposed and incised longitudinally. After hemostasis, the skin was sutured with 4–0 silk and covered with erythromycin ointment to prevent infection. Rats in groups C and R had a sham procedure without incision of the right paw.
Pain behavior tests
All measurements were performed in a quiet, temperature-controlled room at 24 h before plantar incision/remifentanil infusion (baseline) and at 2, 6, 12, 24, and 48 h. To evaluate mechanical allodynia, paw withdrawal mechanical threshold (PWMT) was measured using a digital electronic Von Frey (IITC-Life Science Instruments, CA, USA). Rats were placed individually in elevated, inverted transparent boxes with a steel mesh floor (1-cm grid) and allowed to habituate for approximately 15 min or until exploratory behavior ceased. The electronic Von Frey filament, consisting of a plastic tip fitted in a hand-held force transducer, was applied perpendicularly to the mid-plantar surface of the right hind paw with gradually increased force. A positive response was defined as lifting, shaking, or licking the paw following stimulation. The tests were performed 3 times with intervals of approximately 5 min, and the mean thresholds were calculated.
To evaluate thermal hyperalgesia, radiant heat was applied to the plantar surface of the right hind paw to measure paw withdrawal thermal latency (PWTL). Rats were placed into a moveable transparent plastic cage on an elevated clear glass plate and allowed to habituate for approximately 15 min. A radiant heat source (50-W projector lamp) mounted on a movable holder below the clear, smooth glass floor was placed directly under the plantar surface of the right hind paw. PWTL was defined as the time from the onset of the infrared heat stimulus to the paw withdrawal from the heat source. Before starting the experiment, the infrared heat intensity of the apparatus was adjusted to give an average PWTL of approximately 15 s, and the cutoff latency was set at 20 s to avoid tissue damage. The tests were performed 3 times with intervals of approximately 5 min, and the mean latencies were calculated.
Western blot analysis
After completing the last behavioral tests at 48 h, all rats were sacrificed under deep anesthesia (5% sevoflurane). According to the literature, 27 the L3-L5 right DRGs were rapidly removed and stored in liquid nitrogen. Tissue samples were homogenized in lysis buffer, which was centrifuged at 12,000 r/min for 10 min at 4°C to collect the supernatant. The protein concentration was measured by BCA Protein Assay Kit (Solarbio, Beijing, China). Protein samples were separated on SDS-polyacrylamide gel electrophoresis, and the fractionated proteins were transferred to a polyvinylidene difluoride (PVDF) membrane (Thermo Fisher Scientific, USA). After being blocked with 5% (w/v) skimmed milk for 1 h at room temperature, the membranes were incubated overnight at 4°C with the diluted primary antibodies. The primary antibodies used were: anti-TRPA1 (GTX54765, GeneTex, USA), anti-TLR4 (GTX57153, GeneTex, USA), anti- NF-κB (ab194726, Abcam, USA), and anti-GAPDH (ab8245, Abcam, USA). The membrane was washed with Tris-buffered saline with Tween buffer and incubated with the goat anti-mouse secondary antibody (ab150113, Abcam, USA) or goat anti-rabbit secondary antibody (ab150080 Abcam, USA) for 1 h at room temperature. The bands were scanned by ECL Western Blotting Substrate (Tanon, China) and analyzed by Image J software.
Immunofluorescence staining
Rats were terminally anesthetized with sevoflurane and perfused transcardially with PBS followed by 4% paraformaldehyde. After perfusion, the L3-5 DRGs were collected, post-fixed in 4% paraformaldehyde overnight at 4°C, and then dehydrated in sucrose. Later, tissues were embedded in OCT (Tissue-Tek, Japan) and serially sectioned in a cryostat into 15-μm-thick slices. The tissue sections were blocked with 10% goat serum in 0.3% Triton for 1 h at room temperature and incubated primary antibodies (anti-TRPA1, GTX54765, GeneTex, USA; anti-TLR4, GTX57153, GeneTex, USA) overnight in a wet box at 4°C. Subsequently, the sections were incubated with corresponding secondary antibodies for 1 h at room temperature. The stained sections were visualized under a fluorescence microscope (Leica Microsystems, Wetzlar, Germany). Three DRG tissues (L3-L5) in each tested rat were taken for IF staining and three sections were randomly selected in each tested DRG. Moreover, three rats were randomly selected in each group for IF evaluation. The images for each target were captured under same exposure time and fluorescence intensity in each image was quantized by Image J software for data analysis.
Real-time polymerase chain reaction (RT-PCR)
The expression of Trpa1 and Tlr4 mRNAs, with Gapdh mRNA as an internal control, were detected by RT-PCR. Briefly, total RNA was isolated with TRIzol (TakaRa Bio-technology, Dalian, China). Next, RNA was reverse-transcribed into cDNA with a cDNA Reverse Transcription kit. RT-PCR was performed using SYBR Green dye (Thermo Scientific Molecular Biology) according to the manufacturer’s instructions. The PCR thermocycling parameters were as follows: 50°C for 2 min, 94°C for 15 min, followed by 40 cycles at 94°C for 15 s, 58°C for 30 s, and 72°C for 30 s. The sequences of the oligonucleotide primers were listed as follows: Gapdh (forward: 5′-CAA GGC TGA GAA TGG GAA GC-3′; reverse: 5′-GAA GAC GCC AGT AGA CTC CA-3′), Tlr4 (forward: 5′-TAG CCA TTG CTG CCA ACA TC-3′; reverse: 5′-ACA CCA ACG GCT CTG GAT AA-3′), and Trpa1(forward: 5′-CCA CCC TGT GTG TAG GGA AT-3′; reverse: 5′-AAG GCC ATT CCA GGC TGT AT-3′). Relative quantification of the target gene was calculated with the 2–ΔΔCt method.
Statistical analysis
All data are presented as the mean ± standard deviation (SD). Data were analyzed for normal distribution using the D'Agostino and Pearson omnibus normality test and for homogeneity of variances using Bartlett’s test of sphericity. If data fulfilled the normal distribution and homogeneity of variance test, one-way or two-way analysis of variance (ANOVA) followed by the Tukey’s post hoc test was used to evaluate the presence of significant differences between groups, and Bonferroni-corrected repeated measures analysis of variance was used for intra-group comparison. If data did not follow a normal distribution and homogeneity of variance test, rank-sum test would be performed to measure data from multiple groups. GraphPad Prism 9.0 software (GraphPad Software, CA, USA) was used for all statistical analyses and graphing. p < .05 was considered to be statistically significant.
Results
Remifentanil facilitated pain behaviors and upregulated TLR4 signaling in DRGs
First, in experiment 1, pain behaviors were recorded to verify whether remifentanil exposure would aggravate mechanical allodynia and thermal hyperalgesia. There was no significant difference in the baselines of PWMT and PWTL among all groups before surgery (p > .05). Compared with the baseline, PWMT and PWTL did not present significant changes in the normal control group at the observed time points. Compared with the baseline and normal control group, both PWMT and PWTL were reduced from 2 h to 48 h in rats receiving either incision or remifentanil treatment (p < .05, n = 6; Figures 2(a)–(b)). Moreover, rats in group RIH displayed a significant decrease in PWMT and PWTL compared with the incision group, which started from 2 h and peaked at 48 h. These above results indicated that intraoperative remifentanil exposure facilitated hyperalgesia induced by plantar incision and therefore the rat model of remifentanil-induced postoperative hyperalgesia was successfully established. Time course of PWMT and PWTL after remifentanil and incision. PWMT, paw withdrawal mechanical threshold; PWTL, paw withdrawal thermal latency. 
To determine the expression of TLR4 signaling in DRG neurons, we performed western blot to evaluate the protein levels of TLR4 and phosphorylated NF-κB, the activated form of NF-κB, in DRG. Western blot analysis exhibited a significant increase of TLR4 and phosphorylated NF-κB expression in DRG as remifentanil infusion and plantar incision compared with saline infusion. There was no significant difference in TLR4 or phosphorylated NF-κB expression between groups R and I (p > .05). Furthermore, TLR4 and phosphorylated NF-κB were upregulated in group RIH compared to group R or I (Figure 3(a)–(c)). Consistent with the results of Western blot analysis, the mRNA levels of Tlr4 exhibited a similar expression pattern (Figure 3(e)). We further performed immunofluorescence analysis to confirm the TLR4 signaling alteration in DRG neurons. As shown in Figure 4, TLR4 was mainly expressed in DRG neurons, and fluorescence intensity quantification further confirmed that RIH upregulated TLR4 in DRG (p < .01). Together, these results indicated that TLR4 signaling was activated in DRG neurons in the setting of remifentanil-induced hyperalgesia. Expression of TLR4 signaling and TRPA1 in DRG neurons in rats with RIH. Immunofluorescence of TLR4 and TRPA1 expression in DRG neurons. 

The expression of TRPA1 was increased in remifentanil-induced hyperalgesia rats
To determine the changes in TRPA1 expression following remifentanil exposure, we performed western blot, RT-PCR, and immunofluorescence labeling in the DRG (Figure 3(a)) shows that the expression of TRPA1 was increased in both group R and group I as compared with the normal control group. However, there was no significant difference between group R and group I (Figure 3(d)). Furthermore, group RI showed a further upregulation of TRPA1 as compared with both group R (p < .01) and group I (p < .01). In addition, the mRNA level of Trpa1 in DRG was also detected. RT-PCR analysis revealed that compared with the normal control group, both group R and group I showed a dramatically increased mRNA level of Trpa1. However, no significant difference was detected between group R and group I. Furthermore, the mRNA level of Trpa1 was enhanced in the RIH group as compared with both I group and R group (Figure 3(e)). Furthermore, immunofluorescence labeling revealed that TRPA1 was abundantly and selectively expressed in DRG neurons and TRPA1-positive cells were almost co-stained with TLR4. Immunofluorescence intensity quantification further confirmed the upregulation of TRPA1 in group R and group I as compared with group C (p < .05), and in RIH group as compared with group R and group I (all p < .05). Together, the above results revealed significant upregulation of TRPA1 in DRG sensory neurons of RIH model.
Inhibition of TRPA1 alleviated mechanical hyperalgesia in RIH rats
To clarify whether TRPA1 regulated RIH, TRPA1 antagonist HC-030031 was injected intraperitoneally, followed by pain behaviors recording. The results showed that compared with the rats in group RI, thresholds of PWMT in group HC were remarkably increased after TRPA1 antagonist injection. However, PWTL threshold elevation was only observed at 48 h between the group RIH and group HC (p < .05) (Figure 5). Effects of inhibiting TLR4 or TRPA1 on mechanical and thermal sensitivity. PWMT, paw withdrawal mechanical threshold; PWTL, paw withdrawal thermal latency. 
To further explore the expression of TLR4 and TRPA1 following HC-030031 administration in RIH model, western blot and immunofluorescence were implemented. The western blot analysis showed that the protein level of TRPA1 was significantly downregulated in group HC as compared with group RI (p < .01), while the expression levels of TLR4 and phosphorylated NF-κB in group HC were not altered by HC-030031 (p > .05) (Figure 6(a)–(d)). Similar to the results of the western blot, immunofluorescence and its scatter plot analysis also demonstrated a significant TRPA1 downregulation in DRG from group HC compared with group RI (p < 0.05) (Figure 7(a) and (c)). Meanwhile, RT-PCR analysis revealed that the mRNA level of Trpa1, but not Tlr4, was significantly decreased in DRG from group HC compared with group RI (Figure 6(e)). Expression of TLR4 signaling and TRPA1 in DRG neurons after inhibiting TLR4 or TRPA1 in RIH model. Immunofluorescence of TLR4 and TRPA1 expression after inhibiting TLR4 or TRPA1 in DRG neurons. 
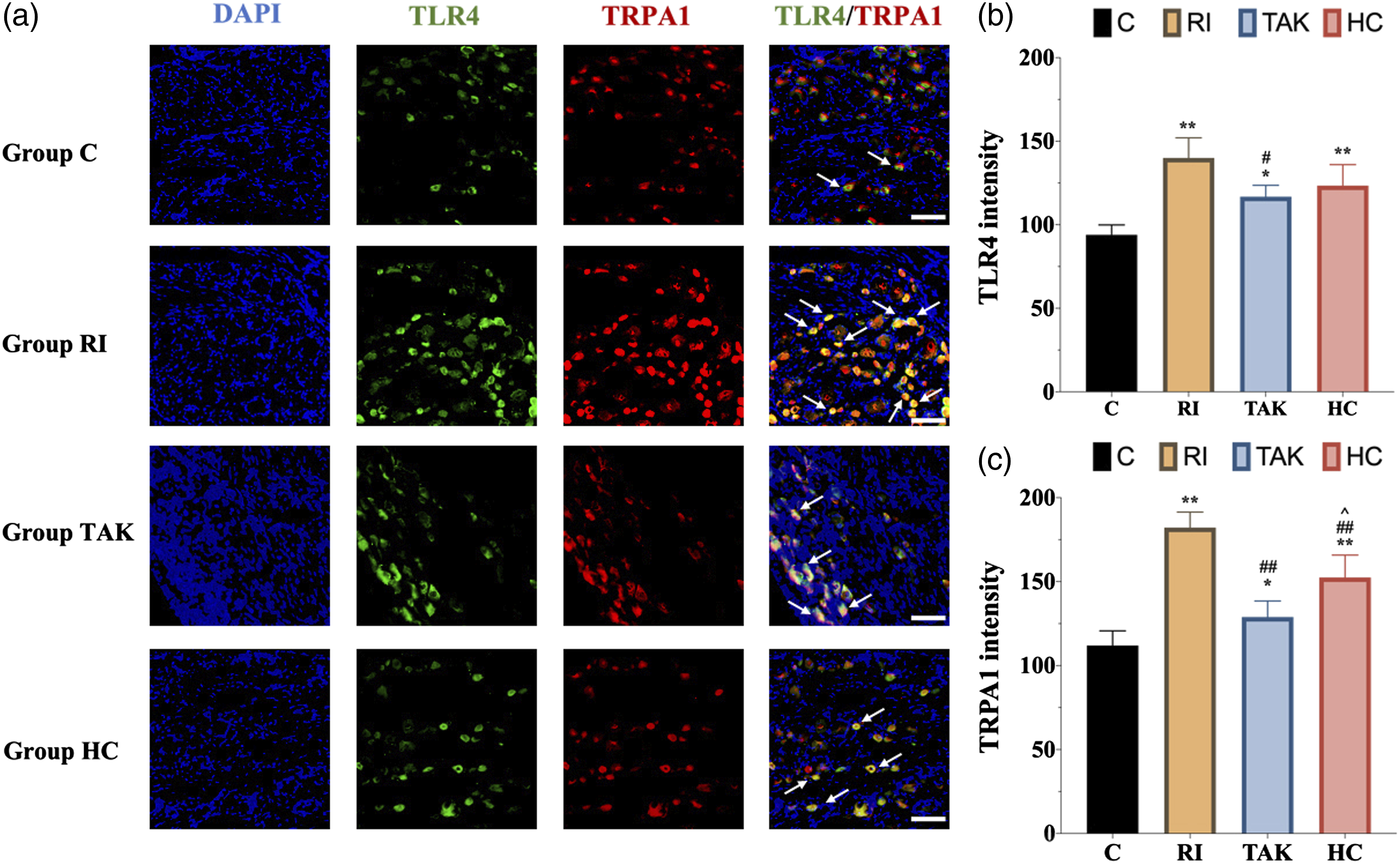
Inhibition of TLR4 alleviated mechanical hyperalgesia and downregulated the expression of TRPA1 in RIH rats
To further confirm the role of TLR4 signaling in RIH, we conducted intraperitoneal injection of TLR4 antagonist TAK-242 to block TLR4 signaling, then recorded pain-associated behavior. As shown in Figure 5, compared with the group RI, TAK-242 pretreatment presented significantly higher threshold of PWMT at different points (all p < .05) in group TAK. However, TAK-242 pretreatment in group TAK did not affect the threshold of PMWL when compared with group RI (p > .05). We further examined the protein expression of TLR4 and activated p-NF-κB. Both western blot and immunofluorescence showed that the protein expression of TLR4 was dramatically decreased in rats of group TAK at 48 h, together with decreased p-NF-κB content (Figure 6(a)–(c)). Similar results were obtained on the mRNA level of Tlr4 in group TAK compared with group RI (p < .05) (Figure 6(e)). These data indicated that TLR4 signaling mediated remifentanil-induced mechanical allodynia.
To figure out whether TRPA1 was the potential downstream regulatory target of TLR4 signaling, we further explored the expression of TRPA1 following TAK-242 administration. Both western blot (Figure 6(a) and (d)) and immunofluorescence (Figure 7) showed that the protein expression of TRPA1 was decreased in group TAK compared with group RI (p < .01). In addition, RT-PCR showed that the mRNA level of Trpa1 was downregulated after TAK-242 administration (p < 0.01) (Figure 6(e)).
Discussion
In the present study, we verified that sensory neurons’ TLR4 signaling pathway indeed participated in the progression of RIH via upregulating TRPA1 channel as follows: (1) Remifentanil-induced mechanical allodynia and thermal hyperalgesia, which were accompanied by upregulation of TLR4 signaling and TRPA1 in DRG neurons; (2) Inhibition of TLR4 attenuated mechanical allodynia in RIH and downregulated the expression of TLR4 signaling and TRPA1 in DRG neurons; (3) Inhibition of TRPA1 alleviated mechanical allodynia together with decreased TRPA1 expression in DRG,
Previous studies have proved that TLR4 signaling pathway in the spinal cord plays an important role in OIH.11,17 Moreover, Aguado D et al. 10 found that the mechanical threshold after infusion of remifentanil was elevated in Tlr4-deficient mice as compared with wild type mice. Our study further identified that RIH upregulated the expression of TLR4 pathway in DRG neurons, and TLR4 antagonist mitigated mechanical allodynia in RIH model. All the above evidence suggested that opioids could induce the activation of TLR4 signaling in sensory neurons and subsequently lead to peripheral sensitization in RIH. Although opioids mainly exert analgesic effects by binding to μ-opioid receptors, multiple studies have verified that opioids can directly activate TLR4 signaling. 28 It has been reported that opioids, including morphine and remifentanil, could non-stereo selectively bind to the TLR4/MD2 complex in glia and DRG neurons. In the current RIH model, opioids might activate the TLR4 downstream signals and finally phosphorylated and activated NF-κB in sensory neurons, which induced the release of proinflammatory mediators and regulated ion channels expression and/or functional status, then subsequently led to nociception and hyperalgesia.17,29 In addition, opioids could indirectly stimulate TLR4 by inducing endogenous TLR4 ligands, which could further challenge TLR4 in sensory neurons and glias. 28 Moreover, previous studies confirmed that blocking TLR4 could significantly alleviate mechanical allodynia in OIH while the analgesic effects of opioids were still preserved.6,30 Given the above factors, the progression of OIH/RIH might be TLR4-dependent, and TLR4 could serve as a promising therapeutic target for OIH/RIH.
TRPA1, which is functionally expressed in peripheral pain-sensing neurons, has been recognized as a pivotal sensor of pain signals. 19 It is known that the expression of TRPA1 in DRG was upregulated in various pain conditions, including inflammatory and neuropathic pain.31,32 In addition, either genetic deletion or pharmacological inhibition of TRPA1 mitigated pain responses.33,34 However, unlike that of TRPV1,24,25 the role of TRPA1 in RIH has not yet been thoroughly investigated. In Experiment 1, we confirmed that the TRPA1 expression in DRG neurons was upregulated in RIH model, revealing a link between increased sensory TRPA1 expression and RIH. In Experiment 2, we further discovered that the mechanical allodynia in RIH was significantly alleviated by TRPA1 antagonist HC-030031, indicating that TRPA1 underlies the ionic basis of mechanical allodynia in OIH. Collectively, our data suggested that TRPA1 upregulation in DRG neurons might mediate peripheral sensitization in RIH.
Interaction between TLR4 signals and TRPA1 has been investigated in prior studies, indicating that TLR4 could affect TRPA1 at both the gene expression and ion channel activity levels .35–37 Accumulating evidence suggested that activation of TLR4 signaling pathway upregulated TRPA1 expression and enhanced TRPA1 sensitivity.35,36 Further studies found that activation of TLR4 and its downstream signaling in sensory neurons triggered the production of neuroinflammatory mediators such as TNF-α and IL-6, which would further contribute to neuropathic and cancer pain via upregulating TRPA1.23,37 In this study, we observed that TLR4 co-expressed with TRPA1 in DRG sensory neurons. Moreover, we found that blocking TLR4 decreased TRPA1 expression in DRG neurons and mitigated mechanical sensitivity after remifentanil exposure. However, no significant changes in TLR4 signaling were observed as inhibiting TRPA1. All these data indicated that sensory neuron TLR4 signaling mediated RIH via upregulating the TRPA1 channel.
Notably, we found that both TLR4 and TRPA1 inhibition significantly alleviated mechanical allodynia rather than thermal hyperalgesia in RIH rats. Similar observations have been previously reported in inflammatory and neuropathic pain rodent models .38,39 Although TRPA1 serves as a key detector of mechanical and chemical stimuli, the thermal sensitivity of TRPA1 remains controversial. 40 Recent evidence put forward the hypothesis that mammalian TRPA1 may be activated by noxious cold and noxious heat,41,42 which was not in accordance with this hypothesis. These data suggested that thermal hyperalgesia in RIH might result from other mechanisms independent of TLR4 and TRPA1, which should be further investigated in the following studies.
There are two limitations of our present study. First, TLR4-mediated upregulation of TRPA1 in the DRG neurons could not fully explain the thermal hyperalgesia in RIH, which could be further determined in the following studies. Second, TRPA1 activity and TRPA1 trafficking were not tested in the current study. Earlier studies have suggested that TRPA1 translocation to the plasma membrane might represent functional activation and sensitization.43,44 Therefore, further research could clarify the alteration of TRPA1 functional status in RIH and the role of TLR4 in this process.
Conclusions
The present study suggests that TLR4 signaling and TRPA1 in the DRG neurons probably play significant roles in developing mechanical allodynia in RIH. The results presented here also demonstrate that TLR4 signaling engages in RIH by amplifying the expression of TRPA1 in the DRG neurons.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the Fundamental Research Funds for the Central Universities (No. 3332022079), and the Scientific Research Fund of China-Japan Friendship Hospital (No. 2017-RC-3 and No. 2018-2-QN-28).