
Abstract
Neuropeptide W (NPW) messenger ribonucleic acid (mRNA) and NPBW1 and/or NPBW2 mRNA are expressed in the descending pain inhibitory system. In the present study, we examined whether NPW microinjected into the descending pain inhibitory system, such as the periaqueductal gray (PAG), locus coeruleus (LC), and rostral ventromedial medulla (RVM), produces an analgesic effect using a rat formalin test. Microinjections of NPW into the PAG ipsilateral and contralateral to the formalin-injected side, LC ipsilateral and contralateral to the formalin-injected side, and RVM produced an analgesic effect. In the RVM study, the analgesic effect was antagonized by WAY100135, a 5-HT1A antagonist, and enhanced by prazosin, an α1 antagonist, and SB269970, a 5-HT7 antagonist. Naloxone, an opioid antagonist, also antagonized the effect of NPW in the RVM study. In the ipsilateral LC study, the analgesic effect was antagonized by WAY100135, idazoxan, an α2 antagonist, and naloxone and was enhanced by prazosin and SB269970. In the contralateral LC study, the analgesic effect was antagonized by prazosin, idazoxan, SB269970, and naloxone. The analgesic effect was antagonized by WAY100135, SB269970, idazoxan, and naloxone in the ipsilateral and contralateral PAG studies. These findings strongly suggest that NPBW1/W2 activation by NPW microinjection into the RVM, LC, and PAG affect the descending pain modulatory system and produce anti-nociceptive and pro-nociceptive effects in the rat formalin test.
Keywords
Opioid-somatostatin-like receptor genes, including NPBW1 and NPBW2 (previously known as GPR7 and GPR8, respectively 1 ), were first cloned in 1995 as orphan receptors. 2 NPBW1 and NPBW2 share 70% nucleotide and 64% amino acid identities. 3 Neuropeptide B (NPB) and neuropeptide W (NPW) have been identified as endogenous ligands for NPBW1 and NPBW2.4,5 NPW has two isoforms, NPW23 and NPW30, which are 23 and 30 amino acids in length, respectively. It has been reported that NPB mainly activates NPBW1, whereas NPW stimulates both receptors with similar potency. 6
It has been reported that the NPB/NPW signaling system has several physiological functions, including regulation of feeding activity, neuroendocrine functions with central regulation, energy homeostasis, autonomic regulation, activation of the stress axis, pain processing, and emotion-related autonomic and neuroendocrine responses. 7 Concerning pain processing, NPB gene knockout mice demonstrate hyperalgesia in response to inflammatory pain. 8 Moreover, we have previously reported that the intrathecal (IT) administration of either NPW or NPB produced an analgesic effect in both rat inflammatory and neuropathic pain models and that intracerebroventricular (ICV) injections of either NPW or NPB had no effect on a rat neuropathic pain model.9,10 These findings suggest that the role of NPBW1/W2 and the NPW/NPB system in spinal pain transmission are different from their role in supra-spinal pain transmission.
In the brain, NPW mRNA and/or NPBW1 mRNA has been reported to be expressed in the periaqueductal gray (PAG), pons, and medulla oblongata. 6 Moreover, NPB/NPW binding was observed in the locus coeruleus (LC) 11 and PAG. 1 The PAG influences descending pain modulation through its reciprocal connections with the rostral ventromedial medulla (RVM). 12 The RVM, located in the medulla oblongata, plays an important role in the spinal serotonergic modulation of pain. The LC acted as the center of a descending noradrenergic pain modulatory system. 12
In the present study, to clarify the role of NPBW1/W2 and the NPW system in the descending pain modulatory system, we investigated the effects of microinjections of NPW into the RVM, LC, and PAG on pain modulation in the rat formalin test, an inflammatory pain model.
Methods
This study was performed according to a protocol approved by the Institutional Animal Care Committee of Kumamoto University, Kumamoto, Japan. Male Sprague-Dawley rats (250–300 g; Japan SLC, Inc., Shizuoka, Japan) were housed individually, maintained on a 12 h dark-light cycle, and provided food and water ad libitum. The animals were handled on arrival and housed for at least three days before catheter implantation. The animals were euthanized immediately after the behavioral study.
RVM, LC, and PAG cannula placements
Injection cannulae were implanted into the RVM, LC, and PAG under isoflurane anesthesia. The rats were placed in a stereotaxic apparatus (Kopf Model 900, David Kopf Instruments, CA). Stainless steel 26G thin wall guide cannulae (C315G, Plastics One, VA) were stereotaxically implanted into the RVM, LC, and PAG through a burr hole (RVM: AP: −11.0 mm, L: 0.0 mm, H: −10.1 mm from bregma; LC: AP: −10.0 mm, L: 1.4 mm, H: −7.2 mm from bregma; PAG: AP: −7.6 mm, L: 0.8 mm, H: −5.0 mm from bregma). Guide cannulae were affixed to the skull with stainless steel screws and cranioplastic cement.
The formalin test was performed seven days after the cannula implantation. All animals displayed normal feeding and drinking behaviors postoperatively. Rats showing neurological deficits after cannula implantation were not included in the study.
After the experiment, 0.5 μl of India ink was injected through the internal cannula after the rats were euthanized. The brains were fixed with paraformaldehyde, and coronal tissue sections were Nissl-stained to confirm the proper injection site. Only rats whose microinjection site was located within the intended nucleus were included in the results.
Drugs and injection
Drugs were dissolved in saline. NPW30 (molecular weight = 3559) was purchased from the Peptide Institute Inc. (Osaka, Japan). WAY100135, a 5-HT1A receptor antagonist, SB269970, a 5-HT7 receptor antagonist, prazosin, an α-1 adrenergic receptor antagonist, idazoxan, an α-2 adrenergic receptor antagonist, and naloxone, an opioid receptor antagonist, were purchased from Sigma-Aldrich (MO).
NPW30 was administered into the RVM, LC, or PAG at a total volume of 0.5 μL for 60 s using a microsyringe pump (EP-60, EICOM, Japan) and a 30G stainless steel internal cannula (C315G, Plastics One, VA) was connected via a polyethylene tube to a 10 μL Hamilton syringe.
In our preliminary study, we found that 1 μg of NPW30 microinjected into the RVM (p > 0.7) and contralateral PAG (p > 0.5) had no analgesic effect as compared with saline-treated rats when injected 10 min before the formalin injection. In the ipsilateral LC study, the analgesic effect in the NPW30 1 μg treated rats was the same as that of NPW30 5 μg treated rats (p > 0.05) when injected 10 min before the formalin injection. The analgesic effect of 1 μg of NPW30 was less than that of 5 μg of NPW30 in the contralateral LC (p < 0.001) and the ipsilateral LC study (p < 0.001) when injected 10 min before the formalin injection. Therefore, we decided that 5 μg of NPW30 should be injected directly into the RVM, LC contralateral or ipsilateral to the formalin-injected side, and PAG contralateral or ipsilateral to the formalin-injected side 10 min before the formalin injection into the footpad. To test the mechanisms of action of NPW30, one of the following drugs was injected 10 min before the injection of NPW30: WAY100135 (46 μg/10 μL, intrathecal [IT]), SB269970 (10 μg/10 μl, IT), prazosin (25 μg/10 μL, IT), idazoxan (30 μg/10 μL, IT), or naloxone (1 mg/kg, intraperitoneal [IP]). The doses of WAY100135, SB269970, prazosin, idazoxan, and naloxone were determined based on previous reports.13–16 Idazoxan, prazosin, WAY100135, and SB269970 were injected via the L5-L6 intervertebral space under 2% isoflurane anesthesia using a 30G needle.
Formalin test
In the formalin test, 50 μL of 5% formalin was injected subcutaneously into the dorsal surface of the hind paw with a 26G needle under brief isoflurane anesthesia. Within 1 min after the formalin injection, spontaneous flinching of the injected paw was observed. Flinching was readily discriminated as a rapid and brief withdrawal or flexion of the injected paw. This pain-related behavior was quantified by counting the number of flinches for 1 min periods at 5 min intervals, 0–60 min after the injection. Two phases of spontaneous flinching behavior were observed: an initial acute phase (phase 1: during the first 6 min after formalin injection) and a prolonged tonic phase (phase 2: beginning approximately 10 min after formalin injection). The phase 1 response is mediated by the direct stimulation of nociceptors by formalin, and the phase 2 response is mediated by the inflammatory response induced by formalin.
Statistical analysis
The time-response data are presented as the mean flinches ± SEM per min for the periods between 1–2 min and 5–6 min after the formalin treatment and then for 1 min periods at 5 min intervals up to 60 min. The data from the phase 1 (0–6 min) and phase 2 (10–60 min) observations were analyzed separately. The effects of NPW30 on formalin-evoked flinches during phase 1 and phase 2 were compared with those of saline using a two-way analysis of variance (ANOVA). For multiple comparisons, the Holm-Sidak method was used. In the antagonist study, a two-way ANOVA was also used. A p-value of <0.05 was considered statistically significant.
Results
In saline-treated control rats, the subcutaneous injection of formalin resulted in a highly reliable biphasic display of flinching of the injected paw, and this behavior was comparable to that previously reported. 17
The injection sites in the RVM, LC, and PAG are shown in Figure 1. India ink (0.5 μL) injected through the internal cannula spread to the RVM, LC, and PAG.
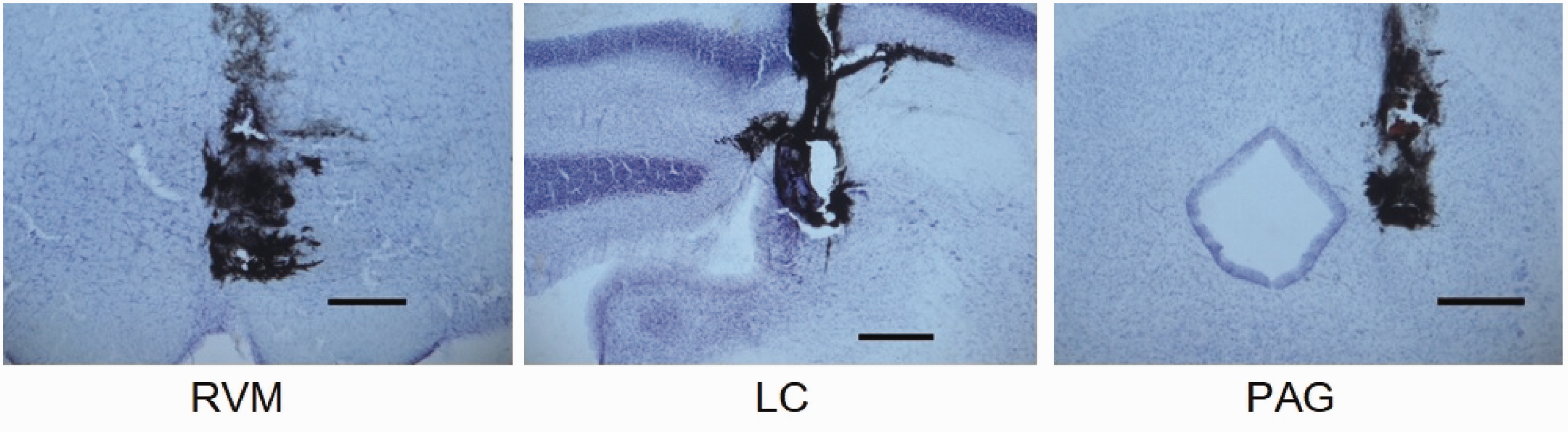
Injection sites in the RVM, LC, and PAG. The spread of 0.5 μl of India ink was injected through the internal cannulae. Nissl-stained sections were used to confirm the injection sites. RVM, LC, and PAG were located within the regions stained by India ink. Scale bar: 200 μm.

The effects of NPW30 microinjected into the RVM (phase 1: p > 0.05, phase 2: p < 0.001). IP naloxone and IT WAY100135 significantly antagonized the effect of NPW30 and IT prazosin. IT idazoxan and IT SB269970 significantly enhanced the analgesic effect of NPW30. Ordinate: number of flinches per min; abscissa: time after drug administration (min).

The effects of NPW30 microinjected into the ipsilateral LC decreased the phase 1 (p < 0.005) and phase 2 (p < 0.001) responses. IP naloxone, IT idazoxan, IT WAY100135 antagonized the phase 2 depressant effect of NPW30. IT idazoxan significantly enhanced the phase 1 analgesic effect of NPW30 and antagonized the phase 2 effect. IT prazosin and IT SB269970 significantly enhanced the phase 2 analgesic effect of NPW30. Ordinate: number of flinches per min; abscissa: time after drug administration (min).

The effects of NPW30 microinjected into the contralateral LC decreased the phase 1 (p < 0.001) and phase 2 (p < 0.001) responses. IP naloxone and IT SB269970 significantly antagonized the phase 1 and phase 2 responses. IT prazosin and IT idazoxan antagonized the phase 2 effect of NPW30. Ordinate: number of flinches per min; abscissa: time after drug administration (min).
Effects of NPW30 injected into the RVM (Figure 2)
NPW30 treatment (n = 9) decreased the phase 2 flinching instances (p < 0.001), but not the phase 1 flinching instances (p > 0.05) as compared with the saline-treated rats (n = 10). The phase 2 depressant effect of NPW30 was significantly antagonized by IP naloxone (n = 8, p < 0.001) and IT WAY100135 (n = 5, p < 0.001). The phase 2 depressant effect of NPW30 was significantly enhanced by IT prazosin (n = 4, p < 0.001), IT idazoxan (n = 8, p < 0.01), and IT SB269970 (n = 5, p < 0.05).
Effects of NPW30 injected into the LC
Ipsilateral LC (Figure 3)
NPW30 treatment (n = 9) decreased phase 1 (p < 0.005) and phase 2 (p < 0.001) flinching instances as compared with the saline-treated rats (n = 7). The phase 2 depressant effect of NPW30 was significantly antagonized by IP naloxone (n = 8, p < 0.05). IT prazosin enhanced the phase 2 depressant effect of NPW30 (n = 4, p < 0.001). IT idazoxan (n = 8) significantly antagonized the phase 2 depressant effect of NPW30 (p < 0.01) and enhanced the phase 1 depressant effect (p < 0.05). The IT WAY100135 significantly antagonized the phase 2 (n = 5, p < 0.05) depressant effect of NPW30. IT SB269970 significantly enhanced the phase 2 (n = 5, p < 0.05) depressant effect of NPW30.
Contralateral LC (Figure 4)
NPW30 treatment (n = 12) decreased phase 1 (p < 0.001) and phase 2 (p < 0.001) flinching instances as compared with the saline-treated rats (n = 10).The phase 1 depressant effect of NPW30 was significantly antagonized by IP naloxone (n = 11, p < 0.005) and IT SB269970 (n = 5; p < 0.05). The phase 2 depressant effect of NPW30 was significantly antagonized by IP naloxone (n = 11, p < 0.001), IT prazosin (n = 4, p < 0.001), IT idazoxan (n = 17, p < 0.001), and IT SB269970 (p < 0.001).
Effects of NPW30 injected into the PAG
Ipsilateral PAG (Figure 5)
NPW30 treatment (n = 7) decreased phase 1 (p < 0.05) and phase 2 (p < 0.001) flinching instances as compared with the saline-treated rats (n = 10).The phase 1 depressant effect of NPW30 was significantly antagonized by IT WAY100135 (n = 5, p < 0.05). The phase 2 depressant effect of NPW30 was significantly antagonized by IP naloxone (n = 8, p < 0.001), IT idazoxan (n = 8, p < 0.001), IT WAY100135 (p < 0.001), and IT SB269970 (n = 5. p < 0.001).
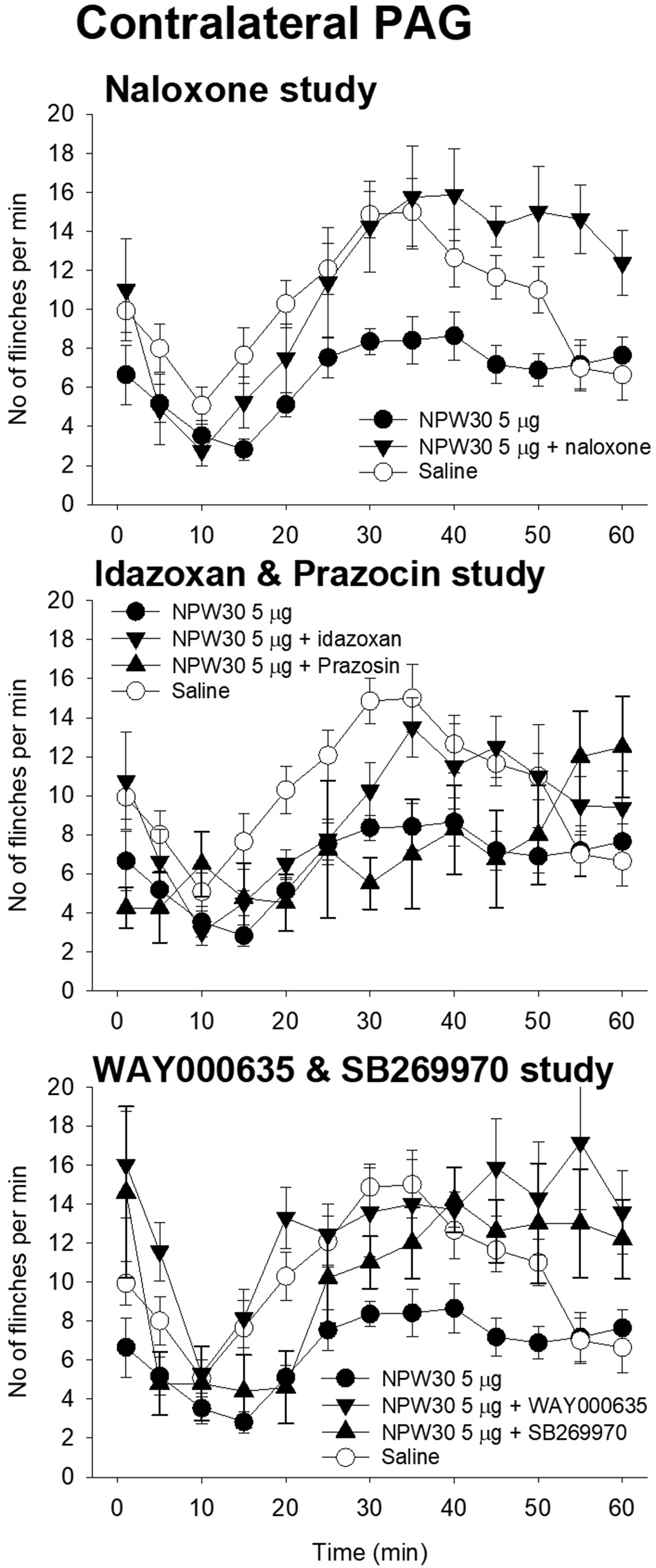
Contralateral PAG (Figure 6)
NPW30 treatment (n = 17) decreased phase 1 (p < 0.05) and phase 2 (p < 0.001) flinching instances as compared with the saline-treated rats (n = 14). The phase 1 depressant effect of NPW30 was significantly antagonized by IT WAY100135 (n = 7, p < 0.05). The phase 2 depressant effect of NPW30 was significantly antagonized by IP naloxone (n = 8, p < 0.001), IT idazoxan (n = 8, p < 0.001), IT WAY100135 (p < 0.001), and IT SB269970 (n = 5, p < 0.001).
Discussion
In the present study, NPW30 was microinjected into the RVM, ipsilateral LC, contralateral LC, ipsilateral PAG, and contralateral PAG. NPW30 was found to decrease the instances of flinching behavior induced by paw formalin injection. This suggests that NPW30 injected into the RVM, ipsilateral LC, contralateral LC, ipsilateral PAG, and contralateral PAG produced an analgesic effect on inflammatory pain. All these analgesic effects of NPW30 were antagonized by IP naloxone, suggesting that the NPW30 induced analgesic effect was, at least partly, mediated by opioidergic mechanisms. On the other hand, IT prazosin, an α-1 adrenergic receptor antagonist, and/or IT idazoxan, an α-2 adrenergic receptor antagonist, sometimes antagonized NPW30 induced analgesia and, at other times, enhanced NPW30 induced analgesia or had no effect on NPW30 induced analgesia. These findings suggest that when NPW30 is injected into the RVM, ipsilateral LC, contralateral LC, ipsilateral PAG, and contralateral PAG, activation of a spinal α-1 adrenergic receptor and spinal α-2 adrenergic receptor occurs, producing either a pronociceptive effect or anti-nociceptive effect. In some cases, NPW30 did not activate a spinal α-1 adrenergic receptor or spinal α-2 adrenergic receptor. The effects of 5-HT antagonists on NPW30 induced analgesia were almost the same as those of the α-1 and α-2 antagonists. IT WAY100135, a 5-HT1A receptor antagonist, and IT SB269970, a 5-HT7 receptor antagonist, also sometimes antagonized NPW30 induced analgesia and, at other times, enhanced NPW30 induced analgesia or had no effect on NPW30 induced analgesia.
Doses of drugs and timing of injection
As described in the Methods section, the doses of NPW30, antagonists, and injection timing were determined according to preliminary studies or previous reports. We found that IP naloxone, IT prazosin, IT idazoxan, IT WAY100135, and IT SB269970 at the doses used in this study antagonized the effect of NPW30 on phase 1 and/or phase 2 responses. These data suggest that the injected doses and the injection timing in this study were adequate to examine the analgesic mechanisms of NPW30.
Injection site in RVM, LC, and PAG
As shown in Figure 1, NPW30 was injected into the intended site in RVM, LC, and PAG. The PAG injection site is within the lateral part of the PAG. It has been reported that predominant Fos expression evoked by thermal noxious stimulation was observed in the lateral PAG. 18 Neurons in the dorsolateral/lateral columns of the PAG mediate active coping strategies that include anti-nociception associated with sympathoexcitation. 19 Thus, we believe that lateral PAG was involved in nociceptive modulation and that our PAG injection site was adequate to investigate the role of NPW30 in the PAG.
Role of opioidergic mechanisms on spinal nociceptive transmission
It has been suggested that gamma-aminobutyric acid (GABA) disinhibition in the RVM stimulates the opioidergic descending analgesic system. 20 In the RVM, microinjection of NPW30 may disinhibit the GABAergic system and activate opioid systems.
It has been reported that LC-mediated descending noradrenergic modulation is implicated in the analgesic mechanisms of opioids.21–23 Thus, it is possible that modulation of LC activity by NPW30 may activate the opioidergic analgesic system.
It has been reported that morphine microinjection into the PAG produces an analgesic effect and that this analgesic effect was antagonized by IT α–2 antagonist. 24 Moreover, PAG electrical stimulation-induced inhibition of nociceptive dorsal horn neurons in rats is associated with the release of noradrenaline. 25 These findings suggest that NPW30 microinjected into both the ipsilateral and contralateral PAG activated the opioidergic system in the PAG and increased spinal noradrenaline release. In this study, the analgesic effect of NPW30 microinjected into the ipsilateral and contralateral PAG was antagonized by both IP naloxone and IT idazoxan.
Role of noradrenaline receptor-mediated mechanisms on spinal nociceptive transmission
Noradrenaline has been reported to play an important role in the descending pain inhibitory system. The source of noradrenaline was LC, and noradrenaline was released in the spinal cord. Released noradrenaline in the spinal cord acts on α–2 receptors on the neuron to produce an analgesic effect. 26 It has been reported that the activation of α–2 receptors is generally associated with a suppression of neuronal activity in the dorsal horn. 27 On the other hand, activation of α–1 receptors exerted a robust excitatory influence upon the neuronal activity, and the α–1 receptor in the dorsal horn has been reported to have both pronociceptive and anti-nociceptive roles. 27
During phase 1, IT prazosin did not affect NPW30 induced analgesia. IT idazoxan enhanced NPW30-induced analgesia in ipsilateral LC studies, but not in other studies. This suggests that during phase 1, NPW30 microinjection into the ipsilateral LC induced noradrenaline release and produced a pronociceptive effect.
During phase 2, IT idazoxan antagonized the NPW30-induced analgesia in the ipsilateral LC, contralateral LC, ipsilateral PAG, and contralateral PAG studies and enhanced in the RVM study. IT prazosin antagonized NPW30 induced analgesia in the contralateral LC study and enhanced NPW30 induced analgesia in the RVM and ipsilateral LC studies. Thus, during phase 2, noradrenaline activated both the α-1 and α–2 receptors and produced pronociceptive and anti-nociceptive effects. It has been reported that when noradrenaline acts on the spinal astrocytes in superficial laminae, noradrenaline produces nociceptive effects by activating the α–1 receptor. 28 The mechanisms of how α–2 receptor activation produces a pronociceptive effect is not yet fully understood. However, our data strongly suggests that spinal noradrenaline produces a pronociceptive effect depending on the situation.
Role of 5-HT receptor-mediated mechanisms on spinal nociceptive transmission
5-HT has been known to have complex pain modulatory roles in the spinal cord, such as pronociceptive and anti-nociceptive effects.27,29 For example, low doses of IT 5-HT were found to produce an analgesic effect, whereas larger doses of IT 5-HT were found to induce a pronociceptive effect. 30 Moreover, either a 5-HT1A receptor agonist or 5-HT7 receptor agonist produced both pronociceptive and anti-nociceptive effects.29,31 In the present study, WAY100135, a 5-HT1A receptor antagonist, and SB269970, a 5-HT7 receptor antagonist, were used to clarify the role of the 5-HT receptor-mediated mechanism on NPW30 induced analgesia.
5-HT input to the spinal cord arose from the nucleus raphe magnus (NRM), and NRM was one of the nuclei composed of RVM. 27 In the RVM study during phase 2, WAY100135 antagonized the analgesic effect of NPW30 and SB269970 enhanced the analgesic effect of NPW30. These data strongly suggest that, in the RVM study, 5-HT1A receptor activation induced an anti-nociceptive effect, and 5-HT7 receptor activation produced a pronociceptive effect.
Electric stimulation of PAG and NRM had been reported to produce 5-HT mediated analgesia, and PAG was known to be another source of 5-HT. 27 In the ipsilateral and contralateral PAG study, either WAY100135 or SB269970 antagonized the analgesic effect of NPW30. These findings suggest that NPW30 microinjections into the PAG activated 5-HT1A and 5-HT7 receptors in the spinal cord, resulting in an analgesic effect.
Although the direct relationship between LC and spinal 5-HT was unclear, WAY100135 antagonized the analgesic effect of NPW30 in the ipsilateral LC study. SB269970 enhanced the analgesic effect of NPW30 in the ipsilateral LC study and antagonized the analgesic effect of NPW30 in the contralateral LC study. These data strongly suggest that NPW30 microinjected into the LC produced spinal 5-HT1A and 5-HT7 receptor-mediated nociceptive modulation.
Conclusion
Our findings strongly indicate that NPBW1/NPBW2 and the NPW system are involved in the descending pain modulatory system. NPW30 microinjection into the RVM, LC, and PAG modulated noradrenaline and 5-HT mediated nociceptive transmission in the spinal cord and produced a pronociceptive and anti-nociceptive effect depending on the situation.
The effects of NPW30 microinjected into the ipsilateral PAG decreased the phase 1 (p < 0.05) and phase 2 (p < 0.001) responses. IT WAY100135 significantly antagonized the phase 1 and phase 2 responses. IP naloxone, IT idazoxan, and IT SB269970 significantly antagonized the phase 2 depressant effect. Ordinate: number of flinches per min; abscissa: time after drug administration (min). The effects of NPW30 microinjected into the contralateral PAG decreased the phase 1 (p < 0.05) and phase 2 (p < 0.001) responses. IT WAY100135 significantly antagonized the phase 1 and phase 2 responses. IP naloxone, IT idazoxan, and IT SB269970 significantly antagonized the phase 2 depressant effect. Ordinate: number of flinches per min; abscissa: time after drug administration (min).
Footnotes
Authors Contribution
Shingo Nakamura: Designed the experimental protocol. Acquired and analyzed the data.
Takahiro Nonaka: Acquired and analyzed the data.
Koji Yoshida: Acquired and analyzed the data.
Toshihiko Yamada: Acquired and analyzed the data.
Tatsuo Yamamoto: Designed the experimental protocol and analyzed the data. Drafted the manuscript.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The authors were supported by a Grant-in-Aid for Scientific Research (C) and Kumamoto University.