
Abstract
Substance P (SP) and calcitonin gene-related peptide (CGRP) have both been considered potential drug candidates in migraine therapy. In recent years, CGRP receptor inhibition has been established as an effective treatment, in particular as a prophylactic for chronic migraine. Curiously, inhibition of neurokinin receptor 1 (NK1R) failed to alleviate acute migraine attacks in clinical trials, and the neurokinins were consequently abandoned as potential antimigraine candidates. The reason behind this has remained enigmatic.
Utilizing immunohistochemistry and semi-quantitative cell counts the expression of neurokinins and their associated receptors was examined in the rat trigeminal ganglion.
Immunohistochemistry results revealed SP co-localization in CGRP positive neurons and C-fibres, where it mainly concentrated at boutons. Neurokinin A (NKA) was observed in a population of C-fibres and small neurons where it could co-localize with SP. In contrast, neurokinin B (NKB) did not co-localize with SP and was observed in large/medium sized neurons and Aδ-fibres. All neurokinin receptors (NK1-3R) were found to be expressed in a majority of trigeminal ganglion neurons and A-fibres.
The functional release of SP and CGRP in the trigeminovascular system was stimulated with either 60 mM K+ or 100 nM capsaicin and measured with an enzyme-linked immunosorbent assay (ELISA). ELISA results established that SP can be released locally from trigeminovascular system. The released SP was comparatively minor compared to the CGRP release from stimulated dura mater, trigeminal ganglion neurons and fibres. We hypothesize that SP and CGRP signalling pathways may work in tandem to exacerbate painful stimuli in the TGV system.
Introduction
Migraine is the most frequent neurological disorder, characterized by moderate to severe headaches lasting for up to 72 h and affecting 1 billion people globally. 1 The global burden of diseases (GBD) study revealed migraine as the second worldwide cause of years lived with disability (YLDs). 2 The origin of a migraine attack has long been debated but is currently suggested to emanate from the CNS, 3 while migraine-pain triggering mechanism relies on the relationship between the trigeminovascular (TGV) system and brainstem nuclei. 4 The trigeminal ganglion (TG) has long been suggested to have an important role in migraine pathophysiology as it contains the primary sensory neurons innervating large parts of the skull and facial area.5,6
The development of immunohistochemistry (IHC) and specific neuropeptide antibodies have provided specific tools to examine the sensory nerves in more detail. One of the first sensory neuropeptide candidates to come into focus was the neurokinin substance P (SP).7,8 This peptide was soon a prime candidate for migraine pharmacotherapy because of its involvement as a mediator in neurogenic inflammation (NI). SP has vasodilatory properties, 7 can excite nociceptive neurons, 9 may activate the trigeminal pathway and cause mast cell degranulation and extravasation of plasma proteins.10,11 A few years later it was discovered that calcitonin gene-related peptide (CGRP) was co-stored with SP, was a stronger and more potent vasodilator, acted differently and was released in primary headache attacks.12–15 Because CGRP had no effect on NI, did not excite the trigeminal afferents and did not induce plasma protein extravasation, it was therefore considered inconsequential to migraine. 16 We now know that CGRP is a key molecule in migraine therapy and blocking it or its receptor can reduce monthly migraine attacks drastically. 17
Erbersberger studied in detail the relative roles of SP, CGRP and prostaglandin E2 on their release after antidromic electrical stimulation and orthodromic chemical stimulation. 18 Amazingly, SP showed no release after the stimulation protocols and hence its role in NI with plasma protein extravasation in the rodent dura mater was diminished. Studies have revealed that blockade of CGRP signalling has both acute and prophylactic effect on human migraine17,19 while clinical trials blocking neurokinin receptors showed no effect on migraine.20–22
Neurokinin A and B (NKA, NKB) are SP-alike neuropeptides and belong to the same family of tachykinins (neurokinins). 23 While NKB is encoded by the TAC3 gene, NKA and SP are encoded by the same gene (TAC1) and derived from the same preprotachykinin. These neuropeptides (SP, NKA and NKB) are all agonists for the neurokinin 1, 2 and 3 receptors (NK1R, NK2R and NK3R) but with varying affinity. SP has a high affinity for NK1R, while NKA and NKB have high affinity for NK2R and NK3R, respectively.
In addition to antiemetic properties, NK1R antagonism has been shown to disrupt itch signalling and reduce itching.24,25 Thus, SP and NK1R have been implicated as potential targets to treat chronic pruritus accompanying multiple conditions. 26
The development of novel tools for IHC prompted us to revisit the neurokinins. The present study was designed to examine in detail the relation between neurokinins and CGRP mechanisms in the TGV system based on preliminary observation of SP and CGRP co-existence in C-fibre boutons. To gain a deeper understanding of putative sites of interaction between the different sensory neuropeptides of the TGV system IHC, semi-quantitative cell counting and ELISA for measuring the amount of CGRP and SP released from different parts of the TGV system (dura mater, soma poor and soma rich parts of the TG) was utilized in this study.
Methods
Animals
All animal procedures were performed in accordance with the European Community Council Directive on ‘The Protection of Animals Used for Scientific Purposes’ (2010/63/EU). Animal procedures were approved by the Lund University Animal Ethics Committee (M43–07) and The Danish Animal Experimentation Inspectorate.
Adult male Wistar rats (n = 8, 260–300 g), housed in groups of 2–3 rats together in Tall IVC Rat Cages (Innovive), were used for the IHC part of the study. For CGRP and SP release experiments additional rats (n = 6, 280–320 g), housed in Euro standard cages (Type VI with 123-Lid) in groups of six rats together, were used.
All animals were kept under standard laboratory conditions in a temperature and humidity-controlled environment with a 12/12 h light-dark cycle, with dark beginning at 7 p.m. The animals had access to water and chow (RM1, SDS) ad libitum. The animals were anaesthetized by CO2 inhalation and subsequently decapitated prior to experiments.
Immunohistochemistry
TGs were carefully dissected and incubated in 4% paraformaldehyde in phosphate buffered saline (PBS) for a duration of 2–4 h in +4°C. The fixated TGs were immersed in 10% sucrose (Sigma) in Sorensen’s phosphate buffer (2–4 h, +4°C) and subsequently immersed in a 25% solution overnight. The following day, the TGs were embedded in a gelatin medium (30% egg albumin, 3% gelatin), frozen (−20°C) and cryosectioned at 10 μm. The sections were mounted on microscope slides (Superfrost, ThermoFisher) and stored at −20°C until use.
Details of primary and secondary antibodies.

Cell counting was performed for all TG sections labelled with SP in combination with CGRP, NKA and NKB (n = 8, sections = 24 i.e. 3x8). Images were obtained of TG neuron clusters in an area of 0.1875 mm2 using a ×20 magnification lens. The chosen exposure time for each filter was 1s (TRITC, CGRP) and 333 ms (FITC, SP). Both negative and immunoreactive neurons were counted in each image. Resulting data was compiled into a diagram using GraphPad Prism 9.1.2 (GraphPad Software, CA, USA).
Release of SP and CGRP from the TGV system
The skull was cut mid-sagittally and the brain halves were carefully removed while the cranial dura was left attached to the skull. Thereafter the TGs were carefully dissected out. For the buffer system, 300 μL of synthetic interstitial fluid (SIF, composition: 108 mM NaCl, 3.5 mM KCl, 3.5 mM MgSO4, 26 mM NaHCO3, 11.7 mM NaH2PO4, 1.5 mM CaCl2, 9.6 mM Sodium Gluconate, 5.6 mM glucose and 7.6 mM sucrose; pH 7.4) at +37°C was used. Each TG was carefully dissected into a neuronal soma rich and a neuronal soma poor portion, in accordance to previous studies. 27
TG samples were randomized, placed in Eppendorf tubes in a heating block at +37°C and washed. For the skull halves, these were also randomized and placed in a humid chamber above a water bath to maintain temperature at +37°C. The release of CGRP and SP was induced by 60 mM potassium or 100 nM of the TRPV1 agonist capsaicin. To maintain equal osmolality in the 60 mM K+ SIF buffer, NaCl had been exchanged for KCl on an equimolar basis. 200 μL samples for measuring CGRP/SP content were collected from both tissues 10 min after stimuli, mixed with 50 μL enzyme immunoassay buffer (containing protease inhibitors) and stored at −20°C until analysis, performed maximally a week after the release experiment was performed. See Supplementary Figure 1 for an outline of the experimental layout.
The samples, 100 μL for CGRP and 100 μL for SP, were processed using commercial EIA kits. The human CGRP ELISA KIT (SPIbio, Paris, France) was used to study CGRP release with a limit of detection of 0.7 pg/mL and a specificity for rat CGRP-α/β at 120%. For SP release, the rat SP ELISA Kit (Enzo, ADI-900-018, NY, USA) with detection range: 9.76–10.000 pg/mL and sensitivity: 8.04 pg/mL was used. Determination of CGRP and SP contents were calculated based a standard curve that was run in duplicates. The protocol was performed following the manufacturer’s instructions and the optical density was measured at 410 nm using a micro-plate photometer (Tecan, Infinite M200, software SW Magellan v.6.3, Männedorf, Switzerland).
Statistics
Statistical calculations were performed using GraphPad Prism. The data is presented as mean ± SEM. Number of experiments = n. The data was analysed with paired Student’s T-test. p < 0.05 was considered significant. All the CGRP and Substance P release data passed the Shapiro-Wilk normality test (alpha = 0.05).
Results
Immunohistochemistry
The immunoreactivity (ir) for SP was expressed in small to medium sized neurons (an approximate diameter of <30–60 μm) and C-fibres (Figure 1). In neurons, the SP ir was observed as a granular pattern in the cytoplasm and was rarely seen in the nuclei. SP ir in C-fibres was often observed to concentrate in C-fibre boutons, however SP ir could sometimes also be observed in full lengths of C-fibre axons. No SP ir was observed in Aδ-fibres, Schwann cells or satellite glial cells (SGCs). Localization of substance P (SP) in relation to calcitonin gene-related peptide (CGRP), neurokinin A (NKA) and neurokinin B (NKB) in the rat trigeminal ganglion. (a) CGRP was expressed in small to medium sized neurons and was regularly detected in whole lengths of C-fibre type axons, thus revealing boutons. Similarly, SP was robustly expressed and co-localized in the same neurons and C-fibres, albeit fewer neurons expressed SP immunoreacitivity (ir) and SP was generally concentrated to boutons in C-fibres. SP ir could also be found in non-bouton axonal structures, such as close to the axon hillock (arrow). Insert: C-fibre axon expressing CGRP co-localizing with SP in boutons (arrow). (b) NKA ir was prominently expressed in C-fibres (arrow) and a majority of small to medium sized neurons with a notable expression around the negative nucleus (arrowhead). Co-staining with SP revealed co-localization in most of these structures. No NKA ir was observed in satellite glial cells (SGCs) and Schwann cells. Insert: NKA immunoreactive C-fibre displaying three SP immunoreactive boutons (arrow). (c) Contrastingly, NKB ir was observed mainly in larger neurons and Aδ-fibres (arrowhead). NKB was not observed in SGCs, Schwann cells or C-fibres. In general, SP did not co-localize with NKB in neither neurons nor C-fibres (arrowhead). Insert: A large neuron distinctly expressing NKB ir (arrowhead), the wavy pattern could be interpreted as ir of the endoplasmic reticulum. Below, a typical SP immunoreactive neuron (arrow) is presented.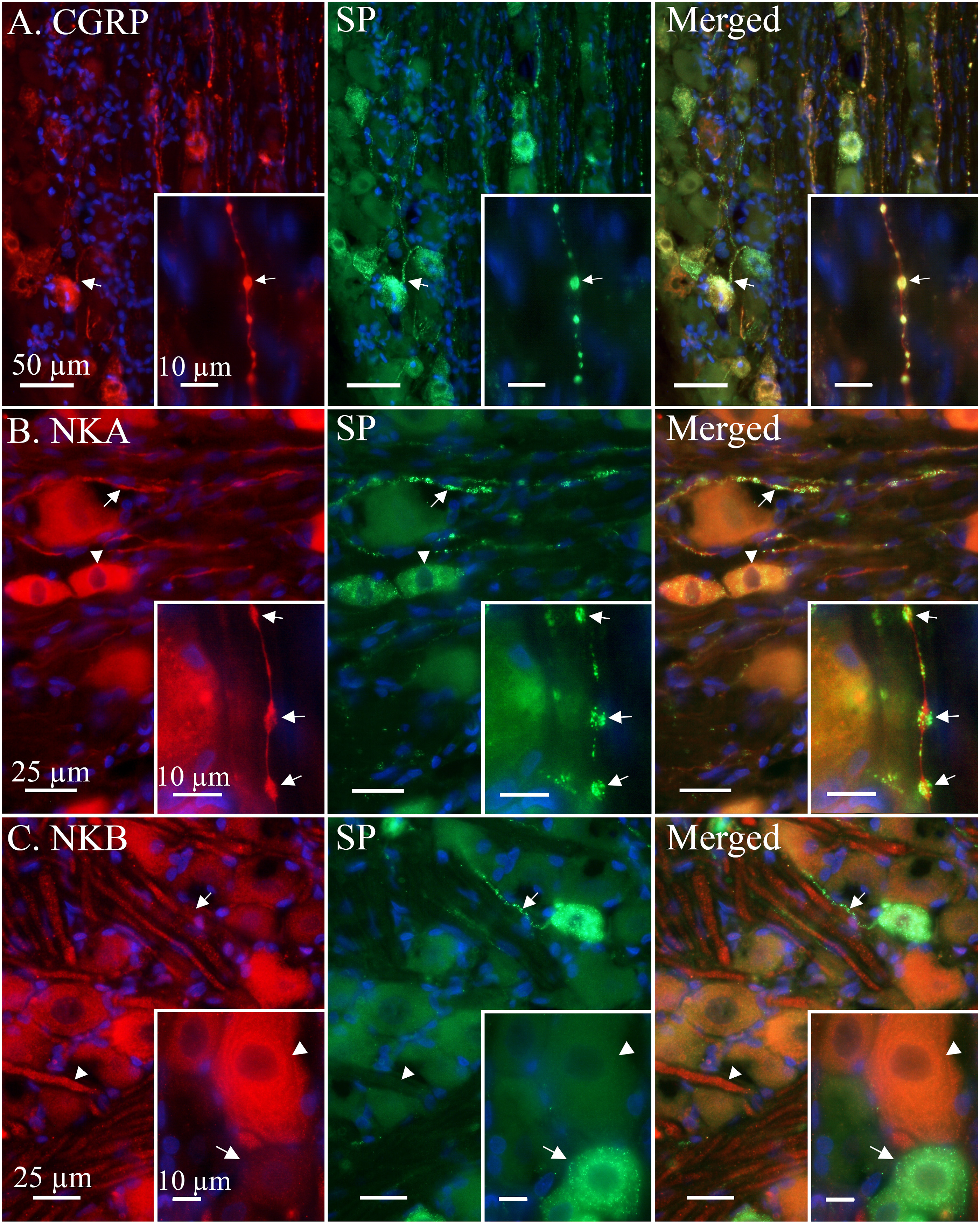
To confirm the SP C-fibre expression double IHC with CGRP was performed (Figure 1(a)). CGRP ir co-localized with SP in both neurons and C-fibres. CGRP ir was not observed in Aδ-fibres, Schwann cells or SGCs. CGRP was also observed in several neurons and C-fibres not expressing SP. Similarly, a few neurons and C-fibres were found to only be immunoreactive for SP and not CGRP. However, CGRP ir was more frequently expressed in the TG C-fibres and neurons than SP.
NKA ir was only expressed in small to medium sized neurons and C-fibres (Figure 1(b)). In these neurons, the ir was strong around the NKA-negative nuclei forming a ring-like structure likely corresponding to the endoplasmic reticulum. The NKA-positive C-fibres were numerous and easily detected but had a weaker staining than both SP and CGRP. Like SP and CGRP, no ir for NKA was detected in Aδ-fibres, Schwann cells or SGCs.
Contrastingly, NKB ir was mainly expressed in medium to large neurons (an approximate diameter >30 μm) and Aδ-fibres (Figure 1(c)). No NKB ir was observed in satellite glial cells (SGCs), Schwann cells or neuron nuclei. Similarly to NKA, in some large neurons organelle structures resembling the endoplasmic reticulum could be seen. Interestingly, NKB did not co-localize with SP in either neurons or fibres.
NK1R was prominently expressed in Aδ-fibres, most neurons and their nuclei (Figure 2(a)). Several instances of NK1R ir in SGCs and Schwann cells were also observed. In addition, NK1R ir was observed in C-fibres co-localizing with SP, suggesting the presence of autoreceptors. Localization of neurokinin receptor 1 (NK1R), neurokinin receptor 2 (NK2R), neurokinin receptor 3 (NK3R) and contactin-associated protein 1 (CASPR) in relation to substance P (SP) in the rat trigeminal ganglion. (a) NK1R displayed robust immunoreactivity (ir) in the majority of observed neurons, neuron nuclei, Schwann cells and A-fibres (Asterix). The NK1R expression in SGCs and C-fibres was slightly weaker. NK1R was observed to co-localize with SP in C-fibres (arrow) and small to medium sized neurons (arrowhead). Insert: Three neurons displaying NK1R ir with one also expressing SP ir (arrowhead). Close-by a SP and NK1R immunoreactive C-fibre (arrow) can be observed. (b) NK2R was expressed in most observed neurons, neuron nuclei (arrow) and A-fibres. The expression in A-fibres was patchy and weaker than that of NK1R. NK2R ir co-localized with SP in neurons (arrowhead) but was not readily distinguished in associated C-fibres. Insert: An unevenly stained NK2R immunoreactive Aδ-fibre (arrowhead) near a SP immunoreactive C-fibre (arrow). (c) In general, NK3R ir resembled NK1R ir, though weaker and without a clearly visible nuclear envelope. NK3R ir was observed in most neurons (arrowhead), A-fibres, SGCs and Schwann cells. When co-stained with SP a weak co-localization could be observed in C-fibres (arrow). Insert: Co-localization of NK3R and SP in a medium sized neuron (arrowhead). A weakly stained C-fibre (arrow) can be seen projecting from its associated neuron. (d) The nodes of Ranvier were robustly revealed utilizing the specific CASPR antibody. Double immunohistochemistry with SP displayed several SP positive C-fibre boutons aligning with the node of Ranvier (arrow). Insert: A close-up on the CASPR positive paranodal regions of the Aδ-fibre axon flanking the node of Ranvier (arrowhead). A C-fibre bouton displaying SP ir (arrow) can be located nearby. The visible red ir in the C-fibre bouton is likely due to an unspecific binding of the polyclonal CASPR primary antibody.
NK2R was most strikingly expressed in neuron nuclei but was also observed in neuron cytoplasm and Aδ-fibres in a granular and patchy fashion (Figure 2(b)).
Most neuronal soma, A-fibres, SGCs and Schwann cells were observed as immunoreactive to NK3R (Figure 2(c)). NK3R resembled NK1R, though weaker and less specific (no clear borders between Schwann cells and A-fibres were observed). Weakly stained C-fibre axons could be observed when co-stained with SP. NK1R and NK2R and NK3R could all be observed to co-localize with SP in neuron cell bodies.
IHC for contactin-associated protein 1 (CASPR) was performed to evaluate if SP ir C-fibre boutons could align with Aδ-fibre nodes of Ranvier as previously reported for CGRP. 27 CASPR ir revealed the paranodal and juxtaparanodal regions of myelinated A-fibres in the TG sections (Figure 2(d)). The majority of observed myelinated axons ranged between 1-5 μm in diameter, which morphologically classifies them as Aδ-fibres. The SP immunoreactive C-fibre boutons could often be found to align with adjacent nodes of Ranvier on CASPR positive Aδ-fibres.
Cell counting
The cell counting of immunohistochemically stained TG sections revealed that 27.3% of neurons were immunoreactive for CGRP and 13.1% for SP (Figure 3(a)). Of these, 15.4% were only positive for CGRP and 1.2% were only positive for SP. 11.9% of the counted cells were positive for both CGRP and SP. In total, 478 neurons were counted and 71.4% of the neurons were considered negative for both CGRP and SP. Cell counting of immunohistochemical sections. (a) 27.3% of the counted neurons were immunoreactive to CGRP and 13.1% for SP. Of these, 15.4% were only positive for CGRP and 1.2% were only positive for SP. 11.9% of the counted cells were positive for both CGRP and SP. In total, 478 neurons were counted and 71.4% of neurons did not exert ir for CGRP or SP. (b) 24.0% of counted neurons displayed ir for NKA and 14.0% for SP. Of these, 12.1% were only positive for NKA and 2.2% were only positive for SP. 11.9% of the counted cells were positive for both NKA and SP. In total, 603 neurons were counted and 73.8% of neurons did not exert ir for NKA or SP. (c) 21.5% of the counted neurons were immunoreactive to NKB and 14.5% for SP. Of these, 20.8% were only positive for NKB and 13.8% were only positive for SP. 0.7% of the counted cells were positive for both NKB and SP. In total, 607 neurons were counted and 64.7% of neurons did not exert ir for NKB or SP.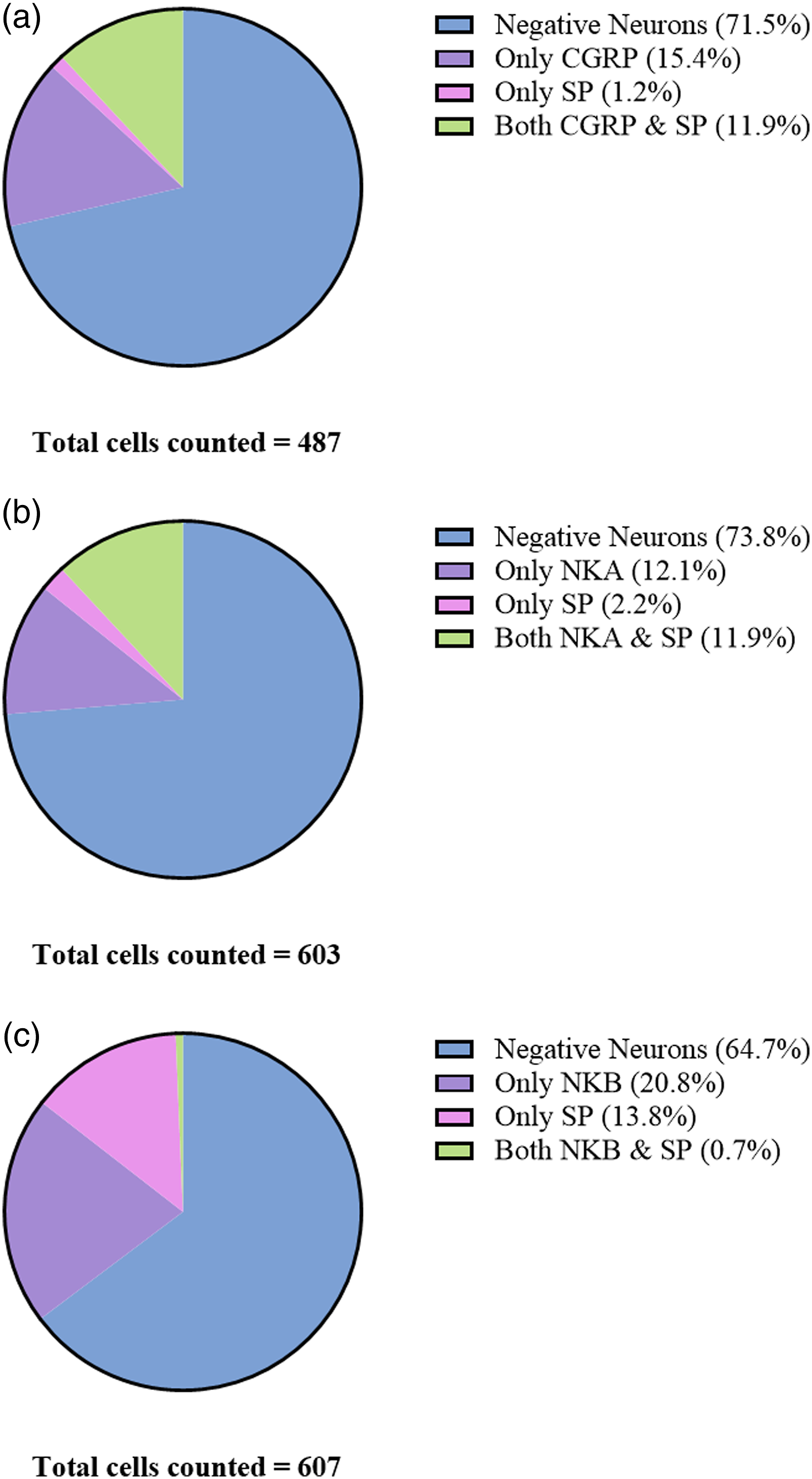
Similarly, 24.0% and 14.0% of counted neurons expressed ir for NKA and SP, respectively (Figure 3(b)). 12.1% displayed ir for NKA exclusively while 2.2% only were immunoreactive for SP. Identical to the CGRP results, NKA and SP co-localized in 11.9% of the neurons. In these samples, 603 neurons were counted in total and 73.8% of the neurons were considered negative. Lastly, neurons stained with NKB and SP were counted (Figure 3(c)). Of these, 21.5% were immunoreactive to NKB and 14.5% for SP. Contrastingly to the previous counts, 20.8% were only positive for NKB and 13.8% expressed SP. In only 0.7% of the counted neurons was co-localization of NKB and SP observed. In total, 607 neurons were counted and 64.7% of neurons did not exert ir for either NKB or SP.
Release of substance P and CGRP from the TGV system
Commercially available ELISA kits were utilized for detecting SP and CGRP from the same sample. This study focused on the peripheral parts of the TGV system including the dura mater, the soma-poor and the soma-rich regions of the TG. When stimulating the samples with 60 mM K+ we observed a minor, but significant, increase in the SP level (from 16.4 ± 0.8 pg/mL to 19.3 ± 0.7 pg/mL, p = 0.006) from the dura mater but not from the soma-free nor the soma-rich part of the TG (Figure 3(a)). This suggests that SP release by depolarization in the TGV system, only occurs in the dura mater. We compared the data to CGRP levels from the same samples. When applying 60 mM K+ CGRP was significantly increased in the dura (43.8 ± 3.9 pg/mL, p = 0.009), the soma-poor TG (40.9 ± 2.8 pg/mL, p = 0.042) and the soma-rich TG (49.8 ± 6.9 pg/mL, p = 0.028). It is evident from the individual data points (Figure 3(b)) that only the dura had a consistent increase in the SP release.
Since CGRP is well known to be released after stimuli with the transient receptor potential vanilloid 1 (TRPV1) agonist capsaicin, we wanted to examine if the results were similar as for the depolarization. This time there was no release of SP from the dura mater (Figure 3(c)). In contrast, we observed significant SP release when stimulating the soma-poor part of the TG (from 24.6 ± 1.1 pg/mL to 30.3 ± 0.8 pg/mL, p = 0.007). We did not observe SP release from the soma-rich part of the TG (from 22.4 ± 1.8 pg/mL to 19.6 ± 1.3 pg/mL, p = 0.5). For CGRP, the release was high and consistent (p < 0.01) in all samples. All individual samples for the soma-poor TG showed an increase in SP levels (Figure 3(d)).
Discussion
While we have previously reported the trigeminal localization of CGRP and the CGRP receptor,27,28 we now asked if the neurokinin system might share similarities with the CGRP system. Our key aim was to delve deeper in understanding the putative sites of interaction between the different sensory nerve fibres and the mechanism of action related to modern anti-migraine drugs (e.g. CGRP-receptor antagonists) within the trigeminal system. We have with a set of novel antibodies shown the localization of the neuropeptides SP, NKA and NKB, as well as of their three receptors NK1-3R in the trigeminal system. Stimulated release experiments with potassium induced depolarization or capsaicin revealed minor SP release as compared to the co-release of CGRP below we discuss their relation to migraine headache.
There exist three neurokinin receptors, but the vasoactive properties of the neurokinins are basically mediated via the NK1R. The NK1R is found throughout the peripheral and central nervous systems. 29 Early nonapeptide NK1R antagonists, such as RPR100893, were found to block plasma protein extravasation within the dura mater following electrical stimulation of the trigeminal ganglion 30 and to inhibit the c-fos expression in the trigeminal nucleus caudalis (TNC) in response to the C-fibre stimulant capsaicin. 31 These and other similar results lead to the hypothesis that NK1R antagonists would be beneficial in the treatment of migraine. However, clinical studies with RPR100893 32 and LY303870 33 were without effect.
In the current work, our IHC experiments show that NK1R was prominently expressed in Aδ-fibres, most neurons and their nuclei. In addition, NK1R was observed in SGCs, Schwann cells and C-fibres co-localizing with SP, which suggests autoreceptors. The expression of SP was about half of that of CGRP but still with a high level of co-localization in both neurons and C-fibres. Further, it was evident that NKA had a slightly lower number of positive cells compared to CGRP and was less robustly expressed in C-fibres, which may suggest a lower stimulated release. NKA may still have a role in migraine pathophysiology, though its preferred receptor NK2R had a weaker expression than NK1R and could thus be less relevant. Surprisingly, the NKA and NKB results suggest that the two neurokinins are expressed in two different populations of neurons within the TG. NKA was mainly expressed in small to medium sized neurons and C-fibres, while NKB was mainly observed in larger neurons and Aδ-fibres.
NKB was robustly expressed, but contrary to SP, seemed to be expressed in larger neurons and Aδ-fibres. This neuron population is likely the same as those expressing the CGRP receptor elements, receptor activity-modifying protein 1 (RAMP1) and calcitonin receptor-like receptor (CLR), and NKB is thus likely released from the neuron cell body and terminal synapses. In theory, NKB could be released from the nodal and paranodal regions of the axon though this seems inefficient as endocytic vesicles fusing with the membrane could disrupt the ion channel density of the nodes of Ranvier. Lastly, the NK3R ir was observed to be similar to, but weaker than, NK1R ir.
These morphological findings point towards a minor role of the neurokinins as compared to CGRP in the TGV system. We therefore turned to investigate the functional release of SP in comparison to CGRP, a well-established experimental technique used to examine trigeminal sensory processing.
34
In agreement with previous studies,
35
the release data suggests that mainly CGRP and not SP is released upon both K+ and capsaicin stimulation. Nevertheless, significant release of SP was observed in the dura mater when stimulating with potassium (Figure 4(a)) and in the soma-poor parts of TG when stimulating with capsaicin (Figure 4(b)). This raises the question whether different release mechanisms are dominant for the C-fibres inhabiting the dura mater and TG, respectively. CGRP/Substance P release. (a) The addition of 60 mM KCl (K+) caused SP release from the dura (n = 6) but not from the soma-poor or soma rich parts of TG. CGRP release was much stronger and significant from the dura, soma-poor and soma-rich TG. (b) Graphical illustration of the sample pairing. (c): The addition of 100 nM capsaicin caused significant SP release from the soma-poor part of TG, but not from the dura or soma-rich TG (n = 6), this contrasts to strong and significant CGRP release form the dura, soma-poor and soma-rich TG. (d) Graphical illustration of the sample pairing. Data are shown as mean ± SEM or their individual data points with pairing, and with *p > 0.05, **p > 0.01, ***p > 0.001 from the paired Student’s T-test being depicted in the graph.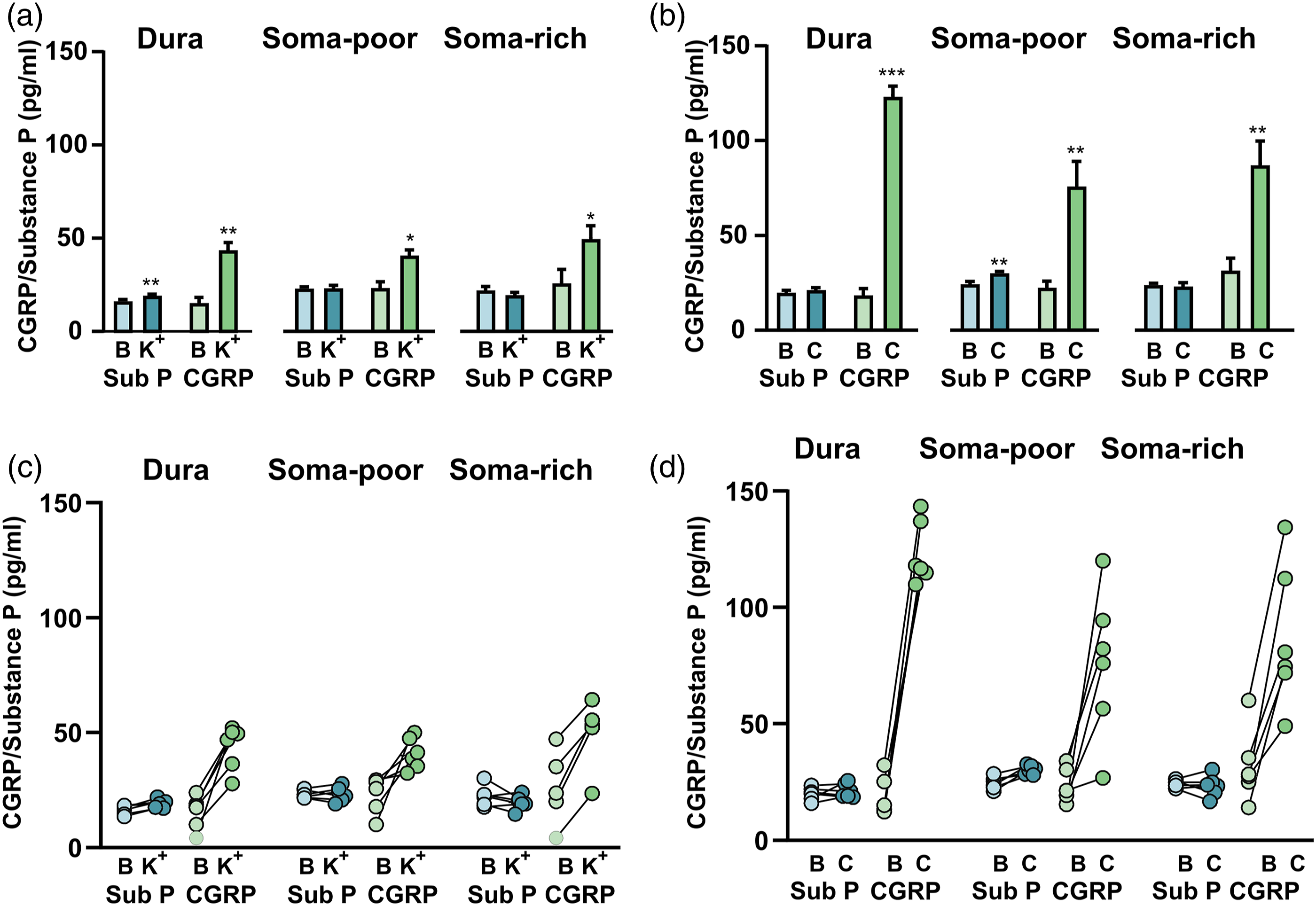
However, no significant SP release was observed from the soma-rich TG when stimulating with either capsaicin or potassium (Figure 3). This suggests that C-fibre boutons, and not the neuron cell bodies, are the principal release sites for SP in the peripheral trigeminal system. An explanation to the results could be the high expression of the NK1R in combination with the high-affinity reuptake process. 36 The presence of autoreceptors on C-fibres seems plausible as it has previously been suggested that SP binding to NK1 autoreceptors could exert a negative feedback on its own release. 37 This could explain why such a small amount of released SP was measured in our ELISA experiments. In addition, unbound SP is reported to have a short half-life in tissues (minutes), compared to plasma (hours), 38 likely due to the presence of p-endopeptidase in the extracellular fluid. 39 Future functional studies could provide insight on which mechanism is more dominant. Combined, these observations agree well with the absence of a significant release of SP in migraine and in cluster headache attacks.40,41
Combing the data from the present work does not exclude the role of the neurokinins and the NKRs, but suggest that they could be part of a more intricate system. Particularly, we believe the findings are interesting in the light of our new cross-talk hypothesis. 27 The use of the CASPR antibody revealed with great clarity the nodes of Ranvier on the unmyelinated part of Aδ-fibres in our previous study in CGRP. 27 The role of the nodes of Ranvier is the induction of saltatory conduction to speed up the transmission rate in the myelinated sensory nerve fibres.42,43 Positive expression of the CASPR antibody can easily be distinguished as a ‘bowtie-like’ pattern on myelinated fibres. This can be seen in Figure 2(d), where the CASPR ir localized to either side of the unmyelinated node of Ranvier on Aδ-fibres.
SP immunoreactive C-fibre boutons could be found to align with proximal nodes of Ranvier in the same manner as previously reported for CGRP. 27 The results suggest that SP, NKA and CGRP are co-localized in C-fibre boutons and thus could be co-released from presynaptic sites. This is supported by the release data where capsaicin triggers both SP and CGRP release from soma-poor areas (Figure 4(b)). Plausible targets for locally released SP and NKA could be proximal NK1R and NK2R expressed on neurons, Schwann cells and, importantly, Aδ-fibres via the node of Ranvier. The Aδ-fibre expression of NK1R, and to a lesser extent, NK2R, suggests the possibility of axon-axon signalling occurring in similarly to the CGRP system. 27 Interestingly, NKB ir was mainly found in larger neurons, resembling CGRP receptor component RAMP1 positive neurons, and Aδ-fibres. NKB was not robustly found to co-localize with SP in smaller to medium sized neurons or C-fibres which suggests a different release pattern for NKB. The NK3R in turn is expressed in C-fibres and CGRP/SP positive neurons; it could potentially be a viable drug target for the neurogenic inflammation hypothesis in migraine therapy. However, the release of NKB in the trigeminal system is likely limited to neuron cell bodies and terminal synapses of A-fibres. Although previous attempts at alleviating migraine pain with NK1R inhibitors have proven ineffective, neurokinin receptors may yet have a role to play in neuro-inflammatory pain signalling and chronification. 44 Notably, both CGRP and SP have been shown to degranulate dural mast cells 45 and NK1R activation has been shown to stimulate the production of cytokines via the MEK/ERK pathway. 38
As peripheral injections of CGRP alone does not induce pain, a significant painful sensation can however be induced in combination with SP or NKA. 46 This suggests that these co-expressed C-fibre neuropeptides may act on two separate pain-promoting systems and/or exacerbate sensory stimuli in the trigeminal system. Both CGRP and SP receptor signalling activates the cyclic AMP pathway leading to a plausible activation of voltage gated ion channels and an increased electrophysiological burst frequency. 47 This mechanism is likely more dependent on CGRP, due to its more prominent expression and release of CGRP, compared to SP. However, SP has been reported to inhibit inhibitory postsynaptic potentials making the propagation of painful stimuli more likely to occur in the axon. 47 This could, in theory, mean that locally released CGRP and SP work in tandem to lower action potential thresholds at the Aδ-fibre nodes of Ranvier and increase the probability for painful stimuli. We therefore hypothesize that CGRP receptor inhibition combined with NK1R or NK2R inhibition may reach a broader spectrum of migraine patients than would a monotherapy.
Supplemental Material
sj-tif-1-mpx-10.1177_17448069211059400 – Supplemental Material for Neurokinins and their receptors in the rat trigeminal system: Differential localization and release with implications for migraine pain
Supplemental Material, sj-tif-1-mpx-10.1177_17448069211059400 for Neurokinins and their receptors in the rat trigeminal system: Differential localization and release with implications for migraine pain by Jacob CA Edvinsson, Philip V Reducha, Majid Sheykhzade, Karin Warfvinge, Kristian A Haanes and Lars Edvinsson in Molecular Pain
Footnotes
Acknowledgments
Not applicable.
Abbreviations
AMP: adenosine monophosphate. BBB: blood–brain barrier. BSA:bbovine serum albumin. cAMP: cyclic adenosine monophosphate. CASPR: contactin-associated protein 1. CGRP: calcitonin gene-related peptide. CLR: calcitonin like receptor. EIA: enzyme immunoassay. ELISA: enzyme-linked immunosorbent assay. GBD: The global burden of diseases. IHC: immunohistochemistry. Ir: immunoreactivity. MAbs: monoclonal antibodies. NI: neurogenic inflammation. NKA: neurokinin A. NKB: neurokinin B. NK1R: neurokinin receptor 1. NK2R: neurokinin receptor 2. NK3R: neurokinin receptor 3. PBS: phosphate buffered saline. PBS-T: phosphate buffered saline with Triton X-100. PNS: peripheral nervous system. RAMP1: receptor activity modifying protein 1. SGCs: satellite glial cells. SP: substance P. SIF: synthetic interstitial fluid. TRPV1: transient receptor potential vanilloid 1. TG: trigeminal ganglion. TGV: trigeminovascular. TNC: trigeminal nucleus caudalis. YLDs: years lived with disability.
Author contributions
JE, LE and KAH designed the study. JE, PR and KAH performed the experiments. LE provided tools and reagents. JE, LE and KAH wrote the manuscript. MS and KW made further critical manuscript revisions. All authors read and approved the final manuscript.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: KAH was supported by a Lundbeck foundation Fellowship (R345-2020-1977). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Ethics approval and consent to participate
The experimental procedures for the IHC in Sweden were approved by the Lund University Animal Ethics Committee (M43–07). The CGRP release experiments performed in Denmark were approved by The Danish Animal Experimentation Inspectorate. All experiments were performed in accordance with the European Community Council Directive on ‘The Protection of Animals Used for Scientific Purposes’ (2010/63/EU).
Consent for publication
Not applicable.
Availability of data and materials
The datasets generated during and/or analysed during the current study are available from the corresponding author on reasonable request.
Supplementary material
Supplementary material for this article is available online.