
Abstract
Chronic pain has detrimental effects on one’s quality of life. However, its treatment options are very limited, and its underlying pathogenesis remains unclear. Recent research has suggested that fragile X mental retardation protein is involved in the development of chronic pain, making it a potential target for prevention and treatment. The current review of literature will examine the function of fragile X mental retardation protein and its associated pathways, through which we hope to gain insight into how fragile X mental retardation protein may contribute to nociceptive sensitization and chronic pain.
Introduction
Chronic pain, defined as “pain that persists or recurs for more than three months,” can have a wide range of etiologies, such as inflammation, nerve injury, cancer, post-trauma, and post-surgery. 1 It is a prevalent condition with an enormous cost to the society and profound socioeconomic impacts.2–4 On an individual level, chronic pain sufferers report poor quality of life, increased tendency for substance abuse, and a higher incidence of mental disorders, such as anxiety and depression. 5
Despite now being recognized as a major group of diseases by the World Health Organization,6,7 treatments for chronic pain are still limited. Pharmacological agents, such as opioids and anticonvulsants, are effective for temporary pain relief, but have many undesirable side effects and their efficacy diminishes with long-term use.8–10 While central nociceptive sensitization is considered as a major contributing factor to the development and the maintenance of chronic pain,11–13 the lack of detailed mechanistic understanding of how sensitization occurs has remained a major barrier for developing new pharmacological treatment.
Fragile X mental retardation protein (FMRP), encoded by fragile X mental retardation 1 (Fmr1) gene, has been implicated in pathogenesis of chronic pain. Mutation in Fmr1 gene is thought to lead to Fragile X syndromes (FXS), a spectrum of clinical conditions characterized by self-injurious behavior and abnormal pain processing.14–16 Fmr1 knock-out (KO) mice show decreased nociceptive behaviors, which suggests FMRP playing a vital role in the pathogenesis of abnormal pain states. 14
In this review, we will examine the molecular function of FMRP and its role in the development of chronic pain.
The molecular functions of FMRP
FMRP has many functions in the nervous system with a complex protein architecture (Figure 1). 17 Specifically, the K homology (KH) domains and glycinearginine box (RGG) of FMRP are protein-binding regions that have direct and indirect role in neuronal synaptic plasticity. 18
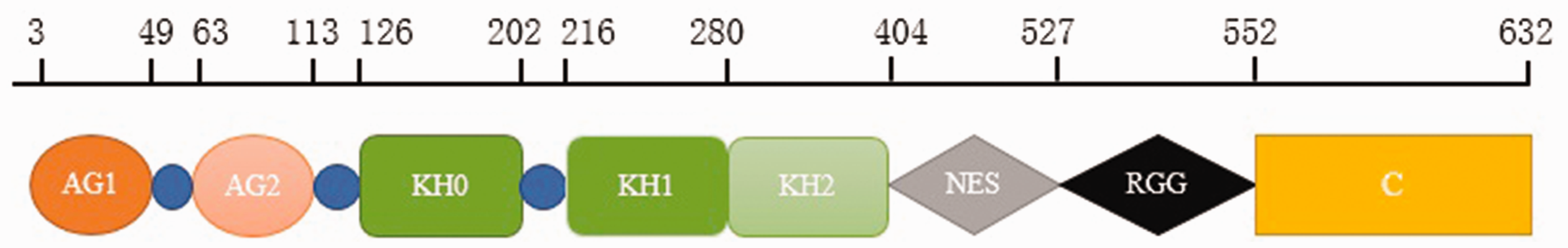
The protein architecture of FMRP. The highly modular architecture of FMRP includes two AG domains, three KH domains, and unstructured regions containing NES, RGG, and C-terminal domains. 17 KH: K homology; AG: Agenet; RGG: glycinearginine box; NES: nuclear export sequences.
Localization studies have found FMRP expressed in dorsal root ganglia (DRG), trigeminal ganglia (TG), as well as in primary afferent neurons in rats. 19 It is found to be most concentrated in the neuronal soma and dendrites.20,21 The loss of FMRP causes the dysregulation of synaptic protein synthesis, resulting in abnormal synaptic connectivity. 18
Molecularly, FMRP regulates RNA translation through interacting with various post-transcriptional factors or the messenger RNA (mRNA) structure itself.17,22,23 For example, it can modulate mRNA alternative splicing through binding with alternative-splicing-associated RNA-binding protein. 17 Alteration of this interaction results in abnormal mRNA expression in hippocampal neurons. 24 FMRP has also been shown to complex with the G-quadruplex structure within the coding or non-coding regions of mRNAs, through which it helps regulate translocation and translation of mRNAs in neurons. 25
FMRP also plays an important role in signaling pathways that impact synaptic morphology and network plasticity. Khayachi et al. provided evidence to support FMRP’s involvement in the small ubiquitin-like modifier (SUMO) pathway in the brain. 26 They showed metabotropic glutamate receptors (mGluRs)-induced sumoylation of FMRP is important in regulating maturation and elimination of dendritic spines. 26 Furthermore, FMRP is necessary for the expression of diacylglycerol kinase kappa (DGKκ), an important component of the diacylglycerol and phosphatidic acid signaling pathways. 27 The absence of FMRP in neurons eliminates group 1 mGluR-dependent DGKκ activity and reduces its expression of Dgkκ. This subsequently leads to abnormal lipid signaling in dendritic spines. 27 Lastly, work by Guo et al. demonstrated that FMRP interacts directly with transcriptional regulators (e.g., taurine-upregulated gene 1-Ski-related novel protein N complex) and regulates axonal development. 28
These studies provide compelling evidences of FMRP being essential for RNA translation and trafficking, synaptic signaling, and network plasticity of the nervous system.
FMRP in chronic pain
Fmr1 mutation- and FMRP defect-related conditions, such as FXS, are associated with fibromyalgia and peripheral neuropathy. 29 Rodriguez-Revenga et al. found that 24.4% of the female with Fmr1 mutation complained of chronic muscle pain in 398 Spanish FXS families. 30
FMRP has been implicated in the development of chronic pain. It is important in regulating synaptic plasticity in the anterior cingulate cortex, a region that is strongly implicated in pain perception and modulation.31,32 Moreover, the deficits of FMRP in Fmr1 KO mice leads to abnormal gamma aminobutyric acid (GABA) metabolism, downregulated α2, β1, and δ GABA receptors expression, and impaired GABAergic transmission in a number of pain-relevant circuits, such as seen in hippocampus.33,34 Given the critical role of GABAergic circuits in chronic pain, FMRP may be a major neuronal modulator in the nociceptive circuit.35,36
One of the hallmark characteristics of FXS is that patients often display self-injurious behaviors, which suggests abnormal pain perception. Price et al. demonstrated reduced response to neuropathic pain in Fmr1 KO mice, and it is mediated by group 1 mGluR(mGluR1/5) and mammalian target of rapamycin (mTOR). 14 Unlike in the wild-type (WT) animals, intrathecal or peripheral injection of mGluR1/5 agonist in Fmr1 KO mice fails to develop hyperalgesia. Nociceptive behaviors induced by formalin and mGluR1/5 agonist can be inhibited by mTOR inhibitor, rapamycin, in WT mice, but not in Fmr1 KO mice. 14 Collectively, this indicates that mTOR signaling pathway is defected in Fmr1 KO animals, and that FMRP is an important mediator for mGluR1/5 and mTOR-dependent nociceptive sensitization.
Lastly, recent evidence showed that interleukin 6 (IL-6) can stimulate nociceptive sensitization in DRG and TG neuronal axons, 37 while nociceptive sensitization induced by IL-6 was significantly reduced in Fmr1 KO mice. 38 This suggests that FMRP is essential in the downstream effects of nociception signaling mediated by IL-6.
The potential mechanisms of FMRP regulating chronic pain
Ion channels and neuronal excitability
Studies on animal models of FXS have shown that FMRP plays an important role in influencing neuronal excitability by regulating mRNA expression of specific ion channels, and interacting with channel subunits and altering channel mechanics. 39
Potassium channels appear to be heavily influenced by FMRP. It was demonstrated to bind to voltage-gated potassium channels mRNAs, such as Kv 3.1 and Kv 4.2, and regulate channel expression in the dendritic compartments of neurons. 39 Brown et al. showed that FMRP interacts directly with sodium-activated potassium (Slack) channel, which activates the channel and induces persistent change in firing patterns in neurons. 40 FMRP also interacts directly with the auxiliary β4 subunit of calcium-activated potassium (BK) channels, thereby increases the calcium-dependent activation of BK channels on the presynaptic terminal in hippocampal CA3 pyramidal neurons. This interaction alters the presynaptic action potential duration and calcium signaling, which subsequently changes the mechanics of neurotransmitter release. 41
FMRP also influences calcium channel metabolism in nociception pathways. By targeting membrane voltage-gated calcium (CaV) channel for proteasomal degradation, FMRP alters presynaptic CaV channel density and leads to a reduction of membrane current in DRG. Moreover, given the important role of presynaptic CaV channels in neurotransmission, it is thus not surprising to see an FMRP-induced reduction of presynaptic CaV channels can lead to a decrease in vesicle exocytosis and neurotransmitter release in both DRG and hippocampal neurons. 42
This body of work thus collectively supports the notion that changes in ion channel activity and plasma membrane potential mediated by FMRP play a vital role in nociceptive sensitization. 39
The mGluRs signaling pathways
mGluRs are involved in glutamatergic neurotransmissions as well as synaptic development and activity-dependent synaptic plasticity. 43 They are widely expressed throughout the nociceptive pathways and influence the transmission of nociceptive signals. 44 For example, Xie et al. demonstrated that hyperalgesia induced by chemotherapy drug, paclitaxel, is mediated by increased presynaptic expression and activity of mGluR5 in rat DRG and spinal cord. 45
Zhang et al. showed that increased amplitude of primary glutamate excitatory post-synaptic currents in spinal nerve ligation rats, and this phenomenon is mediated by group 3 mGluRs (mGluR4/6/7/8). 46
Group 1 mGluR-induced synaptic plasticity requires FRMP. Yang et al. demonstrated that group 1 mGluR activation increases neural network activity by activating an FMRP-depending signaling pathway. This pathway downregulates a translation inhibitor, murine double minute-2 (Mdm2), which subsequently induces protein translation. 47
Activation of the c-Jun N-terminal kinase (JNK) pathway has been implicated in the development and maintenance of chronic pain, 48 and FMRP plays an important role in JNK-mediated protein synthesis after mGluR activation. 49 In a mouse model, mGluR activation induces an increase in FMRP level, and the subsequent increase in JNK activity. This is not seen in Fmr1 KO mice. Furthermore, the loss of FMRP in Fmr1 KO animals leads to a reduction in JNK activity and a decrease in post-synaptic protein expression. This is consistent with the earlier study that showed mGluR-dependent gene transcription is regulated by JNK activity. 47
The mammalian target of rapamycin (mTOR) signaling pathways
mTOR, a serine/threonine protein kinase, is essential for regulating protein synthesis,50,51 and recent work suggests that dysfunction of mTOR and its associated pathways can lead to abnormal pain states. Studies that employed neuropathic and inflammatory pain models in rats have shown similar increase in the expression of mTOR in the spinal cord. Intrathecal administration of a mTOR inhibitor, rapamycin, significantly reduces mechanical and thermal hyperalgesia in these animals.52,53
FMRP is an important modulator for mTOR. Sharma et al. found that the activity of phosphorylated mTOR (active form) is increased in hippocampal CA1 synapses in Fmr1 KO mice, and that the mGluR-induced long-term depression in these animals is also enhanced. 43 This finding is consistent with the work by Price et al. as discussed above. 14 Together, these studies imply that FMRP plays a vital role in nociception sensitization, and it is at least in part mediated by the mTOR signaling pathway.
Conclusions and perspectives
FMRP deficiency, caused by Fmr1 mutation, can affect pain perception, as seen in FXS patients. However, the precise mechanism through which FMRP modulates pain remains unclear. FMRP has long been known as an important modulator for RNA translation and translocation as well as for a number of signaling pathways in neurons. Recent studies have demonstrated that FMRP can influence nociception processing and sensitization via various means, such as altering ion channel expressions and activities, and modulating the mTOR and the mGluRs signaling pathways. Therefore, the current review provides important insights into how Fmr1 gene and FMRP may contribute to nociceptive sensitization in chronic pain.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was supported by a grant from Projects of medical and health technology program of Zhejiang, China (WKJ-ZJ-2025).