
Abstract
Trigeminal neuralgia commonly results in pain behaviors and cognitive impairment. Convincing evidence suggests that TWIK-related spinal cord K+ (TRESK) exerts antinociceptive and neuroprotective effects. However, its possible potentials in trigeminal neuralgia remain unclear. Trigeminal neuralgia model was established in rats by generating an infraorbital nerve chronic constriction injury, and rats received intrathecal injections of TRESK-overexpressing lentivirus and siRNA expression vector-targeted against TRESK (si-TRESK) into the trigeminal ganglions. Mechanical allodynia was evaluated by mechanical withdrawal threshold. Cognitive capacity was tested using Morris water maze. The TRESK expression was determined by quantitative real-time polymerase chain reaction and Western blotting. Results showed that the mRNA and protein levels of TRESK were significantly downregulated in trigeminal ganglions in injured rats. Intrathecal treatment with TRESK reduced mechanical allodynia and relieved learning and memory deficits in trigeminal neuralgia rats, while si-TRESK injection caused neuropathic pain and cognitive deficits. In summary, the present study concluded that TRESK ameliorated pain-associated behaviors and cognitive deficits, which was useful as an alternative approach in management of trigeminal neuralgia.
Keywords
Introduction
Trigeminal neuralgia (TN) is a common chronic pain disease with an annual incidence of about 4% to 13%, which adversely affects patient’s quality of life. 1 There are two main theories about the etiology of TN. 2 Firstly, the peripheral pathogenetic mechanism is triggered by abnormal compression on gasserian ganglion and pons. Secondly, central pathogenetic mechanism theory holds that TN is a kind of sensory epileptic-like seizure caused by the pathologic paroxysmal type irritation in the trigeminal spinal cord nucleus in the brain stem. Because its pathogenesis is not completely clear, so far, there is still a lack of ideal analgesics in clinical practices.
At onset of TN, sensory neurons at all levels of the trigeminal axis showed enhanced neural excitability pain facilitation and decreased pain threshold. The increased neural excitability results in burst discharges of action potentials, leading to allodynia pain signal transduction. In mammalian sensory neurons, changes in the expression levels of ion channels, receptors, growth factors, and neuropeptides may lead to increased input resistance and decreased action potential threshold and ectopic excitation. 3
TWIK-related spinal cord K+ (TRESK), encoded by KCNK18, is the most recently identified member of two-pore domain K+ (K2P) channel family. A large quantity of evidence has revealed that TRESK is abundantly expressed in dorsal root ganglions (DRGs) which are peripheral sensory ganglia that do not belong to the spinal cord and other sensory ganglions such as trigeminal ganglions. 4 TRESK is the most recently identified member of the two-pore-domain potassium channel (K(2P)) family, constituting a significant component of background potassium currents in murine DRG neurons. 5 A dominant-negative mutation in the TRESK potassium channel is associated with familial migraine with aura, further supporting the role of this channel as a potential therapeutic target. 6 Tulleuda et al. clearly uncovered an important role of TRESK channels in determining neuronal excitability in specific DRG neurons subpopulations and showed that axonal injury downregulated TRESK channels, therefore contributing to neuronal hyperexcitability. 7 More recent studies by Royal et al. revealed that migraine-associated TRESK mutations increased neuronal excitability through alternative translation initiation and inhibition of potassium two pore domain channel subfamily K member 2 (TREK). 8 Our previous study found that mutant TRESK subunits exhibited a dominant-negative effect on the leak K+ currents through the endogenous TRESK channels, increased Rin in trigeminal ganglion neurons, and led to an increase in neuronal excitability. 9 Guo and Cao reported that overexpression of TRESK subunits increased the background K+ currents, reduced the excitability of small-diameter trigeminal ganglion neurons, and inhibited capsaicin-evoked spikes. 10 TRESK is recognized as a potential target for the development of novel analgesics. For example, Zhou et al. discovered that intrathecal injection of TRESK gene recombinant adenovirus alleviated mechanical allodynia by mediating extracellular signal-regulated kinase (ERK) and p38 mitogen-activated protein kinase (MAPK) apoptotic signaling pathway. 11 Conversely, Yang et al. uncovered that downregulation of TRESK by siRNA increased neuronal hyperexcitability and pain hypersensitivity in DRG neurons. 12
Currently, the effect of TRESK in TN has not been fully reported, which is worthy of further investigation. In the present study, we monitored pain-related behaviors and cognitive capacity in a rat model of TN generated by infraorbital nerve chronic constriction injury (ION-CCI). We examined TRESK expression in trigeminal ganglions and whether altered TRESK expression contributed to the development of neuropathic pain states and cognitive deficits.
Materials and methods
Experimental animals
Adult male Sprague-Dawley rats weighing 200 to 250 g obtained from the Laboratory Animal Center of Central South University were used in this experiment. All rats were housed individually at a room temperature of 22 ± 2°C on a 12-h light/dark cycle, with food and water available ad libitum. All experimental procedures were approved by the Ethics Committee of Xiangya Hospital.
ION-CCI and experimental design
To examine expression changes of TRESK in trigeminal ganglions in a rat model of TN, rats were randomly divided into two groups (n = 15 per group): sham operation group (sham) and ION-CCI model group (ION-CCI). The ION-CCI model was generated as a TN model. Each rat was anesthetized by intraperitoneal injection of 2% pentobarbital sodium (50 mg/kg) before surgical procedures. After skin preparation, approximately 1 cm incision was made along the anterior one-third segment of the lower margin of the right cheekbone. The lower margin of zygomaticus muscle was exposed by blunt dissection. We pushed the perimuscular fascia aside and exposed the suborbital foramen. Under the microscope, the inferior orbital nerve was ligated with two wires (4-0 chromic gut). The interval between the two ligatures was about 2 mm. Subsequently, the incision was routinely sutured. In the sham group, the suborbital nerve was left untouched. Trigeminal ganglion specimens were collected preoperatively and at postoperative 1, 7, 15, and 30 days. We sacrificed three rats for each time point.
In another experiment, to examine the underlying role of TRESK in TN, rats were randomly divided into five groups (n = 6 in each group): control, sham, ION-CCI, LV-NC, and TRESK overexpressing lentivirus (LV-TRESK). Behavior tests were performed at different time points. Measurement of mechanical withdrawal threshold (MWT) was performed 1 day before and at 1, 4, 7, 10, 13, 16, 22 and 28 days after induction of ION-CCI. Morris water maze (MWM) test was performed at one, two, three, and four days after induction of ION-CCI. At the end of the experiments (at postoperative d28), rats were sacrificed and the mRNA and protein samples were taken from trigeminal ganglion specimens for TRESK expression determination.
In another experiment, to further examine the role of TRESK in TN, rats were randomly divided into three groups (n = 6 in each group): control, siRNA, and siRNA expression vector-targeted against TRESK (si-TRESK). Behavior tests were recorded at different time points indicated in Figure 3, and at the end of the experiments (at postoperative d35), rats were sacrificed and trigeminal ganglions were harvested for TRESK expression determination.
Intrathecal injection
Isoflurane anesthetized rats were placed in the right lateral position. A 50 µL LV-TRESK with green fluorescent protein, si-TRESK, or their negative controls were injected into the trigeminal ganglia through a 25-gauge needle connected to a nerve stimulator (0.15 mA). The site of injection was 1 mm infraorbital foramen behind the lower side of the zygomatic projection. Ten minutes later, the syringe was withdrawn, and rats were subjected to ION-CCI.
Measurement of MWT
Behavioral experiments were performed by double-blind method. Rats were tested 1 day before and at 1, 4, 7, 10, 13, 16, 22, 28, and 35 days after induction of ION-CCI between 9:00 a.m. and 2:00 p.m. Rats were placed individually in a transparent plastic cage and allowed to acclimatize for 1 h. A graded series of von Frey hairs (Stoelting, Chicago, IL, USA) was applied to the ipsilateral ION territory near the center of the vibrissal pad and held for ∼2 s with a 30-s interval. One or more of the following behaviors such as shaking heads, shrinking, and scratching faces were considered as a positive response to mechanical pain. MWTs were calculated to measure mechanical sensitivity.
MWM test
One week after induction of ION-CCI and injection of LV-TRESK or si-TRESK, MWM was used to evaluate the spatial learning and memory ability. The water maze was a circular pool (180 cm diameter and 60 cm high) filled with 19°C to 22°C opaque water which was divided into four equivalent quadrants. A black escape platform (10 cm diameter and 2 cm above the water level) was placed in a quadrant. Rats were placed into the water facing the pool at different predetermined positions and allowed to swim freely for 90 s. Swimming speed and escape latency were recorded. Each test interval was 20 min, and the experiment lasted four days. On day 5, the hidden platform was removed. Rats were placed in the pool at the location opposite to the platform position and allowed to swim freely for 120 s. The time spent in the target quadrant and times across platform were then recorded.
Quantitative real-time PCR
Total RNA was extracted from trigeminal ganglion specimens using the Trizol reagent (Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s instructions and reversely transcribed into cDNAs using SuperScript III (Invitrogen). The quantification of TRESK expression levels was performed using the Real-Time Quantitative PCR SYBR Green kit (Takara, Tokyo, Japan) on the ABI 7500 Fast Real-Time PCR system (Applied Biosystems, Carlsbad, CA, USA). Primer sequences used were as follows: TRESK, forward: 5′-
Western blotting
Protein samples (50 µg) were subjected to 12% sodium dodecyl sulfate-polyacrylamide gel electrophoresis and transferred to polyvinylidene difluoride (Millipore, Bedford, MA, USA) membranes. Blots were hatched overnight with goat antimouse monoclonal TRESK antibody (Santa Cruz Biotechnology, Santa Cruz, CA, USA) followed by incubation with horseradish peroxidase-conjugated rabbit antigoat IgG antibody (Santa Cruz Biotechnology). Specific bands were visualized using chemiluminescence.
Statistical analysis
Data were exhibited as mean ± standard deviation and analyzed by SPSS 22.0 statistical software (SPSS, Chicago, USA). Student’s t test and one-way analysis of variance were used to compare the differences between groups. A P value of <0.05 was considered statistically significant.
Results
TRESK was downregulated in trigeminal ganglions in a rat model of TN
We firstly used quantitative real-time polymerase chain reaction (qRT-PCR) and Western blotting to determine the relative mRNA and protein levels of TRESK in trigeminal ganglions before and after nerve injury. As compared with the sham rats, the expressions of TRESK at mRNA (Figure 1(a)) and protein levels (Figure 1(b)) were significantly decreased in trigeminal ganglions after ION-CCI.

TRESK expression in trigeminal ganglions in trigeminal neuropathic rats. The mRNA (a) and protein levels (b) of TRESK in trigeminal ganglions preoperatively and at postoperative 1, 7, 15 and 30 days in sham-operated rats and ION-CCI rats determined by qRT-PCR and Western blotting. *P < 0.05 versus sham group. n = 3 in each time point. GAPDH: glyceraldehyde 3-phosphate dehydrogenase; ION-CCI: infraorbital nerve chronic constriction injury; TRESK: TWIK-related spinal cord K+.
TRESK overexpression ameliorated TN induced by ION-CCI
To understand the underlying role of TRESK in TN, we intrathecally injected LV-TRESK into rats followed by ION-CCI treatment, and the MWT of rats was further assessed. There was no significant difference in the mechanical thresholds between the control and sham group. Compared to rats in the sham group, animals in the ION-CCI group exhibited a significant decrease in the MWT, which lasted for the 28-day observation period. Treatment with LV-TRESK markedly ameliorated the mechanical allodynia in ION-CCI-induced TN rats (Figure 2(a)). To examine whether TRESK protected cognitive function, we compared behavioral performance in MWM tests. Rats in ION-CCI group displayed significantly longer escape latency in reaching the platform from day 1 after injury than the sham group, indicating that neuropathic pain resulted in cognitive impairment. On the contrary, rats injected with LV-TRESK presented a significant decrease in escape latency (Figure 2(b)). However, no significant differences were observed in swimming speed between all groups (Figure 2(c)). In the ION-CCI group, the swimming time in target quadrant was shortened and the time across platform was reduced compared with those in the sham group; however, both parameters were significantly prolonged after the injection of LV-TRESK (Table 1). At the end of the experiments, rats were sacrificed and the trigeminal ganglion specimens were collected for qRT-PCR and Western blotting analysis. The mRNA and protein levels of TRESK in trigeminal ganglions were confirmed to be markedly upregulated after LV-TRESK injection (Figure 2(d) and (e)).

TRESK expression in trigeminal ganglions after intrathecal injection of LV-TRESK. (a) Time course of MWT, the escape latency (b), and swimming speed (c) at different time points in the groups of control, sham, ION-CCI, LV-NC, and LV-TRESK. The mRNA (d) and protein levels (e) of TRESK in trigeminal ganglions at postoperative d28 in the groups of control, sham, ION-CCI, LV-NC, and LV-TRESK. *P < 0.05 versus sham; &P < 0.05 and $P<0.05 versus LV-NC; #P < 0.05 versus pre-op; ns, no significance. n = 6 in each group. GAPDH: glyceraldehyde 3-phosphate dehydrogenase; ION-CCI: infraorbital nerve chronic constriction injury; LV-NC: a negative control lentiviral vector; LV-TRESK: TRESK-overexpressing lentivirus; MWT: mechanical withdrawal threshold; TRESK: TWIK-related spinal cord K+.
Time in target quadrant and times across platform (n = 6, mean ± SD).
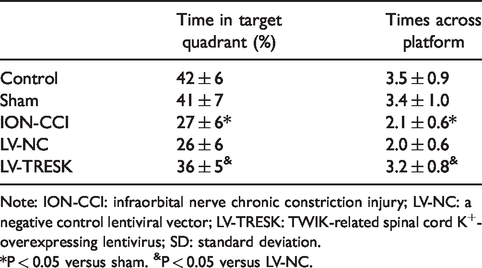
Note: ION-CCI: infraorbital nerve chronic constriction injury; LV-NC: a negative control lentiviral vector; LV-TRESK: TWIK-related spinal cord K+-overexpressing lentivirus; SD: standard deviation.
*P < 0.05 versus sham. &P < 0.05 versus LV-NC.
TRESK knockdown contributed to trigeminal neuropathic pain following ION-CCI
We further investigated whether TRESK silencing in vivo could cause neuropathic pain-like behaviors in rats. As shown in Figure 3(a), si-TRESK injection into the trigeminal ganglion was found to produce decreased MWT. Furthermore, si-TRESK group increased their latency to reach the platform as relative to the siRNA group (Figure 3(b)), while swimming speed was similar between groups (Figure 3(c)). Comparisons among probe trials showed decreased percentages of the total probe trial time in the target quadrant and platform site crossings in si-TRESK group as compared to the siRNA group (Table 2). As expected, the mRNA (Figure 3(d)) and protein levels (Figure 3(e)) of TRESK in trigeminal ganglions were confirmed to be memorably decreased after intrathecal injection of si-TRESK.
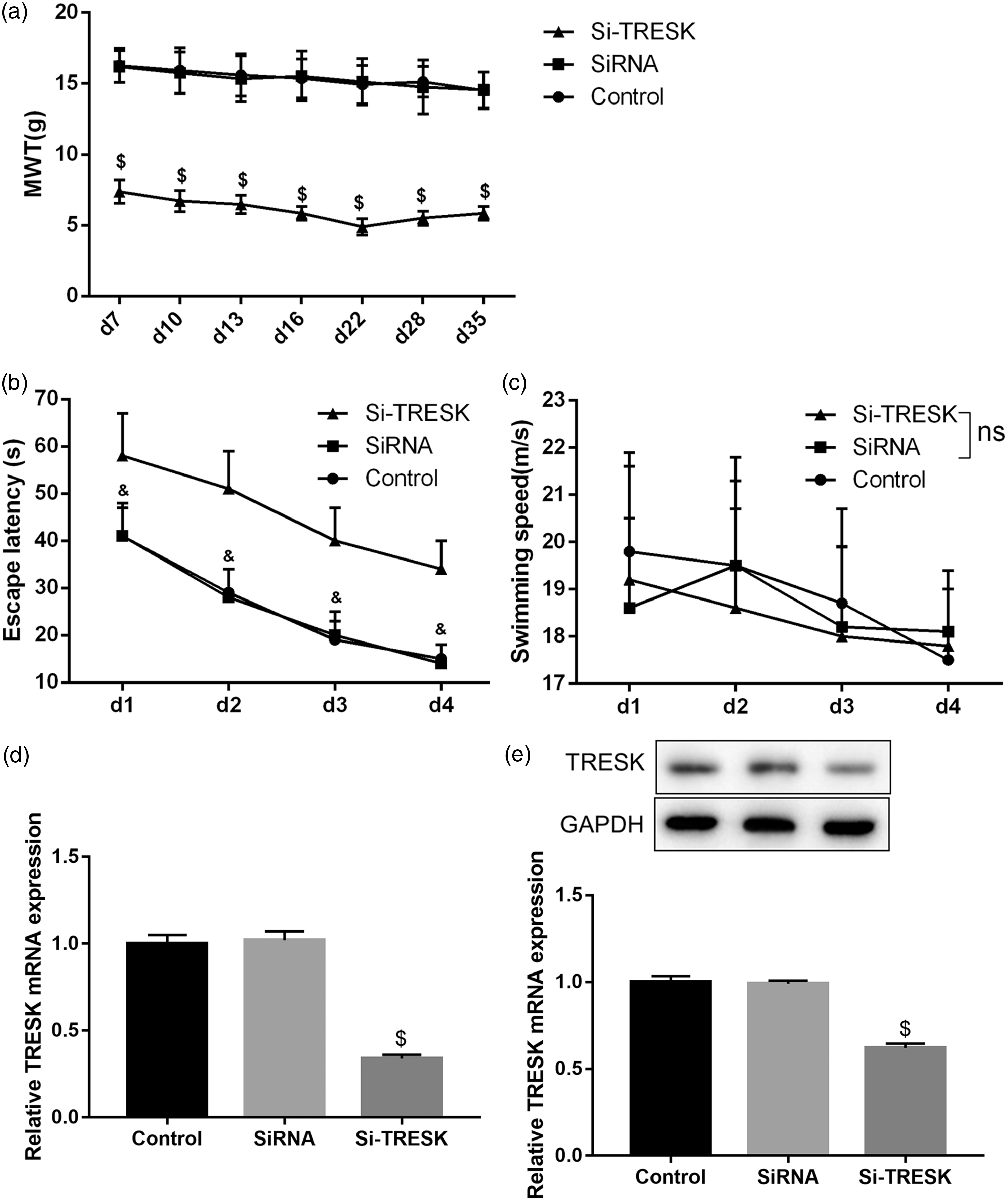
TRESK expression in trigeminal ganglions after intrathecal injection of si-TRESK. (a) Time course of MWT, the escape latency (b), and swimming speed (c) at different time points in the groups of control, siRNA, and si-TRESK. The mRNA (d) and protein levels (E) of TRESK in trigeminal ganglions at postoperative d35 in the groups of control, siRNA, and si-TRESK. $P < 0.05 versus siRNA group; &P<0.05 versus si-TRESK group; ns, no significance. n = 6 in each group. GAPDH: glyceraldehyde 3-phosphate dehydrogenase; MWT: mechanical withdrawal threshold; TRESK: TWIK-related spinal cord K+; si-TRESK: siRNA expression vector-targeted against TRESK.
Time in target quadrant and times across platform (n = 6, mean ± SD).

Note: SD: standard deviation; si-TRESK: siRNA expression vector-targeted against TWIK-related spinal cord K+.
$P < 0.05 versus siRNA.
Discussion
Neuropathic pain is widely accepted as a direct consequence of a lesion or disease affecting the somatosensory system, 13 and the exact pathological mechanisms are poorly understood. The K2P channel is involved in regulating the excitability of sensory nociceptive neurons and the conduction of resting membrane potential, which is therefore considered as an important mediator of pain perception. 14 Several lines of evidence highlighted the importance of TREK1, TREK2, and TRESK channels among K2P channels. TRESK is a subtype of K2P associated with the occurrence and development of various pain states. 15 To the best of our knowledge, the mechanism of pain modulation is not completely clear. Here, this study provided evidence that decreased TRESK channels in trigeminal ganglions contributed to the development of TN. Moreover, TRESK overexpression rescued pain and cognitive deficits in an ION-CCI-induced TN rat model, whereas TRESK knockdown produced neuronal hyperexcitability of trigeminal ganglion neurons and pain hypersensitivity in normal rats.
In this study, the CCI-ION model was used as a TN model. 16 The mRNA and protein levels of TRESK in trigeminal ganglions were firstly determined in the present study. Previous studies have reported the expression of TRESK in DRGs and spinal cords. 17 Recently, a few studies demonstrated the single-channel kinetics of TRESK in trigeminal ganglions. For example, Park et al. described the functional expression of TRESK in trigeminal ganglion neurons by recording single-channel currents. 18 In addition, TRESK overexpression reportedly repressed the excitability of trigeminal ganglion nociceptors. 10 The expression level of TRESK mRNA in trigeminal ganglions rats was detected by RT-PCR, and we confirmed that TRESK was remarkably downregulated in a rat model of TN induce by ION-CCI. The result of Western blotting was consistent with the results of qRT-PCR. These findings revealed the involvement of TRESK in the occurrence of TN, which might contribute to pain transmission.
Although many patients with neuropathic pain have a response to analgesic therapy,19,20 cognitive deficits represents the most common complaints. 21 On the first-day postoperation, the MWTs in the ION-CCI group was decreased, which persisted for 28 days. Intrathecal injection of LV-TRESK was performed before operation. The results showed that TRESK overexpression effectively decreased mechanical allodynia. Zhou et al. previously elucidated that intrathecal injection of TRESK gene recombinant adenovirus (25 µL) into the region of lumbar enlargement in a rat model of spared nerve injury could upregulate the mRNA expression of TRESK in the dorsal root ganglia and reduce tactile allodynia. 22 Zhou et al. illustrated that intrathecal injection of TRESK gene recombinant adenovirus significantly upregulated the mRNA and protein levels of TRESK in DRGs and spinal cords and reduced mechanical allodynia. 23 A large quantity of evidence has revealed that exercise in the MWM test can improve the ability of spatial learning and memory. 24 ION-CCI group improved their latency to reach the platform, indicating that TN led to learning dysfunction of the rats. However, the overexpression of TRESK led to a decrease in escape latency, suggesting that TRESK restored the spatial learning ability. Rats in the LV-TRESK group exhibited significantly higher percentages of total probe time in the target quadrant and more numbers of platform crossings, indicating that TRESK overexpression also repaired spatial reference memory deficits. However, TRESK knockdown group led to an opposite effect. Throughout the experiments, swimming speeds were similar among groups, suggesting that motor impairment might not be the leading cause of the differences in the acquisition period of the MWM.
In conclusion, our data suggested that intrathecal administration of LV-TRESK could produce neuroprotection in TN induced by ION-CCI by alleviating deficits in abilities of spatial learning and memory. These data suggest a new potential therapeutic approach to treating TN.
Footnotes
Authors’ Contributions
YL conceived and designed the experiments. YL, HJ, WR, and FR performed the experiments. YL, HJ, and FR analyzed the data. FR wrote the draft paper. All authors read and approved the final manuscript.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was funded by Natural Science Foundation of Hunan Province (2017JJ2383).