
Abstract
Spinal D-serine plays an important role in nociception via an increase in phosphorylation of the N-Methyl-D-aspartate (NMDA) receptor GluN1 subunit (pGluN1). However, the cellular mechanisms underlying this process have not been elucidated. Here, we investigate the possible role of neuronal nitric oxide synthase (nNOS) in the D-serine-induced potentiation of NMDA receptor function and the induction of neuropathic pain in a chronic constriction injury (CCI) model. Intrathecal administration of the serine racemase inhibitor, L-serine O-sulfate potassium salt (LSOS) or the D-serine degrading enzyme, D-amino acid oxidase (DAAO) on post-operative days 0–3 significantly reduced the CCI-induced increase in nitric oxide (NO) levels and nicotinamide adenine dinucleotide phosphate-diaphorase staining in lumbar dorsal horn neurons, as well as the CCI-induced decrease in phosphorylation (Ser847) of nNOS (pnNOS) on day 3 post-CCI surgery. LSOS or DAAO administration suppressed the CCI-induced development of mechanical allodynia and protein kinase C (PKC)-dependent (Ser896) phosphorylation of GluN1 on day 3 post-surgery, which were reversed by the co-administration of the NO donor, 3-morpholinosydnonimine hydrochloride (SIN-1). In naïve mice, exogenous D-serine increased NO levels via decreases in pnNOS. D-serine-induced increases in mechanical hypersensitivity, NO levels, PKC-dependent pGluN1, and NMDA-induced spontaneous nociception were reduced by pretreatment with the nNOS inhibitor, 7-nitroindazole or with the NMDA receptor antagonists, 7-chlorokynurenic acid and MK-801. Collectively, we show that spinal D-serine modulates nNOS activity and concomitant NO production leading to increases in PKC-dependent pGluN1 and ultimately contributing to the induction of mechanical allodynia following peripheral nerve injury.
Introduction
D-serine serves as a co-agonist for the glycine site on N-Methyl-D-aspartate (NMDA) receptors and as such is now being considered as an important modulator of nociceptive transmission in the central nervous system (CNS) via activation of NMDA receptors.1,2 D-serine is synthesized from L-serine by the activation of serine racemase (Srr), which is expressed in the astrocytes of spinal dorsal horn. 3 In previous studies, we have shown that chronic constriction injury (CCI) of the sciatic nerve increased the expression of Srr and concomitant D-serine production in the lumbar spinal cord dorsal horn. 4 Inhibition of D-serine by administration of the Srr inhibitor, L-serine O-sulfate potassium salt (LSOS) or the D-serine degrading enzyme, D-amino acid oxidase (DAAO) significantly suppressed the development of mechanical allodynia in peripheral nerve-injured mice. 4 In addition, sigma-1 receptor modulated the expression of Srr and D-serine production in astrocytes of spinal dorsal horn, which significantly increased protein kinase C (PKC)-dependent phosphorylation of the NMDA receptor GluN1 subunit (pGluN1), resulting in facilitation of NMDA-induced nociception.4,5 Although it has been suggested that D-serine plays an important role as a gliotransmitter that is released from astrocytes and contributes to the development of neuropathic pain via activation of NMDA receptors on postsynaptic neurons in the spinal cord dorsal horn,2,5,6 there is limited understanding of the cellular mechanisms underlying D-serine’s role in the development of central sensitization.
Nitric oxide (NO) plays an important role as a neuro-messenger in the nociceptive processing that occurs in the CNS. 7 Neuronal nitric oxide synthase (nNOS) is an NO-producing enzyme, and its activation is involved in the initiation and/or maintenance of chronic neuropathic pain induced by peripheral nerve injury. 7 In addition, it has been suggested that inhibition of nNOS significantly suppresses carrageenan-induced inflammatory pain. 8 However, other studies have shown that intrathecal (i.t.) administration of L-arginine significantly attenuates peripheral nerve injury-induced neuropathic pain. 9 Furthermore, NO derived from nNOS inhibits spinal NMDA receptor activity in the spinal cord dorsal horn primarily through S-nitrosylation. 9 In addition, inhibition of spinal NO production increases the frequency of background firing activity in dorsal horn neurons. 10 Since NO in the spinal cord clearly plays a complex and diverse role in the nociceptive system, more research is needed to investigate the mechanisms underlying NO-related nociceptive processing.
Phosphorylation is considered an important mechanism for the regulation of nNOS activation. nNOS is phosphorylated at different residues by various protein kinases. Phosphorylation of nNOS (pnNOS) at Ser847 inhibits nNOS activity by suppressing Ca2+/calmodulin binding,11,12 thus, dephosphorylation of nNOS at Ser847 is an important step in the activation of nNOS. In addition, nNOS is colocalized with NMDA receptors and is linked via binding to the scaffolding protein postsynaptic density-95. 13 Since Ca2+ influx through NMDA receptors efficiently stimulates nNOS via Ca2+/calmodulin signaling, the formation of this ternary complex contributes to the structural and functional coupling of nNOS to NMDA receptors. 13 In previous study, we demonstrated that activation of nNOS is closely related to the increase in PKC-dependent GluN1 phosphorylation and pain hypersensitivity induced by spinal sigma-1 receptor activation. 14 Since nNOS is involved in the functional potentiation of NMDA receptors, we speculate that spinal NO produced by nNOS can be a downstream effector of D-serine as it relates to spinal pain transmission, particularly in relation to D-serine-induced central sensitization.
Thus, the main goals of this study were to investigate: (1) whether nNOS activation is modulated by D-serine and plays a critical role in the D-serine-induced increase in PKC-dependent GluN1 phosphorylation accounting for the potentiation of NMDA receptor function and (2) whether this pathway contributes to the development of peripheral neuropathy. We designed experiments to determine if (1) the inhibition of D-serine with LSOS or DAAO changes the pnNOS at Ser847 in the spinal cord dorsal horn of CCI mice; (2) the NO donor, 3-morpholinosydnonimine hydrochloride (SIN-1) restores the development of neuropathic pain and PKC-dependent GluN1 phosphorylation that were suppressed by the administration of LSOS or DAAO; (3) exogenous D-serine modulates both NO production and nNOS phosphorylation; and finally (4) inhibition of nNOS with 7-nitroindazole (7-NI) or antagonism of NMDA receptors suppresses the D-serine-induced increase in mechanical hypersensitivity, NO production, and PKC-dependent GluN1 phosphorylation, which accounts for the facilitation of NMDA-induced spontaneous nociception.
Materials and methods
Experimental animals and peripheral nerve injury model
Four-week-old male ICR mice (20–25 g) were purchased from the Seoul National University (Seoul, Republic of Korea) Laboratory Animal Center. Animals had free access to food and water and were kept in temperature and light-controlled rooms (23±2°C, 12/12h light/dark cycle with lights on at 08:00) for at least three days prior to the beginning of the experiment. The experimental protocols for animal usage were reviewed and approved by the SNU Animal Care and Use Committee and were consistent with the Guide for the Care and Use of Laboratory Animals published by the U.S. National Institutes of Health (NIH publication No. 96–01, revised 1996).
A CCI of the common sciatic nerve was performed in mice according to the method originally described in rats by Bennett and Xie with minor modification. 15 Although chromic gut is primarily used as a ligation material in rat CCI neuropathic pain models, 16 several investigators have shown that silk or prolene sutures work better in the murine CCI model.17–19 Thus, in the present study, silk was used as the ligation suture material, since silk ligatures produce more stable neuropathic pain behaviors than catgut ligatures in murine CCI models. 20 Briefly, mice were anesthetized with 3% isoflurane in a mixture of N2O/O2 gas. The right sciatic nerve was exposed and three loose ligatures of 6-0 silk were placed around the nerve with a 1.0–1.5 mm interval between each ligature. Sham surgery was performed by exposing the sciatic nerve in the same manner, but without ligating the nerve.
Drugs and i.t. administration
The following drugs were used: LSOS (10 nmol, a Srr inhibitor); DAAO (0.1 U, a D-serine degrading enzyme); D-serine (500 nmol); N-Methyl-D-aspartic acid (NMDA; 0.4 nmol, an NMDA receptor agonist); SIN-1 (50 nmol, an NO donor); 7-NI (20, 200 nmol, a specific nNOS inhibitor); 7-chlorokynurenic acid (7-CK; 1, 10 nmol, a selective and noncompetitive antagonist of the glycine binding site of the NMDA receptor); (+)-MK-801 hydrogen maleate (MK-801; 1, 10 nmol, a selective antagonist that binds to a site located within the NMDA associated ion channel). LSOS was purchased from Santa Cruz Biotechnology Inc. (CA, USA), and NMDA and 7-CK were purchased from Tocris Cookson Ltd. (Bristol, UK). The remaining drugs were obtained from Sigma–Aldrich (St. Louis, MO, USA). The doses of all drugs used in the present study were selected based on doses used in previous studies from our laboratories showing that these doses produce maximal effects with no detectable side effects.4,5,14,21–23 7-NI was dissolved in 5% DMSO in Corn oil, and 7-CK was dissolved in 5% DMSO in saline. The remaining drugs were dissolved in physiological saline. The injection volume was 5 µl and all drugs were administrated intrathecally. In CCI mice, drugs were administrated twice a day on post-operative days 0–3, during the induction phase of neuropathic pain development. In naïve mice, 7-NI (200 nmol), 7-CK (10 nmol), or MK-801 (10 nmol) was administered 10 min prior to i.t. injection of D-serine.
Intrathecal drug administration was performed using a 50 µl Hamilton syringe connected to a 30-gauge needle based on the technique previously described by Hylden and Wilcox. 24 Briefly, the mouse was held tightly between the thumb and middle finger at the level of both iliac crests, and the fifth lumbar spinous process was palpated with the index finger. The needle was inserted through the vertebral column into the L5-6 intervertebral space and successful insertion of the needle into the i.t. space was determined by a tail flick response. Each drug was slowly injected over a 10-s period. Then, the needle was carefully removed from the spinal cord. The drug control groups received an identical injection of vehicle without drug.
Behavioral assessments
To assess nociceptive responses to innocuous mechanical stimuli (mechanical allodynia), we measured paw withdrawal response frequency (PWF) by using a von Frey filament with a force of 0.16 g (North Coast Medical, Morgan Hill, CA, USA) as described previously. 25 Mice were placed in acrylic cylinders on a wire mesh floor and allowed to habituate before testing. A von Frey filament was applied to the plantar surface of the hind paw for a 3-s period before being removed, and we recorded whether there was a withdrawal of the hind limb to the application of the filament. The filament was applied 10 times to the hind paw with a 10-s interval between each application. Then, the number of paw withdrawal responses was counted, and the results were expressed as a percent withdrawal response frequency (PWF, %), which represented the percentage of paw withdrawals out of a maximum of 10 (applied to the ipsilateral hind paw) in CCI mice and a maximum of 20 (10 for the right hind paw and 10 for the left hind paw) in naïve mice.
Nociceptive behavioral tests were performed on the ipsilateral hind paw one day before CCI surgery to obtain normal baseline values of paw withdrawal responses to mechanical and thermal stimuli. Then, animals were randomly assigned to experimental and control groups. Post-operative behavioral tests were performed three days post-surgery. In naïve mice, mechanical hypersensitivity was measured before and at 15, 30, 60, 90, 120, and 180 min after D-serine (or vehicle) administration. All behavioral analyses were performed blindly.
To assess nociceptive responses to noxious heat stimuli (thermal hyperalgesia) in CCI mice, we measured paw withdrawal response latency (PWL, s) using a plantar analgesia meter (Model 390, IITC Life Science Inc., Woodland Hills, CA, USA) as previously described by Hargreaves et al. with minor modifications. 26 Mice were placed in acrylic cylinders on a glass floor and allowed to habituate before testing. A radiant heat source was positioned under the floor beneath the hind paw. The test was duplicated in the ipsilateral hind paw of each mouse, and the mean withdrawal latency was calculated. A cutoff latency in the absence of a response was set at 20 s to prevent tissue damage.
NMDA-induced spontaneous nociceptive behaviors
Each mouse was acclimated to an acrylic observation chamber for at least 30 min before the first drug injection. NMDA (0.4 nmol in 5 µl physiological saline) was injected intrathecally in order to produce spinally mediated NMDA-induced nociceptive behaviors. Following injection, animals were immediately placed in an observation chamber and nociceptive behaviors including caudally directed licking, scratching, and biting were recorded for a 10-min period. The cumulative response time (s) of these behaviors was measured. D-serine (500 nmol) was intrathecally injected 30 min prior to NMDA injection. 7-NI (20, 200 nmol), 7-CK (1, 10 nmol), or MK-801 (1, 10 nmol) was administered 10 min prior to i.t. injection of D-serine.
Measurement of NO in the spinal cord
NO is highly reactive and this complicates its direct measurements. In the present study, NO was determined in the spinal cord dorsal horns from the lumbar enlargement using a NO detection kit (cat# ADI-917–020, Enzo Life sciences Inc., USA). This NO (total) detection kit is based on the enzymatic conversion of nitrate to nitrite by the enzyme nitrate reductase, followed by the Griess reaction to form a colored azo dye product. Animals were euthanized on day 3 post-CCI surgery or 30 min after D-serine injection. Mice were deeply anesthetized with 3% isoflurane in a mixture of N2O/O2 gas and perfused transcardially with calcium-free Tyrode’s solution. The isolated spinal cords were homogenized in phosphate-buffered saline (PBS) by sonication. Homogenates were subsequently centrifuged at 400g for 10 min at 4°C and, then, the supernatant was used for NO detection following the manufacturer’s recommendation.
Western blot assay
For Western blot analysis, separate groups of animals were deeply anesthetized with 3% isoflurane in a mixture of N2O/O2 gas, and mice were euthanized on day 3 post-CCI surgery or 30 min after D-serine injection. Animals were perfused transcardially with calcium-free Tyrode’s solution, and then the spinal cords were collected into an ice-cooled, saline-filled glass dish. The Western blot assay was performed as described in a previous report from our laboratories. 25 The spinal cord dorsal horns from the lumbar enlargement were homogenized in lysis buffer (20 mM Tris-HCl, 10 mM EGTA, 2 mM EDTA, pH 7.4, and proteinase inhibitors) containing 1% Triton X-100. The homogenates were subsequently centrifuged at 15,000 rpm for 40 min at 4°C, and the supernatant was used for Western blot analysis.
The protein concentration was estimated using the Bradford dye assay (Bio-Rad Laboratories, Waltham, MA, USA). Spinal cord homogenates (25–30 µg protein) were separated using 10% SDS-polyacrylamide gel electrophoresis and transferred to nitrocellulose membrane. After the blots had been washed with TBST (10 mM Tris-HCl, pH 7.6, 150 mM NaCl, and 0.05% Tween-20), the membranes were blocked with 5% skimmed milk for 1 h at room temperature (RT) and incubated at 4°C overnight with a primary antibody specific for PKC-dependent pGluN1 (rabbit polyclonal anti-pGluN1 Ser896 antibody, 1:1,000, cat# ABN88, Millipore Co., USA), GluN1 (rabbit polyclonal anti-GluN1 antibody, 1:1,000, cat# 07–362, Upstate Biotechnology, USA), pnNOS (rabbit polyclonal anti-pnNOS Ser847 antibody, 1:1,000, cat# ab16650, Abcam plc., USA), nNOS (mouse monoclonal anti-nNOS antibody, 1:3,000, cat# 610308, BD Biosciences, USA), or β-actin (mouse monoclonal anti-β-actin antibody, 1:5,000, cat# sc-47778, Santa Cruz Biotechnology Inc., USA). After washing with TBST, membranes were incubated for 4 h at 4°C with horseradish peroxidase-conjugated anti-rabbit or anti-mouse antibody (1:10,000, Santa Cruz Biotechnology Inc.). The bands were visualized by an enhanced chemiluminescence (Thermo Scientific, USA) and scanned with a ChemiDoc™ XRS+ imaging system (Bio-Rad). The positive pixel area of specific bands was measured using ImageJ software (ImageJ 1.45s; National Institutes of Health, USA) and normalized against the corresponding β-actin loading control bands. For analysis of pGluN1 (Ser896) or GluN1 expression, the value of the control groups was set at 100% and, then, the percent change relative to the control groups was calculated for each group. To analyze activation of nNOS, the ratio of pnNOS (Ser847) to nNOS expression was calculated. The value of the ratio of pnNOS to nNOS expression in the control groups was set at 100%. Thus, the percent change in pnNOS to nNOS expression was examined for each group.
NADPH-diaphorase staining and image analysis
Nicotinamide adenine dinucleotide phosphate (NADPH)-diaphorase staining was performed to demonstrate the presence of functional NOS enzyme as described previously with minor modifications. 27 Mice were deeply anesthetized with 3% isoflurane in a mixture of N2O/O2 gas at day 3 post-CCI surgery and perfused transcardially with calcium-free Tyrode’s solution and subsequently with fixative containing 4% paraformaldehyde in 0.1 M phosphate buffer (pH 7.4). The spinal cords were collected after perfusion, post-fixed in the identical fixative overnight, and then placed in 30% sucrose in PBS (pH 7.4) at 4°C. Serial transverse sections (40 µm) of the L4-5 spinal cord were cut using a cryostat (Leica CM1520, Leica Biosystems, Germany). Spinal tissue sections were washed in 0.1 M Tris buffer (pH 7.4) and incubated in β-NADPH (1 mg/ml, Sigma–Aldrich Co.), nitro blue tetrazolium (0.25 mg/ml, Sigma–Aldrich Co.), and 0.5% Triton X-100 for 1 h at 37°C in the dark. After tissue sections had been washed with PBS, several sections were blocked with 3% normal goat serum for 1 h at RT and incubated overnight at RT with a primary antibody specific for c-Fos (rabbit polyclonal anti-c-Fos antibody, 1:10,000, cat# PC38, Calbiochem, USA), NeuN (mouse monoclonal anti-NeuN antibody, 1:1,000, cat# MAB377, Millipore Co.), GFAP (mouse monoclonal anti-GFAP antibody, 1:1,000, cat# MAB360, Millipore Co.), or ionized calcium-binding adapter molecule 1 (Iba-1; rabbit anti-Iba1 antibody, 1:1,000, cat# 019–19741, Wako Pure Chemical Industries, Ltd., Japan). After washing with PBS, the sections were incubated in biotinylated anti-mouse IgG (1:200, Vector Laboratories, USA) or anti-rabbit IgG (1:200, Vector Laboratories) for 1 h at RT and then incubated with avidin-biotin-peroxidase complex. The immunoreactivity was visualized by incubating with PBS containing 3,3-diaminobenzidine and H2O2. Tissue sections were mounted on slides and examined under a Leica DM500 microscope (Leica Biosystems, Germany).
For analysis of NADPH-diaphorase stained cells in the spinal cord dorsal horn, three to five spinal cord sections were randomly selected from each animal and analyzed using a computer-assisted image analysis system (Metamorph version 7.7.2.0; Molecular Devices Corporation, PA, USA). To maintain a constant threshold for each image and to compensate for subtle variability of the staining, we only counted cells that were 70% darker than the average grey level of each image after background subtraction and shading correction. The positive cells were quantified in the following three dorsal horn regions: (1) the superficial dorsal horn (SDH; laminae I and II); (2) the nucleus proprius (NP, laminae III and IV); and (3) the neck region (NECK, laminae V and VI). The average number of positive cells from each animal was obtained, and these values were averaged across each group and presented as group data. All analytical procedures described above were performed blindly without knowledge of the experimental conditions.
Statistical analysis
Data are expressed as the mean ± standard error of the mean. Statistical analysis was performed using Prism 5.0 (Graph Pad Software, San Diego, CA, USA). Repeated measures two-way analysis of variance (ANOVA) was performed to determine differences in the data from the time course changes in pain hypersensitivity, and post hoc analysis was performed using the Bonferroni’s multiple comparison test in order to determine the P value among experimental groups. Statistical analyses for other data were performed with One-way ANOVA, followed by the Newman-Keuls multiple comparison test for post hoc analysis. P values of < 0.05 were considered statistically significant.
Results
Effect of i.t. administration of LSOS or DAAO on the development of neuropathic pain and total NO production in the spinal cord dorsal horn of CCI mice
Peripheral nerve injury increased paw withdrawal response frequency (PWF, %) to innocuous mechanical stimuli (mechanical allodynia) and decreased paw withdrawal response latency (PWL, s) to noxious thermal stimuli (thermal hyperalgesia) as compared to the Sham group (Figure 1(a) and (b); **P < 0.01, ***P < 0.001 vs. Sham group). Repeated i.t. administration of the Srr inhibitor, LSOS or the D-serine degrading enzyme, DAAO on post-operative days 0–3 (b.i.d., during the induction phase) in CCI-induced peripheral nerve-injured mice significantly suppressed the development of mechanical allodynia (Figure 1(a); ##P < 0.01 vs. VEH-treated CCI group). In contrast, D-serine inhibition with LSOS or DAAO administration had no effect on the development of thermal hyperalgesia in CCI mice (Figure 1(b)). CCI-induced sciatic nerve injury significantly increased total NO concentration (measured indirectly as the concentration of its stable decomposition product nitrate) in the spinal cord dorsal horns as compared with that of the sham group, and this increase was significantly reduced by administration with LSOS or DAAO (Figure 1(c); *P < 0.05 vs. Sham group, #P < 0.05, ##P < 0.01 vs. VEH-treated CCI group).
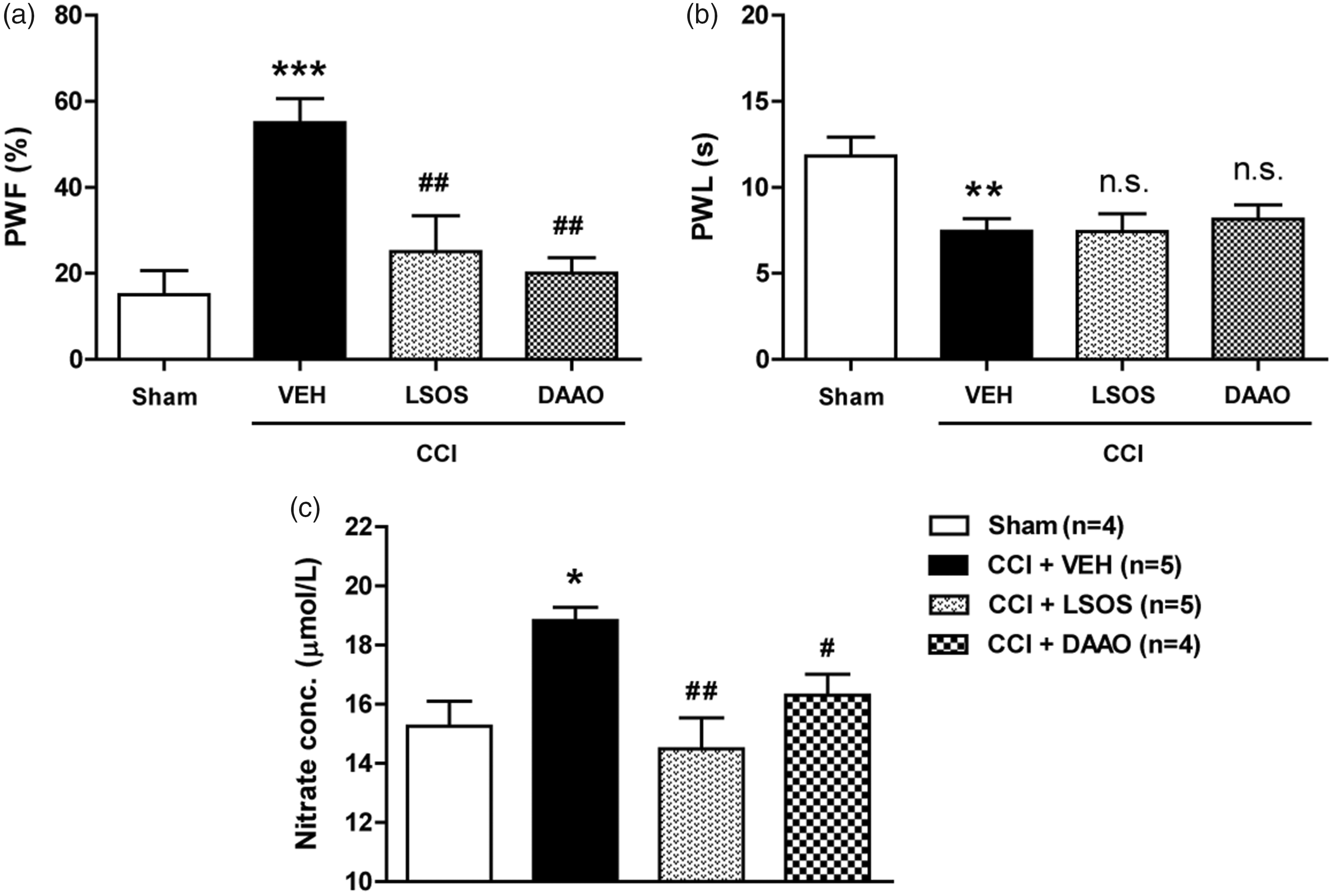
Graphs illustrating the effect of i.t. administration of the Srr inhibitor, LSOS (10 nmol) or the D-serine degrading enzyme, DAAO (0.1 U) on the development of neuropathic pain and total NO production in the spinal cord dorsal horn of CCI mice. (a and b) CCI-induced sciatic nerve injury significantly increased PWF (%) to innocuous mechanical stimuli (a) and decreased PWL (s) to noxious thermal stimuli (b). Intrathecal administration of LSOS or DAAO on post-operative days 0–3 attenuated the development of mechanical allodynia but had no effect on the development of thermal hyperalgesia. n = 6 mice/group. (c) The CCI-induced increase in total NO concentration (measured indirectly as the concentration of its stable decomposition product nitrate) in the spinal cord dorsal horns was significantly reduced by treatment with LSOS or DAAO. n = 4–5 mice/group. *P < 0.05, **P < 0.01, ***P < 0.001 vs. Sham group; #P < 0.05, ##P < 0.01 vs. VEH-treated CCI group; n.s. vs. VEH-treated CCI group. n.s.: not significant; PWF: paw withdrawal frequency; PWL: paw withdrawal latency; LSOS: L-serine O-sulfate potassium salt; DAAO: D-amino acid oxidase; CCI: chronic constriction injury; VEH: vehicle.
Effect of i.t. administration of LSOS or DAAO on nNOS phosphorylation (Ser847) and NADPH-diaphorase staining in the spinal cord dorsal horn of CCI mice
To investigate the effect of D-serine on nNOS activation in a mouse model of neuropathic pain, we intrathecally injected LSOS or DAAO on post-operative days 0–3 in CCI-induced peripheral nerve-injured mice and examined the pnNOS and NADPH-diaphorase staining as an indirect measure of NOS function as previously reported. 28 pnNOS at Ser847 inhibits nNOS activity by suppressing Ca2+/calmodulin binding, thus, dephosphorylation of nNOS at Ser847 is an important step in the activation of nNOS.11,12 Because the total protein expression of nNOS did not change following sciatic nerve injury, a decrease in the ratio of pnNOS to nNOS is likely to reflect an increase in the active forms of nNOS.12,14 CCI-induced sciatic nerve injury significantly decreased the ratio of pnNOS to nNOS, and this decrease was significantly reduced by repeated daily (from day 0 to day 3 post-surgery) administration of LSOS or DAAO (Figure 2(a); *P < 0.05 vs. Sham, #P < 0.05 vs. VEH-treated CCI group).

Graphs illustrating the effect of i.t. administration of the Srr inhibitor, LSOS (10 nmol) or the D-serine degrading enzyme, DAAO (0.1 U) on the phosphorylation (Ser847) of nNOS and NADPH-diaphorase staining in the spinal cord dorsal horn of CCI mice. (a) Results of Western blot analysis showed that the ratio of pnNOS to nNOS was decreased in the spinal cord dorsal horn following CCI and this CCI-induced decrease was completely blocked by repeated daily LSOS or DAAO administration on post-operative days 0–3. n = 6 mice/group. (b) Photomicrographs illustrate NADPH-diaphorase staining in representative spinal cord sections. The black dotted lines in the first panel depict the location of each of the three regions analysed in the spinal cord dorsal horn: the SDH (lamina I–II), the NP (lamina III–IV) and the NECK (lamina V–VI). Red arrows indicate representative NADPH-diaphorase-positive cells. Scale bar = 100 µm. (c) A graph showing that the number of NADPH-diaphorase-positive cells was significantly increased in the SDH region of the spinal cord following CCI and this increase was greatly attenuated by LSOS or DAAO treatment. n = 6 mice/group. (d) Photomicrographs showing the colocalization of c-Fos or NeuN (a neuronal marker, brown) with NADPH-diaphorase positive staining (purple) in the spinal cord dorsal horn. The red arrows depict examples of colocalization. There was no colocalization of NADPH-diaphorase staining with GFAP (an astrocyte marker, brown) or Iba-1 (a microglial cell marker, brown). Scale bar = 25 µm. *P < 0.05, ***P < 0.001 vs. Sham group; #P < 0.05, ###P < 0.001 vs. VEH-treated CCI group. LSOS: L-serine O-sulfate potassium salt; DAAO: D-amino acid oxidase; CCI: chronic constriction injury; SDH: superficial dorsal horn; NP: nucleus proprius; NECK: neck region; NADPH: nicotinamide adenine dinucleotide phosphate; nNOS: neuronal nitric oxide synthase; pnNOS: phosphorylation of nNOS; NeuN: neuronal nuclei; VEH: vehicle; GFAP: glial fibrillary acidic protein; Iba-1: ionized calcium-binding adapter molecule 1.
In addition, the number of NADPH-diaphorase-positive cells was increased in the SDH of CCI mice, and this increase was attenuated by both LSOS and DAAO administration (Figure 2(b) and (c); ***P < 0.001 vs. Sham group, ###P < 0.001 vs. VEH-treated CCI group). As illustrated in Figure 2(d), NADPH-diaphorase-positive cells were colocalized with both Fos- and NeuN-immunoreactivity, while there was no colocalization of NADPH-diaphorase staining with GFAP or Iba-1 immunostaining. These results demonstrate that spinal D-serine contributes to the CCI-induced development of mechanical allodynia and the increase in nNOS activity in the SDH neurons of CCI mice.
Effect of i.t. co-administration of SIN-1 with LSOS or DAAO on the development of neuropathic pain and pGluN1 expression in the dorsal horn of CCI mice
In a previous study, we demonstrated that D-serine increases PKC-dependent phosphorylation (Ser896) of the NMDA receptor GluN1 subunit (pGluN1), which contributes to the potentiation of NMDA receptor function. 5 Next, we examined whether NO contributes to the increase in NMDA receptor function and the development of neuropathic pain as a downstream effector of D-serine. To do this, we intrathecally injected the NO donor, SIN-1 in combination with LSOS or DAAO on post-operative days 0–3 (b.i.d., during the induction phase) in CCI mice. LSOS or DAAO administration significantly attenuated the CCI-induced development of mechanical allodynia (Figure 3(a); **P < 0.01 vs. Sham group, ##P < 0.01 vs. VEH-treated CCI group). Treatment with the NO donor, SIN-1 (50 nmol in combination with LSOS or DAAO) mimicked the effect of D-serine in CCI mice by inducing mechanical allodynia (Figure 3(a); ‡‡P < 0.01 vs. LSOS or DAAO-treated CCI group). Conversely, administration of LSOS or DAAO alone or in combination with SIN-1 did not effect the development of CCI-induced thermal hyperalgesia (Figure 3(b); ***P < 0.001 vs. Sham group).
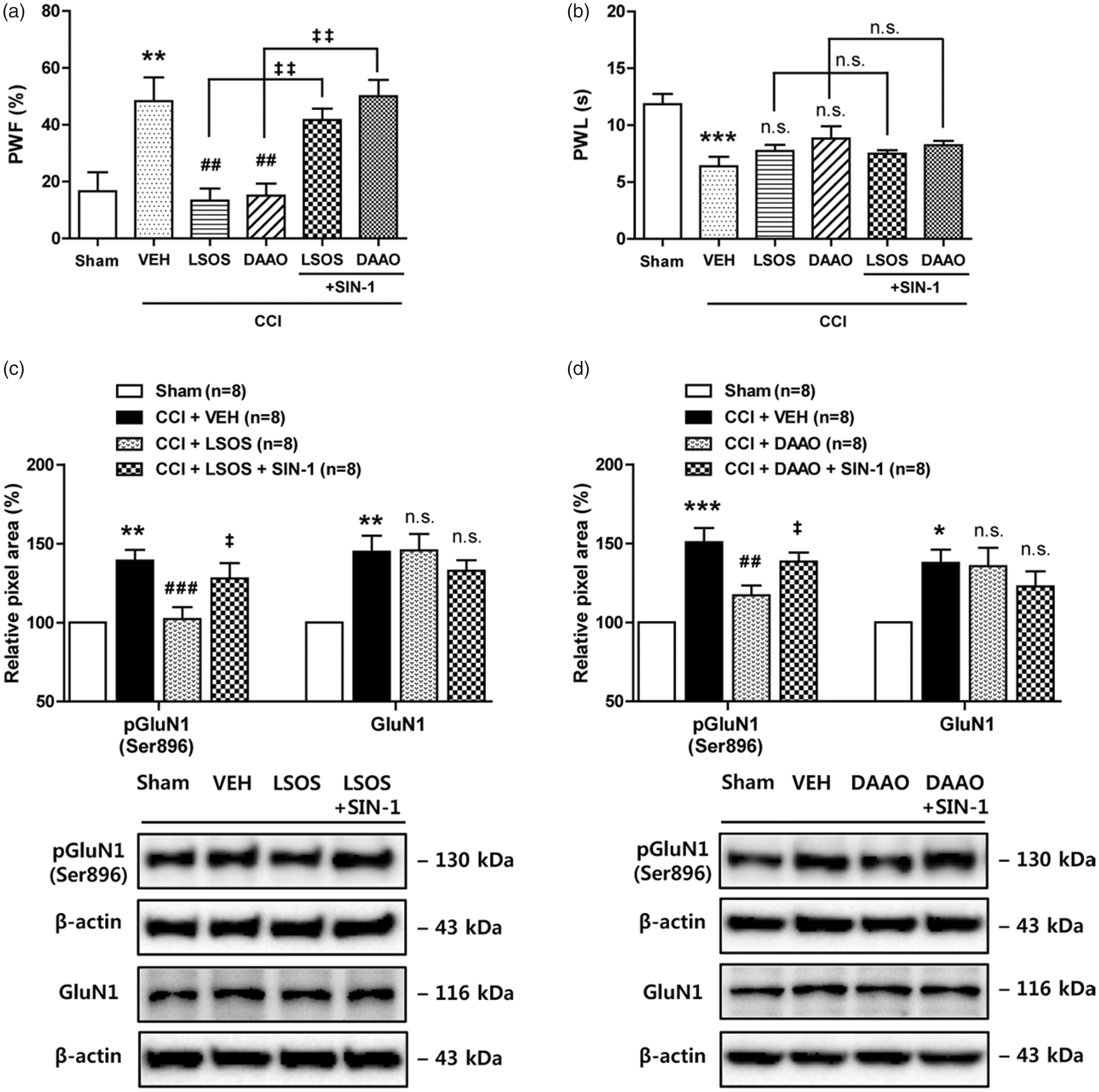
Graphs illustrating the effect of i.t. administration of the NO donor, SIN-1 (50 nmol) in combination with LSOS (10 nmol) or DAAO (0.1 U) on the development of neuropathic pain and the expression of PKC-dependent (Ser896) pGluN1 and GluN1 in CCI mice. (a) Repeated daily administration of LSOS or DAAO on post-operative days 0–3 blocked the increase in PWF (%) that occurred in VEH-treated CCI mice. Treatment with SIN-1 in combination with LSOS (LSOS + SIN-1) or DAAO (DAAO + SIN-1) restored the CCI-induced mechanical allodynia that was blocked by LSOS or DAAO in CCI mice. n = 6 mice/group. (b) Conversely, the decrease in PWL (s) to heat stimuli was unaffected by repeated i.t. injection of LSOS or DAAO alone or in combination with SIN-1. n = 6 mice/group. (c and d) Results of Western blot analysis showed that i.t. LSOS or DAAO administration significantly reduced the CCI-induced increase in PKC-dependent (Ser896) pGluN1 as compared with the VEH-treated group. Treatment with SIN-1 in combination with LSOS (LSOS + SIN-1) or DAAO (DAAO + SIN-1) restored the CCI-induced increase in pGluN1 that was originally blocked by LSOS or DAAO alone, while the CCI-induced increase in GluN1 expression was unaffected by repeated i.t. injection of LSOS or DAAO alone or in combination with SIN-1. n = 8 mice/group. *P < 0.05, **P < 0.01, ***P < 0.001 vs. Sham group; ##P < 0.01, ###P < 0.001 vs. VEH-treated CCI group; ‡P < 0.05, ‡‡P < 0.01 vs. LSOS or DAAO-treated CCI group; n.s. vs. VEH-treated CCI group. n.s.: not significant; VEH: vehicle; PWF: paw withdrawal frequency; PWL: paw withdrawal latency; LSOS: L-serine O-sulfate potassium salt; DAAO: D-amino acid oxidase; CCI: chronic constriction injury.
LSOS or DAAO administration significantly attenuated the CCI-induced increases in PKC-dependent (Ser896) pGluN1 (Figure 3(c) and (d); **P < 0.01, ***P < 0.001 vs. Sham group, ##P < 0.01, ###P < 0.001 vs. VEH-treated CCI group). Treatment with the NO donor, SIN-1 (50 nmol in combination with LSOS or DAAO) mimicked the effect of D-serine in CCI mice by increasing pGluN1 (Figure 3(c) and (d); ‡P < 0.05 vs. LSOS or DAAO-treated CCI group). Conversely, administration of LSOS or DAAO alone or in combination with SIN-1 did not effect the development of the CCI-induced increase in total GluN1 expression (Figure 3(c) and (d); *P < 0.05, **P < 0.01 vs. Sham group). These results suggest that spinal NO is a downstream effector of D-serine and mediates D-serine’s effects on both CCI-induced mechanical allodynia and CCI-induced PKC-dependent phosphorylation of the GluN1 subunit.
Effect of i.t. administration of D-serine on total NO production and phosphorylation (Ser847) of nNOS in the spinal cord dorsal horn of naïve mice
To investigate whether exogenous D-serine increases total NO production and nNOS activity, we examined the effect of i.t. D-serine on spinal nitrate concentration and the pnNOS at Ser847. Intrathecal administration of D-serine significantly increased total NO levels at the 30-min post-injection time point, and this increase was attenuated by pretreatment with DAAO (Figure 4(a); *P < 0.05 vs. VEH + VEH-treated group, #P < 0.05 vs. VEH + D-ser-treated group). In addition, exogenous D-serine decreased the ratio of pnNOS to nNOS, and this decrease was significantly reduced by i.t. treatment with DAAO (Figure 4(b); *P < 0.05 vs. VEH + VEH-treated group, #P < 0.05 vs. VEH + D-ser-treated group). These results suggest that spinal D-serine increases nNOS activity by decreasing the phosphorylated form of nNOS at Ser847.
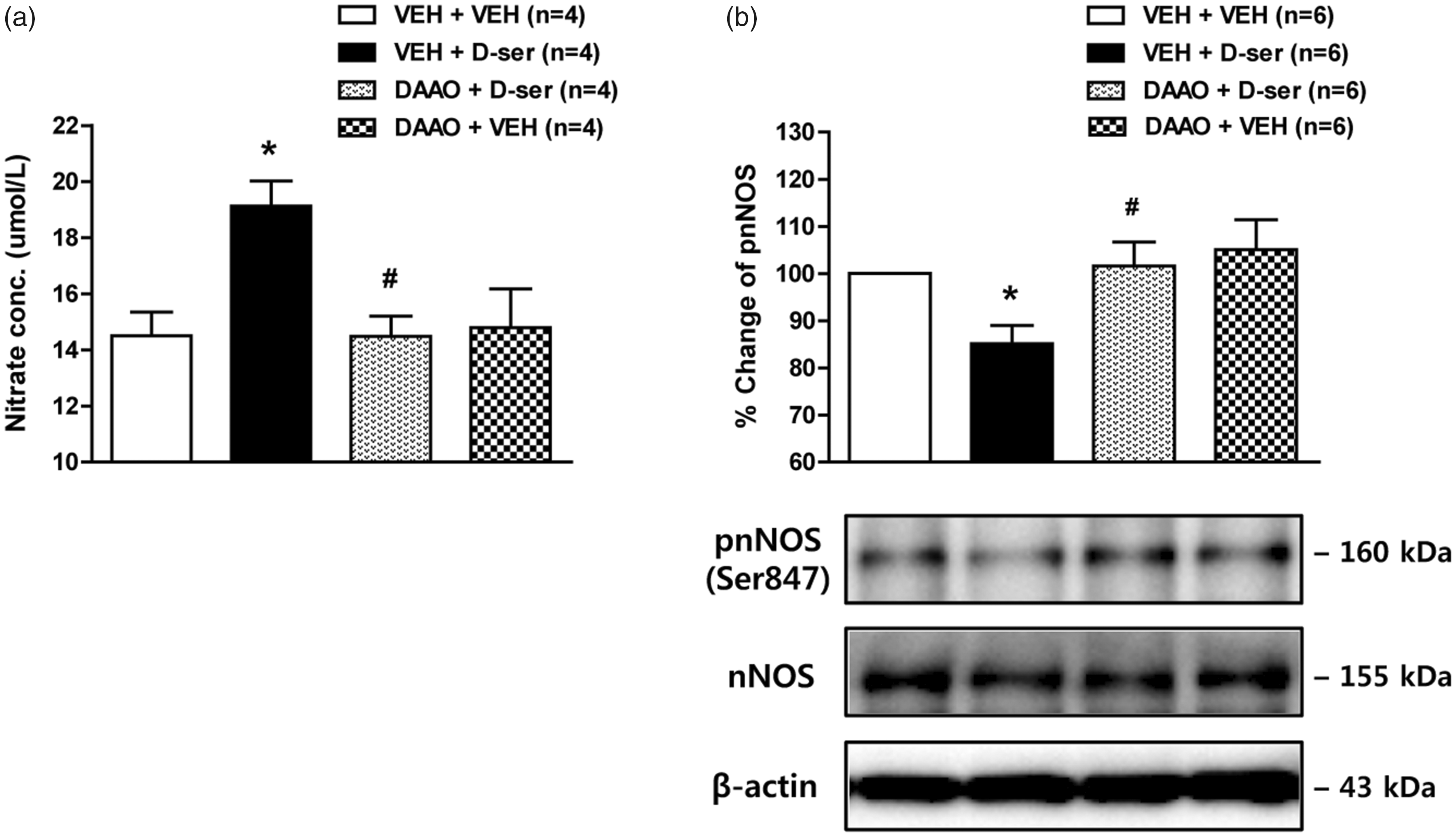
Graphs illustrating the effect of i.t. administration of D-serine (500 nmol) on total NO production and phosphorylation (Ser847) of nNOS in the spinal cord dorsal horn of naïve mice. (a) Exogenous D-serine significantly increased total NO concentration (measured indirectly as the concentration of its stable decomposition product nitrate) in the lumbar spinal cord dorsal horn at the 30 min post-injection time point and this increase was suppressed by pretreatment with the D-serine degrading enzyme, DAAO (0.1 U). n = 4 mice/group. (b) Results of Western blot analysis showed that the ratio of pnNOS to nNOS was decreased in the spinal cord dorsal horn following D-serine injection and that this decrease was completely blocked by pretreatment with DAAO (0.1 U). n = 6 mice/group. *P < 0.05 vs. VEH + VEH-treated group; #P < 0.05 vs. VEH + D-ser-treated group. VEH: vehicle; DAAO: D-amino acid oxidase; nNOS: neuronal nitric oxide synthase; pnNOS: phosphorylation of nNOS; D-ser: D-serine.
Effect of i.t. administration of 7-NI on D-serine-induced increases in mechanical hypersensitivity, total NO production, pGluN1 expression, and NMDA-induced spontaneous nociception in naïve mice
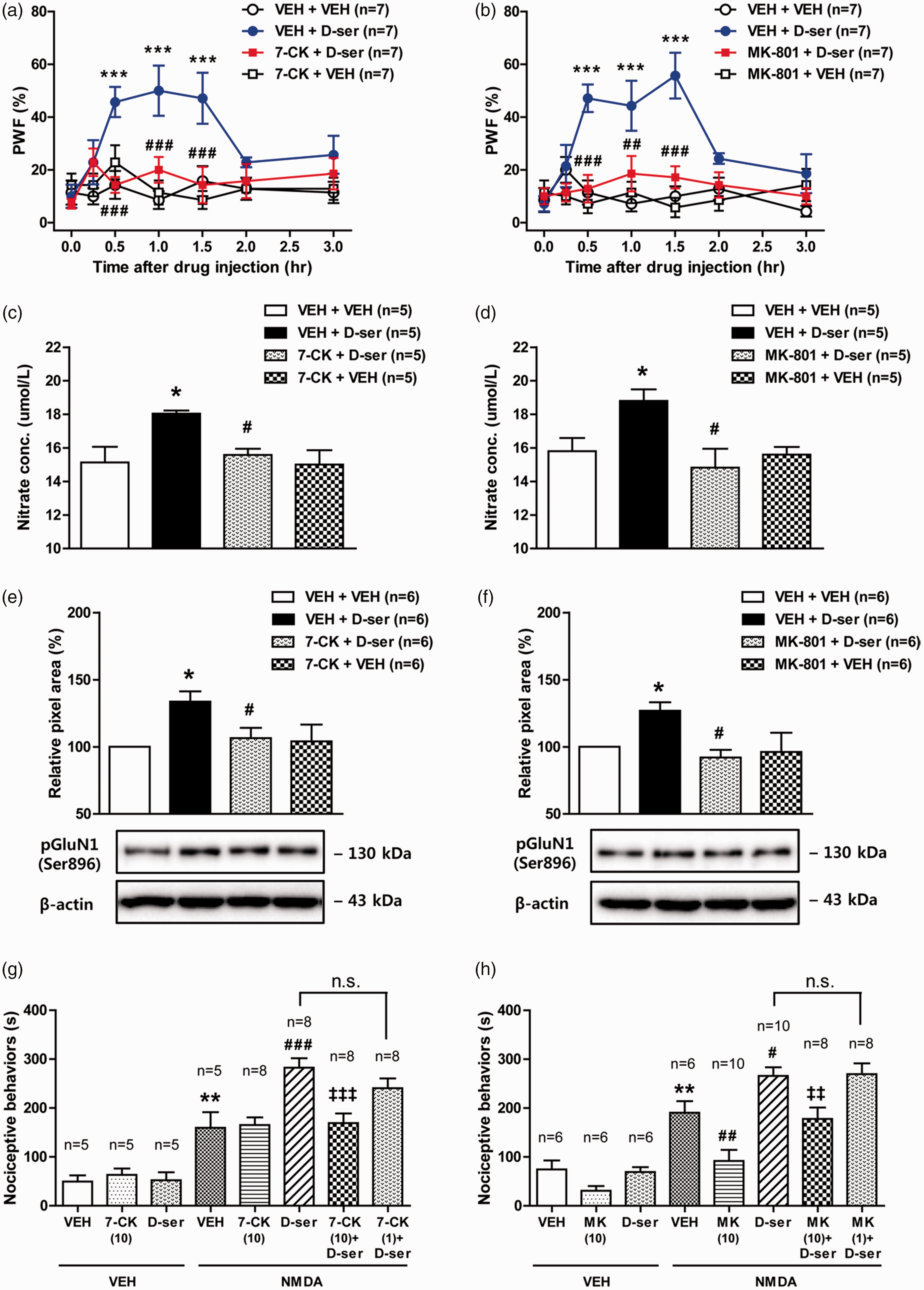
Intrathecal administration of D-serine (500 nmol) significantly increased paw withdrawal frequency (PWF, %) to innocuous mechanical stimuli at 30, 60, and 90 min after D-serine injection as compared with those of the vehicle-treated group (Figure 5(a); ***P < 0.001 vs. VEH + VEH-treated group). Pretreatment with the selective nNOS inhibitor, 7-NI (200 nmol) significantly reduced this D-serine-induced mechanical hypersensitivity (Figure 5(a); #P < 0.05, ###P < 0.001 vs. VEH + D-ser-treated group). In addition, i.t. administration of D-serine significantly increased total NO levels at the 30 min post-injection time point, and this increase was significantly attenuated by pretreatment with 7-NI (200 nmol) (Figure 5(b); *P < 0.05 vs. VEH + VEH-treated group, #P < 0.05 vs. VEH + D-ser-treated group).

Graphs illustrating the effect of i.t. administration of the nNOS inhibitor, 7-NI on the D-serine-induced increases in mechanical hypersensitivity, total NO production, the expression of PKC-dependent (Ser896) pGluN1 and GluN1, and NMDA-induced spontaneous nociceptive behaviors in naïve mice. (a) Intrathecal administration of D-serine (500 nmol) significantly increased PWF (%) to innocuous mechanical stimuli, and this increase was significantly reduced by pretreatment with 7-NI (200 nmol). ***P < 0.001 vs. VEH + VEH-treated group; #P < 0.05, ###P < 0.001 vs. VEH + D-serine-treated group. n = 7 mice/group. (b) Pretreatment with 7-NI (200 nmol) reduced the D-serine (500 nmol)-induced increase in total NO concentration. Intrathecal administration of 7-NI was performed 10 min before D-serine injection and the lumbar spinal cord dorsal horn was sampled 30 min after D-serine injection. *P < 0.05 vs. VEH + VEH-treated group; #P < 0.05 vs. VEH + D-serine-treated group. n = 5 mice/group. (c) Results of Western blot analysis showed that exogenous D-serine increased PKC-dependent (Ser896) pGluN1 and that this increase was significantly attenuated by pretreatment with 7-NI (200 nmol). Exogenous D-serine and 7-NI pretreatment had no effect on total GluN1 expression. *P < 0.05 vs. VEH + VEH-treated group; #P < 0.05 vs. VEH + D-serine-treated group. n = 5–8 mice/group. n.s.: not significant. (d and e) Timepoint of drug(s) administration (d): i.t. administration of NMDA was performed 30 min after D-serine injection. Exogenous D-serine significantly increased NMDA-induced spontaneous nociceptive behaviors (which included caudally directed licking, scratching and biting; (e)). Pretreatment with 7-NI (200 nmol) reduced the D-serine-induced increase in NMDA-induced nociceptive behaviors. *P < 0.05 vs. VEH + VEH-treated group; #P < 0.05 vs. VEH + NMDA-treated group; ‡‡P < 0.01 vs. D-serine + NMDA-treated group. n = 5–10 mice/group. VEH: vehicle; PWF: paw withdrawal frequency; NMDA: N-Methyl-D-aspartate; 7-NI: 7-nitroindazole; D-ser: D-serine; n.s.: not significant; i.t.: intrathecal.
We next investigated whether nNOS activation mediates the exogenous D-serine-induced increase in PKC-dependent (Ser896) pGluN1. To do this, we examined the expression of pGluN1, as well as, the total protein expression of GluN1 using a Western blot analysis. Intrathecal administration of D-serine significantly increased PKC-dependent pGluN1 expression in the spinal cord dorsal horn at the 30-min post-injection time point (Figure 5(c); *P < 0.05 vs. VEH + VEH-treated group). Pretreatment with 7-NI (200 nmol) completely suppressed this D-serine-induced increase in PKC-dependent pGluN1 expression (Figure 5(c); #P < 0.05 vs. VEH + D-ser-treated group). By contrast, exogenous D-serine and 7-NI pretreatment had no effect on total GluN1 expression (Figure 5(c)). In our previous study, we showed that the D-serine-induced increase in GluN1 phosphorylation enhances NMDA-induced spontaneous nociception. 5 We next examined the effect of i.t. pretreatment with the nNOS inhibitor on the D-serine-induced facilitation of NMDA-induced spontaneous nociception. The timeline of drug(s) administration in this experiment is shown in Figure 5(d). Intrathecal administration of 7-NI was performed 10 min before D-serine injection and administration of NMDA was performed 30 min after D-serine injection (this time was selected because it represents a timepoint where there is a significant increase in GluN1 phosphorylation). Intrathecal administration of NMDA (0.4 nmol in 5 µl physiological saline) elicited significant spontaneous nociceptive behaviors including caudally directed licking, scratching, and biting compared to the vehicle-treated group (Figure 5(e); *P < 0.05 vs. VEH + VEH-treated group). Exogenous D-serine administered 30 min prior to NMDA injection significantly increased NMDA-induced nociceptive behaviors (Figure 5(e); #P < 0.05 vs. VEH + NMDA-treated group). This facilitatory effect of D-serine on NMDA-induced nociception was significantly suppressed by pretreatment with 7-NI (200 nmol) in comparison to the vehicle-treated group (Figure 5(e); ‡‡P < 0.01 vs. D-ser + NMDA-treated group). These results suggest that spinal nNOS activation mediates the D-serine-induced increases in mechanical hypersensitivity, total NO levels, and PKC-dependent pGluN1, which leads to the D-serine’s facilitation of NMDA-induced spontaneous nociceptive behaviors.
Effect of i.t. administration of 7-CK or MK-801 on D-serine-induced increases in mechanical hypersensitivity, total NO production, pGluN1 expression, and NMDA-induced spontaneous nociception in naïve mice
To investigate whether D-serine’s effects as illustrated in Figure 5 are mediated by the activation of NMDA receptors, we next examined the effect of NMDA receptor inhibition on the D-serine-induced increases in mechanical hypersensitivity, total NO production, pGluN1 expression, and NMDA-induced spontaneous nociception. Intrathecal administration of D-serine (500 nmol) significantly increased PWF (%) to innocuous mechanical stimuli, and this increase was significantly reduced by the pretreatment with 7-CK (10 nmol, an antagonist of the NMDA receptor glycine binding site) or with MK-801 (10 nmol, an antagonist of the ion channel modulator-binding site of the NMDA receptor) (Figure 6(a) and (b); ***P < 0.001 vs. VEH + VEH-treated group, ##P < 0.01, ###P < 0.001 vs. VEH + D-ser-treated group). In addition, pretreatment with either 7-CK (10 nmol) or MK-801 (10 nmol) significantly reduced the D-serine (500 nmol)-induced increases in spinal NO levels (Figure 6(c) and (d); *P < 0.05 vs. VEH + VEH-treated group, #P < 0.05 vs. VEH + D-ser-treated group).
Graphs illustrating the effect of i.t. administration of the NMDA receptor antagonists, 7-CK and MK-801 on the D-serine-induced increases in mechanical hypersensitivity, total NO production, PKC-dependent (Ser896) pGluN1 expression, and NMDA-induced spontaneous nociceptive behaviors in naïve mice. (a and b) Intrathecal administration of D-serine (500 nmol) significantly increased PWF (%) to innocuous mechanical stimuli, and this increase was significantly reduced by pretreatment with the NMDA receptor glycine antagonist, 7-chlorokynurenic acid ((a); 7-CK, 10 nmol) or the NMDA receptor channel blocker, MK-801 ((b); 10 nmol). ***P < 0.001 vs. VEH + VEH-treated group; ##P < 0.01, ###P < 0.001 vs. VEH + D-serine-treated group. n = 7 mice/group. (c and d) Pretreatment with 7-CK ((c); 10 nmol) or MK-801 ((d); 10 nmol) reduced the D-serine-induced increase in total NO concentration. Intrathecal administration of 7-CK and MK-801 was performed 10 min before D-ser injection and the lumbar spinal cord dorsal horn was sampled 30 min after D-serine- injection. *P < 0.05 vs. VEH + VEH-treated group; #P < 0.05 vs. VEH + D-ser-treated group. n = 5 mice/group. (e and f) Results of Western blot analysis showed that exogenous D-serine- increased PKC-dependent (Ser896) pGluN1 and that this increase was significantly attenuated by pretreatment with 7-CK ((e); 10 nmol) or MK-801 ((f); 10 nmol). *P < 0.05 vs. VEH + VEH-treated group; #P < 0.05 vs. VEH + D-ser-treated group. n = 6 mice/group. (g and h) Exogenous D-serine- significantly increased NMDA-induced spontaneous nociceptive behaviors (which included caudally directed licking, scratching, and biting). Pretreatment with 7-CK ((g); 10 nmol) or MK-801 ((h); 10 nmol) reduced the D-ser-induced increase in NMDA-induced nociceptive behaviors. **P < 0.01 vs. VEH + VEH-treated group; #P < 0.05, ##P < 0.01, ###P < 0.001 vs. VEH + NMDA-treated group; ‡‡P < 0.01, ‡‡‡P < 0.001 vs. D-ser + NMDA-treated group. n = 5–10 mice/group. VEH: vehicle; NMDA: N-Methyl-D-aspartate; D-ser: D-serine; n.s.: not significant; i.t.: intrathecal; PWF: paw withdrawal frequency; pGluN1, phosphorylated GluN1 subunit.
Exogenous D-serine increased the PKC-dependent phosphorylation (Ser896) of the GluN1 subunit in the spinal cord dorsal horn at the 30-min post-injection time point, and this increase was significantly reduced by the pretreatment with 7-CK (10 nmol) or MK-801 (10 nmol) (Figure 6(e) and (f); *P < 0.05 vs. VEH + VEH-treated group, #P < 0.05 vs. VEH + D-ser-treated group). Furthermore, exogenous D-serine administered 30 min prior to NMDA injection significantly enhanced NMDA-induced spontaneous nociceptive behaviors (Figure 6(g) and (h); **P < 0.01 vs. VEH + VEH-treated group, #P < 0.05, ###P < 0.001 vs. VEH + NMDA-treated group). This facilitatory effect of D-serine on NMDA-induced spontaneous nociception was significantly reduced by the pretreatment with either 7-CK (10 nmol) or MK-801 (10 nmol) in comparison to the vehicle-treated group (Figure 6(g) and (h); ‡‡P < 0.01, ‡‡‡P < 0.001 vs. D-ser + NMDA-treated group). These results suggest that D-serine-induced increases in mechanical hypersensitivity, total NO levels, PKC-dependent pGluN1, and enhancement of NMDA-induced spontaneous nociception are mediated by spinal NMDA receptor activation.
Discussion
This study reports three important novel findings. First, i.t. administration of the Srr inhibitor, LSOS or the D-serine degrading enzyme, DAAO significantly reduced total NO levels and nNOS activation (i.e. as evidenced by a significant increase in NADPH-diaphorase staining) in lumbar dorsal horn neurons of CCI mice. In particular D-serine inhibition with LSOS or DAAO administration attenuated the CCI-induced decrease in the ratio of pnNOS (Ser847)/nNOS in the lumbar dorsal horn. Second, co-administration of the NO donor, SIN-1 together with LSOS or DAAO restored both the development of mechanical allodynia and the CCI-induced increase in PKC-dependent (Ser896) pGluN1 expression, which was inhibited by LSOS or DAAO administration alone. Third, exogenous D-serine increased total NO levels and decreased the ratio of pnNOS (Ser847)/nNOS in the lumbar dorsal horn of naïve mice. Pretreatment with the specific nNOS inhibitor, 7-NI or with the NMDA receptor antagonists, 7-CK and MK-801 significantly suppressed the D-serine-induced increases in mechanical hypersensitivity, total NO levels, and pGluN1 expression, which may be one of the major mechanisms associated with D-serine’s facilitative effect on NMDA-induced spontaneous nociception. Collectively, these findings demonstrate that spinal D-serine modulates nNOS activation and that the accompanying increase in NO production is mediated by a reduction in pnNOS and ultimately contributing to the increase in PKC-dependent phosphorylation of NMDA receptor GluN1 subunit and the development of mechanical allodynia in CCI mice.
pnNOS is considered an important mechanism for the regulation of nNOS activation. In particular, phosphorylation at the Ser847 site of nNOS interrupts the binding of the Ca2+/calmodulin to nNOS and thus significantly decreases the activity of this enzyme.11,12 In this regard, a decrease in the phosphorylated forms of nNOS at Ser847 is considered to represent an increase in the active form of the nNOS enzyme. Since there were no changes in the total amount of nNOS protein in the spinal cord dorsal horn of CCI mice at three days post-injury, we evaluated the potential decrease in the ratio of pnNOS (Ser847)/nNOS as an indicator of an increase in nNOS activation. In the present study, D-serine inhibition significantly suppressed total NO concentration and nNOS activation as evidenced by the decrease in the ratio of pnNOS (Ser847)/nNOS. This result is supported in the literature by studies indicating that activation of NMDA receptors modulates spinal nociceptive transmission via nNOS/NO signaling cascades.29,30 In addition, NMDA augments nNOS activity in a concentration-dependent manner and selective D-amino acid degradation has been shown to diminish NMDA-evoked nNOS activity in cerebellar slices. 1 These findings raise the possibility that D-serine-induced activation of NMDA receptors at spinal glutamatergic synapses may be closely linked to nNOS activation and ultimately affect nociceptive transmission. Since we show that NADPH-diaphorase-positive neurons were exclusively located in the SDH region, D-serine could serve as a critical molecular switch that modulates the function of nNOS in SDH neurons contributing to spinal nociceptive signaling transmission.
The NMDA receptor is a tetramer composed of two GluN1 and two GluN2 subunits and has a glycine binding site located on the GluN1 subunit. 31 The receptor activation requires binding of glutamate and a simultaneous binding of the co-agonist D-serine or glycine, which can increase the affinity of glutamate binding and facilitate excitatory transmission. 32 Difference in sensitivity to glycine/D-serine and glutamate has been reported for receptors containing different GluN2 subunits. Paudice et al. demonstrated that in the absence of NMDA, glycine or D-serine alone can activate NMDA receptors, which may display the triple subunit combination GluN1/GluN2A/GluN2B, leading to the release of cholecystokinin and somatostatin from rat neocortical nerve endings. 33 Antagonists selective for the glycine site or for the NMDA receptor channel prevented the effects of glycine/D-serine; these effects were, however, insensitive to blockade of the glutamate recognition site of the NMDA receptor, suggesting that glutamate released from synaptosomes was not involved. Furthermore, glycine/D‐serine might have elicited endogenous glutamate/aspartate release. 33 In the present study, exogenous D-serine increased mechanical hypersensitivity, NO concentration, and GluN1 phosphorylation, and these increases were significantly suppressed by the glycine binding site antagonist, 7-CK and the NMDA receptor channel blocker, MK-801. These results suggest the possibility that i.t. administration of D-serine alone can activate some subtypes of NMDA receptors leading to the sensitization of nociceptive signaling under our experimental conditions. In addition, Papouin et al. demonstrated that D-serine is the co-agonist at synaptic NMDA receptors in the CA1 region of the hippocampus, whereas glycine serves as the major co-agonist at extrasynaptic NMDA receptors suggesting that functional compartmentalization arises from glycine and D-serine bioavailability. 34 Thus, different NMDA receptors can be activated depending on whether D-serine is present in higher amounts versus low levels. We plan to investigate the mechanisms underlying this process in more detail in future studies.
Spinal NMDA receptor phosphorylation is closely related to the processing of nociceptive transmission in the spinal cord dorsal horn. In this regard, Ultenius et al. demonstrated that phosphorylation of the spinal NMDA receptor GluN1 subunit is correlated with the presence of behavioral signs of neuropathic pain (stimulus evoked pain-like behavior) following partial sciatic nerve ligation in the rat. 35 A study by Gao et al. also provided evidence that the increase of GluN1 phosphorylation in the ipsilateral dorsal horn is closely correlated to the development of mechanical allodynia represented by hypersensitivity to innocuous mechanical stimuli after L5 spinal nerve ligation. 36 Moreover, approximately one-third of spinal pGluN1-immunoreactive neurons were identified as projection neurons that convey primary afferent inputs to supraspinal levels. 36 These results are consistent with our present data suggesting that PKC-dependent GluN1 phosphorylation is closely related to peripheral nerve injury-induced chronic neuropathic pain and in particular to the development of mechanical allodynia. In addition, i.t. administration of D-serine increases PKC-dependent GluN1 phosphorylation resulting in mechanical hypersensitivity and a significant increase in NMDA-induced nociceptive behaviors. It has been suggested that PKC-dependent pGluN1 at the Ser896 site results in trafficking of GluN1 to the cell membrane and potentiation of NMDA receptor function by removing the Mg2+ block and increasing the probability of channel openings.37,38 Thus, phosphorylation of NMDA receptors during the induction phase of neuropathic pain or following administration of D-serine increases neuronal excitability in postsynaptic neurons contributing to both the enhancement of synaptic efficacy and the development of central sensitization. Since both D-serine production and the expression of its synthesizing enzyme, Srr only increase during the early phase of neuropathic pain, 4 we speculate that there are other regulating mechanisms responsible for the maintenance of central sensitization that is sustained by NMDA receptor activation during the late phase of chronic pain.
In the present study, nNOS/NO was shown to mediate the D-serine-induced increase in PKC-dependent pGluN1 expression in the spinal cord dorsal horn. Several reports have suggested that NO donors selectively activate the PKCα isoform in kidney cells and the PKCε isoform in the heart.39,40 PKCα is a conventional subtype, which is Ca2+- and diacylglycerol (DAG)-dependent, while PKCε is a novel subtype, which is Ca2+-independent and DAG-dependent. 41 Wright et al. reported that NO can modulate the activation of phospholipase C, an enzyme that synthesizes both DAG and IP3, in response to external oxidant stress, 42 and thus, it is plausible that PKCα and/or PKCε may be regulated by NO indirectly. Furthermore, there is converging evidence from several laboratories that the PKCγ isoform is primarily associated with the production of mechanical allodynia after peripheral nerve injury.43,44 Since PKCγ immunoreactivity is confined to interneurons of the inner part of lamina II,43,44 where the positive NADPH-diaphorase staining exists, it is plausible that nNOS-induced NO modulates the PKCγ isoform regulating the integration process of noxious and non-noxious inputs to dorsal horn neurons, which ultimately contributes to the development of mechanical allodynia. While data clearly support this hypothesis, further research is needed to investigate the underlying mechanisms of NO-induced PKC activation. Since the total protein expression of GluN1 was not affected by either D-serine or nNOS activation, post-translational modification via phosphorylation of GluN1 appears to be regulated by different mechanisms in comparison to the gene transcriptional changes in total protein expression of GluN1.
In the present study, administration of LSOS and DAAO as well as co-administration of these drugs with SIN-1 significantly modulated the development of mechanical allodynia in neuropathic mice. On the other hand, these treatments had no effect on the development of thermal hyperalgesia in CCI animals. In light of these results, we would suggest that D-serine and nNOS signaling in the spinal cord plays an important role in the development of mechanical allodynia rather than thermal hyperalgesia following sciatic nerve injury. This is consistent with several reports suggesting that different mechanisms are involved in the development of mechanical allodynia versus thermal hyperalgesia in animal models of neuropathic pain.45,46 In contrast, while several studies have demonstrated that the inhibition of NO signaling or blockade of NMDA receptors significantly attenuates both mechanical and thermal hyperalgesia in animal models of pain,8,29,47 this did not occur in our study. This discrepancy may be due to: (1) the differences in the animal models being examined; (2) the different time course of nociception in each model; (3) the different behavioral assays used to evaluate nociception; and/or (4) the nervous system location being examined. These differences require further investigation to clarify the mechanisms underlying the development and maintenance of mechanical allodynia and thermal hyperalgesia in different models of pain.
Following peripheral nerve injury, the prolonged activity of nociceptive C-fibers release a diverse array of neurotransmitters, including substance P, glutamate, and calcitonin gene-related peptide, each of which binds to its own receptors on adjacent astrocytes and second order neurons. 48 Activation of NMDA receptors by glutamate and by D-serine released from neighboring activated astrocytes induces an extracellular Ca2+ influx into neurons that triggers a cascade of events, including activation of nNOS through dephosphorylation of nNOS (Ser847) catalyzed by calcineurin, leading to an increased production of NO.1,30,49 NO can diffuse out of the neuron, acting on other neurons, astrocytes and microglial cells located as some distance from the neuron of origin, but can also diffuse across the synaptic cleft back into the synapse that originally released glutamate. 30 In addition, it has been suggested that spinal NO participates in glutamatergic mechanisms of descending facilitation triggered by CNS inflammation, weakening the influence of descending inhibition upon dorsal horn neurons by interfering with GABAergic and glycinergic inhibitory tone. 7 Furthermore, in the present study, we show that NO increases PKC-dependent pGluN1. This action of NO further increases neuronal excitability, 50 eventually contributing to the induction of “central sensitization” (see the schematic diagram illustrated in Figure 7). Collectively, this series of events may cause normally non-painful stimuli from mechanoreceptive Aβ-fibers to be perceived as pain, 51 which is one of the hallmarks of mechanical allodynia development. On the other hand, experimental and clinical evidence have demonstrated that NO is also able to induce analgesia.7,9,10 In this regard, further studies are needed to investigate the control mechanisms underlying NO-related pain transmission and to support the clinical use of NO or NOS inhibitors in pain therapy.

Schematic diagram depicting the proposed mechanisms underlying CCI-induced central sensitization and the development of mechanical allodynia. Peripheral nerve injury induces abnormal nociceptive signaling resulting in the subsequent release of mediators from primary afferents. D-serine can be produced by serine racemase activation in spinal astrocytes. D-serine released from astrocytes binds to the NMDA receptor glycine site and activates NMDA receptors with glutamate leading to increased Ca2+ influx into postsynaptic neurons. Ca2+/calmodulin-dependent activation of calcineurin then catalyzes the dephosphorylation of nNOS at the Ser847 site resulting in an increase in NO production. These NO signaling cascades are likely to stimulate PKC, which in turn induces phosphorylation of the PKC-dependent NMDA receptor GluN1 subunit at the Ser896 site, resulting in the development of central sensitization and ultimately leading to the development of mechanical allodynia associated with neuropathic pain. DAAO: D-amino acid oxidase; LSOS: L-serine O-sulfate potassium salt; NO: nitric oxide; PKC: protein kinase C; NMDAR: N-Methyl-D-aspartate receptor.
In conclusion, the present study demonstrates that astrocyte D-serine, which serves as a co-agonist for the glycine site on NMDA receptors, increases NO production and nNOS activation by reducing the pnNOS in lumbar spinal cord dorsal horn neurons. Moreover, nNOS activation also appears to mediate D-serine’s effects on PKC-dependent phosphorylation of the NMDA receptor GluN1 subunit and the development of CCI-induced mechanical allodynia. Collectively, these findings suggest that the increase in NO signaling that occurs via D-serine-induced NMDA receptor activation plays a critical role in spinal D-serine-induced central sensitization and ultimately contributes to the development of neuropathic pain.
Footnotes
Declaration of Conflicts of Interest
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was supported by the National Research Foundation of Korea grant funded by the Korean Government (NRF-2017R1A2A2A05001402).