
Abstract
Background
Chronic pain has been shown to depend on nociceptive sensitization in the spinal cord, and while multiple mechanisms involved in the initiation of plastic changes have been established, the molecular targets which maintain spinal nociceptive sensitization are still largely unknown. Building upon the established neurobiology underlying the maintenance of long-term potentiation in the hippocampus, this present study investigated the contributions of spinal atypical protein kinase C (PKC) isoforms PKCι/λ and PKMζ and their downstream targets (p62/GluA1 and NSF/GluA2 interactions, respectively) to the maintenance of spinal nociceptive sensitization in male and female rats.
Results
Pharmacological inhibition of atypical PKCs by ZIP reversed established allodynia produced by repeated intramuscular acidic saline injections in male animals only, replicating previously demonstrated sex differences. Inhibition of both PKCι/λ and downstream substrates p62/GluA1 resulted in male-specific reversals of intramuscular acidic saline-induced allodynia, while female animals continued to display allodynia. Inhibition of NSF/GluA2, the downstream target to PKMζ, reversed allodynia induced by intramuscular acidic saline in both sexes. Neither PKCι/λ, p62/GluA1 or NSF/GluA2 inhibition had any effect on formalin response for either sex.
Conclusion
This study provides novel behavioural evidence for the male-specific role of PKCι/λ and downstream target p62/GluA1, highlighting the potential influence of ongoing afferent input. The sexually divergent pathways underlying persistent pain are shown here to converge at the interaction between NSF and the GluA2 subunit of the AMPA receptor. Although this interaction is thought to be downstream of PKMζ in males, these findings and previous work suggest that females may rely on a factor independent of atypical PKCs for the maintenance of spinal nociceptive sensitization.
Keywords
Introduction
The maintenance of chronic pain in spinal cord dorsal horn (SCDH) and memory within the hippocampus share similar underlying molecular mechanisms, including distinct early and late phases underlying the initiation and maintenance of the plastic changes.1–3 Although multiple signalling molecules (PKC, CAMKII, and ERK1,2) involved in the early initiation of spinal nociceptive sensitivity have been established,4–6 the molecular targets that maintain this hypersensitivity remain largely unclear. How nociceptive sensitivity is maintained is thought to contribute to the transition from acute to chronic, persistent pain. 2 The established neurobiology of long-term potentiation (LTP) maintenance in the hippocampus may be used as a guide to identify targets involved with the maintenance of plasticity in the SCDH; a critical area of study for understanding the persistence of a centralized pain state and pain chronicity.3,7
Protein kinase M zeta (PKMζ), an autonomously active atypical PKC isoform, has been implicated in the maintenance of hippocampal LTP and memory,8–11 as well as persistent pain states.3,12–16 PKMζ is increased in the SCDH for a prolonged period following intraplantar (i.pl.) injection of formalin or capsaicin, as well as direct stimulation of SCDH nociceptive neurons by spinal injection of the glutamate agonist dihydroxyphenylglycine.12,16 Behavioural and electrophysiological findings supporting the maintenance role of PKMζ have been largely generated using the cell permeable zeta pseudosubstrate inhibitory peptide (ZIP), a decoy peptide that mimics PKMζ’s missing regulatory region that would normally reduce the autonomous activity of the kinase.3,8–16
Notably, the effects of spinal PKMζ inhibition using ZIP appear to be demonstrated only in pain tests which depend primarily on central sensitization. There has been a demonstrated lack of effect of ZIP in pain tests known to be dependent on nociceptor sensitization, 12 or those with high ongoing peripheral input, for which reducing central sensitization may not be enough to reduce pain behaviours.3,16–18 As such, hyperalgesic priming, in which an ordinarily sub-threshold challenging stimulus is delivered following the acute sensitization from an initial priming stimulus,19,20 has often been used to isolate the centrally-mediated effects of ZIP.3,13–15
The specificity of ZIP, and consequently, the role of PKMζ, has been recently questioned following genetic ablation studies in the hippocampus.22,23 Constitutive PKMζ (PRKCZ) knockout mice reported no deficits in late-LTP or hippocampal-dependent learning and memory, and administration of ZIP to the PRKCZ knockout mice still reversed late-LTP, learning and memory; suggesting non-specific effects of the peptide inhibitor.22,23 ZIP is a myristoylated autoinhibitory pseudosubstrate amino acid sequence found in the regulatory domains of all atypical PKC isoforms: PKCζ, PKMζ and PKCι/λ. As such, ZIP has been shown to inhibit additional isoforms in in vitro assays, 21 as well as in vivo,23,24 and it was proposed that these may be the target of ZIP, responsible for the maintenance of plasticity. 23 However, conditional knockout of PRKCZ identified a compensatory maintenance mechanism for LTP by PKCι/λ; proposing that the permanent deletion of the PRKCZ gene through constitutive knockout resulted in a functional redundancy in atypical PKC signalling pathways. 24 These authors showed that a selective PKCι/λ-antagonist (ICAP) disrupts late-LTP and spatial memory in PKMζ-null mice, but not in wild-type mice, 24 proposing that PKCι/λ compensates for the action of PKMζ, only if it has been deleted. 24 Moreover, the distinct atypical PKC pathways mediating AMPA receptor (AMPAR) trafficking in the early and late phases of synaptic potentiation have also been shown to undergo a mechanistic switch following conditional PKCι/λ knockout studies, switching between the established early PKCι/λ-p62/GluA1-driven AMPAR exocytosis and late PKMζ-NSF/GluA2-driven reduction in AMPAR endocytosis,25,26 further demonstrating atypical PKC compensation to maintain memory function.
In contrast to the hippocampal findings, constitutive knockout of PRKCZ was shown to have consistent effects with spinal administration of ZIP in models of neuropathic, inflammatory, and referred muscle and visceral pain. 3 However, knowledge of the mechanisms maintaining spinal nociceptive sensitization is far behind that of hippocampal LTP. While it has been established that the atypical isoform PKCζ does not maintain persistent pain states, 12 there has been little investigation into the contribution of PKCι/λ or the potential downstream targets of either PKCι/λ or PKMζ (p62/GluA1 and NSF/GluA2, respectively). Moreover, nearly all research on PKMζ and pain has solely used male animals, with the exception of one study. 3 Nasir et al. 3 demonstrated that both spinal ZIP administration and constitutive PRKCZ ablation reversed allodynia produced by repeated injections of intracolonic capsaicin or intramuscular (i.m.) acidic saline in male rodents, but neither showed a reduction in the persistent allodynia developed in females following the same treatment. 3 This sexual divergence underlying the same painful endpoint suggests that females likely rely on mechanisms other than PKMζ for the maintenance of spinal nociceptive sensitization. Given the demonstrated ability of PKCι/λ to maintain late-LTP in PKMζ-null mice, 24 it is possible that PKCι/λ, rather than PKMζ, may actively maintain the spinal nociceptive plasticity underlying persistent pain in females.2,7
Given the above outlined issues with the specificity of the aPKC inhibitor ZIP, the lack of research on downstream targets of aPKCs and sex differences in the contribution of aPKCs to the maintenance of spinal nociceptive sensitization, we performed the following three experiments which assessed nociceptive behaviours in rat models of persistent pain: (1) the non-specific pharmacological manipulation of atypical PKCs, (2) the specific pharmacological manipulation of PKCι/λ and downstream target p62/GluA1, and (3) the specific pharmacological manipulation of downstream targets of PKMζ, NSF/GluA2.
Materials and methods
Animals
All procedures performed on rats were approved by the McGill University Animal Care Committee and followed the ethical guidelines of the Canadian Council on Animal Care and the International Association for the Study of Pain.
Female and male Long-Evans hooded rats (eight weeks of age) were supplied by Charles River Laboratories (Saint-Constant, QC), housed locally, and kept on a 12-h dark-light cycle with food and water available ad libitum. Prior to the beginning of experiments, naïve rats were acclimatized to the animal care facility for a period of two to five days and then habituated to the testing apparatus for 30-min per day for three consecutive days.
Drug administration
All drugs were administered in a volume of 20 µL into the intrathecal (i.t.) space by lumbar puncture. Rats were anesthetized with isoflurane (4% induction, 2% maintenance), and a 26-gauge needle was inserted between the L5 and L6 vertebrae into the cauda equina; correct placement was verified by a positive reflexive tail flick at initial needle placement and following drug injection. The following drugs were used: ZIP (Myr-SIYRRGARRWRKL-OH, Tocris), scr-ZIP (Myr-RLYRKRIWRSAGR-OH, Tocris Bioscience, Oakville, ON) both dissolved in sterile water at a dose of 10 nmol, 12 ICAP, [4-(5-amino-4-carba-moylimidazol-1-yl)-2,3-dihydroxycyclopentyl] methyl dihydrogen (graciously received from Dr. Robert Farese) dissolved in sterile water at a dose of 20 nmol, Tat-GluA1(L2-3) (YGRKKKRRQRRR-DISPRSLSGR), scr-Tat-GluA1(L2-3) (YGRKKKRRQRRR-IFNSDGAFMF) dissolved in sterile water at a dose of 10 µg 23 (Tat-GluA1(L2-3) and scr-Tat-GluA1(L2-3) were synthesized by CanPeptide Inc, Pointe-Claire, QC), myr-pep2m (Myr- KRMKVAKNAQ, Tocris) and myr-pep4c (Myr- KRMKVAKSAQ, Tocris), dissolved in sterile water at a dose of 10 µg. 13 I.t. doses were selected from previously reported effective doses or obtained from preliminary studies to assess potential toxicity and/or behavioural impairments.
Von Frey testing
Animals were tested in acrylic test chambers (12 cm W
I.m. acidic saline
Two i.m. injections of 100 µL of acidic saline (pH adjusted to 4.0) spaced five days apart were administered to the gastrocnemius muscle of naïve rats under brief isoflurane anesthesia (induction 3.5%, maintenance 2%) to produce persistent hind paw mechanical allodynia lasting at least four to five weeks. 28 A chart outlining the experimental method is shown in Figure 1(a).
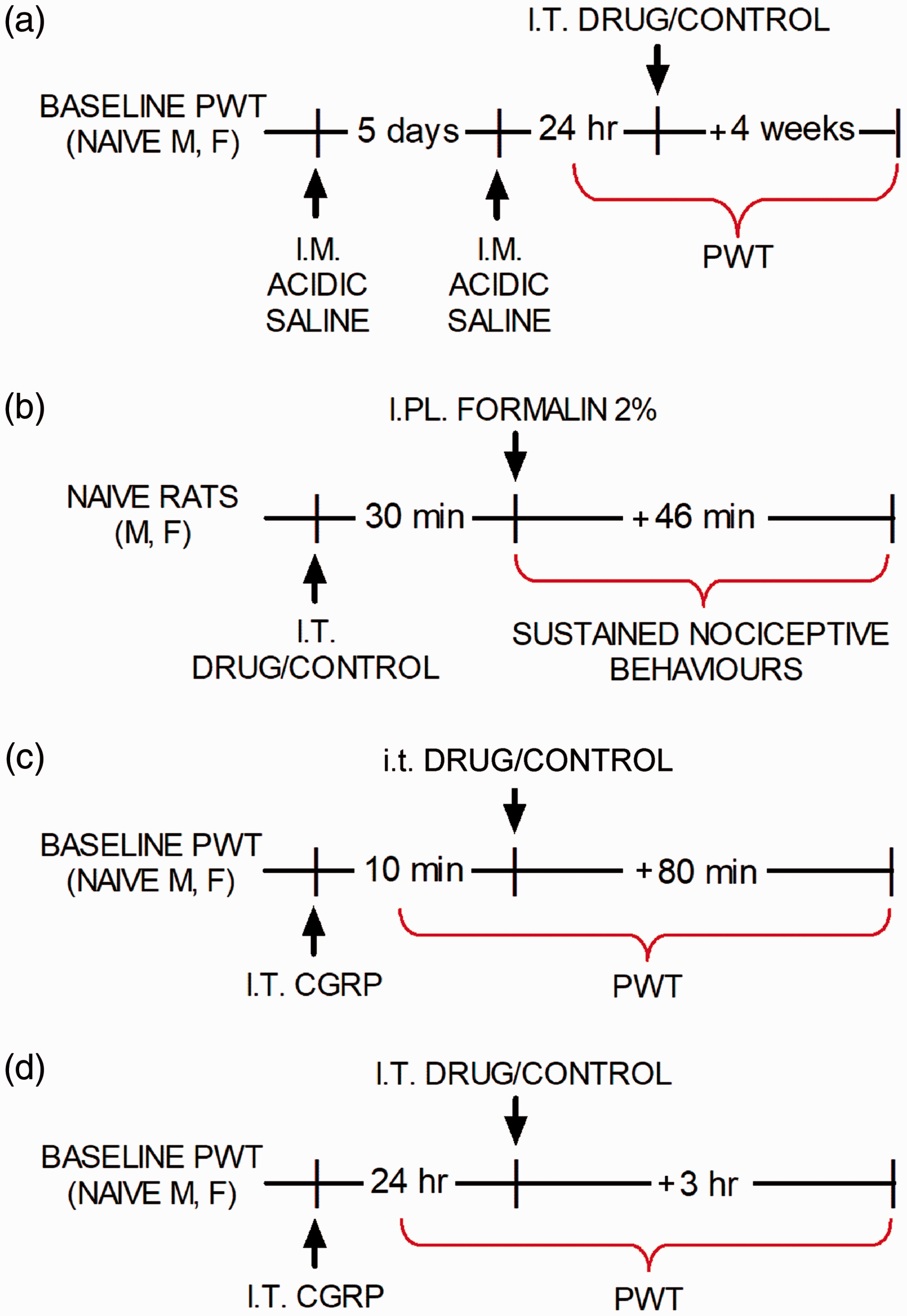
Experimental design of studies using: (a) intramuscular acidic saline, (b) formalin test, (c) acute intrathecal CGRP and (d) persistent intrathecal CGRP.
Formalin test
Rats were tested in acrylic observation boxes (30 cm W
I.t. calcitonin gene-related peptide
Acute mechanical allodynia was induced by i.t. injection of calcitonin gene-related peptide (CGRP; 0.5 µM, in a volume of 20 µL) and assessed by von Frey testing first at 10 min and then every 20 min up to 90 min following the injection (see Figure 1(c)). Long-term CGRP-induced allodynia was produced by i.t. injection of a higher dosage (5 µM, 20 µL volume) and assessed by von Frey testing 24 h after injection (see Figure 1(d)). 30
Data analysis
All data are presented graphically (Prism 5.0, Graphpad Software, La Jolla, CA) as mean
Results
Effects of ZIP on allodynia following i.m. acidic saline
We first examined the role of spinal atypical PKCs (PKMζ, PKCζ, PKCι/λ) in the maintenance of nociceptive sensitization by assessing the post-treatment effects of the non-specific atypical PKC inhibitor ZIP on allodynia produced by repeated i.m. acidic saline injections. Two i.m. injections of acidic saline significantly decreased ipsilateral PWTs from baseline measures in male and female rats, establishing mechanical allodynia in both sexes. There were no differences between males and females in PWTs either before (baseline) or after acidic saline treatment, prior to ZIP administration (Tukey tests, p > 0.05). After treatment, a significant Time

Effect of ZIP (atypical PKC inhibition) on mechanical allodynia after i.m. acidic saline in (a) males and (b) females (n=6/group). Asterisks indicate differences in PWTs between rats treated with ZIP and scrambled control peptide (**p<0.01, ***p<0.001, Tukey test). Daggers indicate differences in PWTs compared to baseline measures for ZIP-treated animals (†††p<0.001, Dunnett test), while double daggers indicate difference from baseline in Scr-ZIP-treated animals (‡‡‡p<0.001, Dunnett test).
Effects of ICAP in the formalin test and following i.m. acidic saline
We then examined the specific contributions of the atypical PKC isoform PKCι/λ to spinal nociceptive processing in the formalin test. PKCι/λ was inhibited by the cell permeable specific PKCι/λ antagonist ICAP
24
administered by i.t. injection 25 min prior to formalin. The formalin test produced typical biphasic nociceptive responses over time in both female and male rats. Pre-treatment spinal inhibition of PKCι/λ by ICAP did not elicit significantly different nociceptive responses in the formalin test compared to the behaviours produced by pre-treatment of the vehicle control in either male (Figure 3(a)) or female rats (Figure 3(b)), and thus no sex differences occurred.
Effect of ICAP (PKCι/λ inhibition) on nociceptive responses in the formalin test in (a) males (ICAP n=8; VEH n= 7) and (b) females (ICAP n=8; VEH n=6) or on mechanical allodynia after i.m. acidic saline in (c) males and (d) females (n=6/group). Asterisks indicate significant differences in responses between ICAP- and vehicle control-treated rats (*p<0.05, ***p< 0.001, Tukey test). Daggers indicate differences in PWTs compared to baseline measures for ICAP-treated animals (†p<0.05, †††p<0.001, Dunnett test), while double daggers indicate difference from baseline in vehicle-treated animals (‡‡‡p<0.001, Dunnett test).
We next examined the effects of the ICAP on allodynia produced by i.m. acidic saline. Mechanical allodynia was established in both sexes by the i.m. acidic saline injections, with PWTs significantly reduced from baseline levels prior to ICAP treatment. There were no sex-related differences in PWTs either at baseline or before drug treatment (Tukey test, p > 0.05). Analysis of PWTs identified a significant interaction of Sex
Effects of Tat-GluA1(L2-3) in the formalin test and following i.m. acidic saline
We next examined the potential downstream targets of spinal PKCι/λ to nociceptive processing by disrupting the interaction between scaffolding protein p62 and the GluA1 subunit of AMPAR, targets shown to be downstream of PKCι/λ in the hippocampus.
23
The p62/GluA1 interaction was disrupted by the spinal administration of cell-permeable peptide Tat-GluA1(L2-3).
23
There was no significant change in the biphasic pattern of nociceptive behaviours produced by hind paw formalin injection in either male or female animals pre-treated with Tat-GluA1(L2-3) compared to formalin-induced nociceptive behaviours that occurred following pre-treatment with scrambled-Tat-GluA1(L2-3) (Figure 4(a) and (b)). Thus, no sex differences were found after the inhibition of interactions of p62/GluA1 by Tat-GluA1(L2-3) in the formalin test.
Effect of Tat-GluA1(L2-3) (p62/GluA1 inhibition) as compared to scrambled peptide on nociceptive responses in the formalin test in (a) male and (b) female rats, or on mechanical allodynia after i.m. acidic saline in (c) males and (d) females (n=6/group). Asterisks indicate significant differences between PWTs following Tat-GluA1(L2-3) and scrambled control peptide treatment (**p<0.01, Tukey test). Daggers indicate differences in PWTs compared to baseline measures for Tat-GluA1(L2-3)-treated animals (†p<0.05, ††p<0.01, †††p<0.001, Dunnett test), while double daggers indicate difference from baseline in Scr-Tat-GluA1(L2-3)-treated animals (‡‡‡p<0.001, Dunnett test).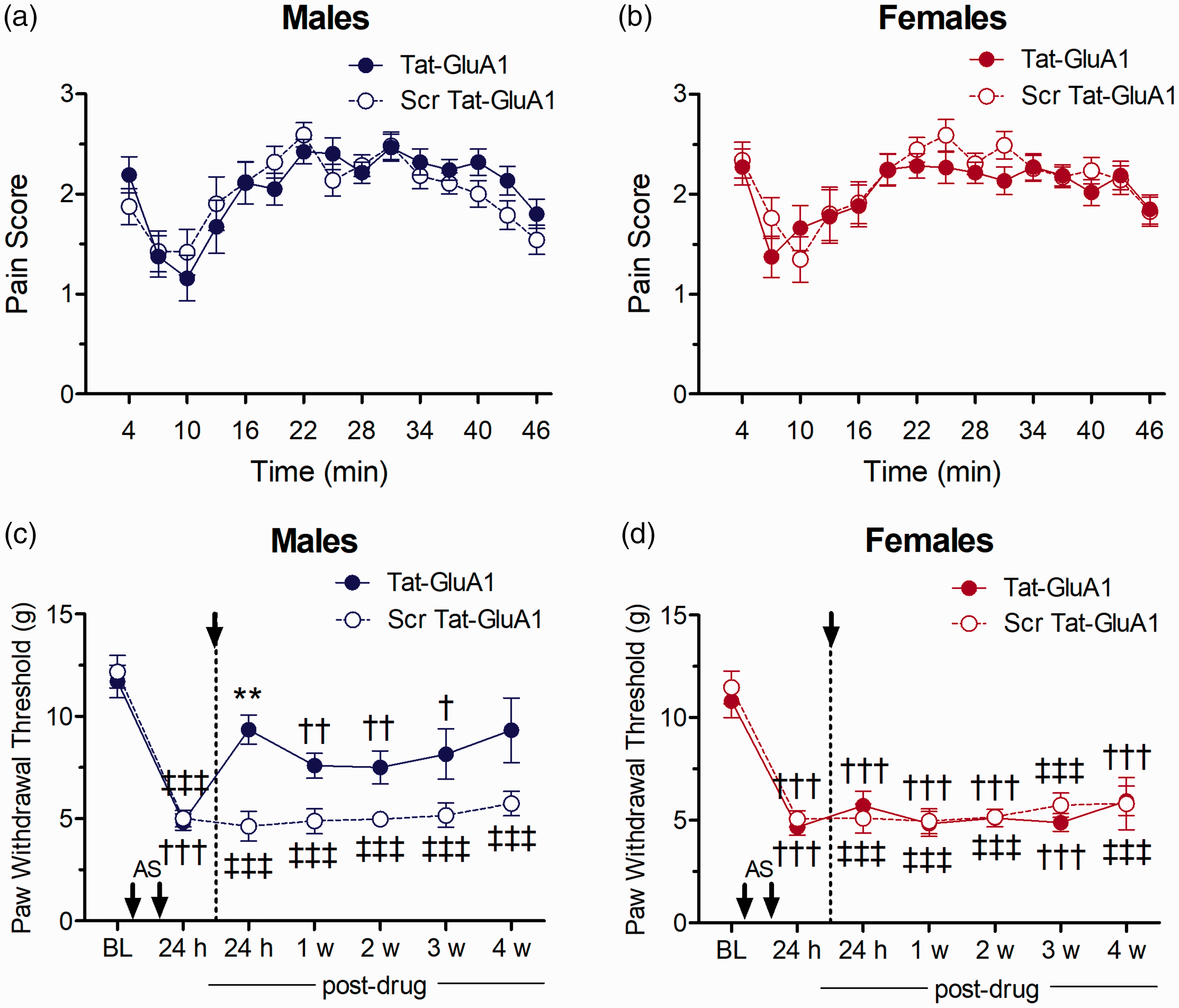
We next assessed the effects of Tat-GluA1(L2-3) and its control peptide on allodynia in i.m. acidic saline-treated rats. Mechanical allodynia was established 24 h after the second acidic saline injection for both sexes, with a significant reduction in PWTs compared to baseline measures. There were no sex-related differences in PWTs either at baseline or before drug treatment (Tukey test, p > 0.05). Post-treatment of spinal Tat-GluA1(L2-3) resulted in an increase in mean PWTs in males, which were significantly higher than PWTs for rats treated with scr-Tat-GluA1(L2-3) at 24 h after i.t. drug administration (p = 0.0117; Figure 4(c)). In contrast, PWTs in females remained significantly reduced from baseline following treatment with Tat-GluA1(L2-3) and did not differ significantly from those in rats treated with scr-Tat-GluA1(L2-3) at any time point (Figure 4(d)), giving rise to a significant interaction of Sex
Effects of myr-pep2m in the formalin test and following i.m. acidic saline
We next examined the downstream targets of spinal PKMζ on nociceptive processing by disrupting the interaction between NSF and the GluA2 subunit of the AMPAR. Previous work, using only male animals, showed that spinal inhibition of NSF/GluA2 prevented the development of mechanical allodynia in a hyperalgesic priming test using interleukin-6/prostaglandin E2 (IL6/PGE2). 13 The NSF/GluA2 interaction was disrupted by the cell-permeable peptide myr-pep2m and compared to effects of the inactive control peptide, myr-pep4c. 13 In the formalin test, pre-treatment with i.t. myr-pep2m did not significantly alter the biphasic pattern of nociceptive behaviours compared with pre-treatment myr-pep4c, in either sex (Figure 5(a) and (b)).

Effect of myr-pep2m (NSF/GluA2 inhibition) on nociceptive responses in the formalin test in (a) males and (b) female rats, or on mechanical allodynia following i.m. acidic saline in (c) males and (d) females (n=6/group). Asterisks indicate significant differences between myr-pep2m- and scrambled control peptide-treated rats (*p<0.05, **p<0.01, ***p<0.001, Tukey test). Daggers indicate differences in PWTs compared to baseline measures for myr-pep2m-treated animals (†††p<0.001, Dunnett test), while double daggers indicate difference from baseline in myr-pep4c-treated animals (‡‡‡p<0.001, Dunnett test).
The role of spinal PKMζ substrate NSF/GluA2 was then assessed on the established allodynia produced by i.m. acidic saline, using the inhibitor myr-pep2m. The repeated i.m. injections of acidic saline produced a significant reduction in PWTs from baseline measures in both female and male rats, establishing mechanical allodynia for both sexes. There were no sex differences in PWTs either at baseline or before drug treatment (Tukey test, p > 0.05). Post-treatment spinal administration with myr-pep2m resulted in an increase in PWTs, in both male and female rats at various time points tested (Time
NSF/GluA2 disruption following i.t. CGRP
We assessed whether female-specific atypical PKC mechanisms may be driven by CGRP, as spinal administration of a CGRP receptor agonist was previously shown to reverse mechanical allodynia produced by hyperalgesic priming in females, but not males. 31 I.t. administration of low dose CGRP 30 resulted in a robust acute allodynic effect, with a significant reduction in PWTs from baseline lasting for 90 min following injection in both males and females. Spinally administered myr-pep2m delivered after CGRP-induced allodynia was established did not elevate PWTs across the 90-min testing period for either sex, with no significant differences between the PWTs in female or male rats compared to thresholds in rats treated with the inactive vehicle control. Thus, no sex differences were found for the effects of inhibition of NSF/GluA2 on acute CGRP-induced allodynia (Figure 6(a) and (b)).
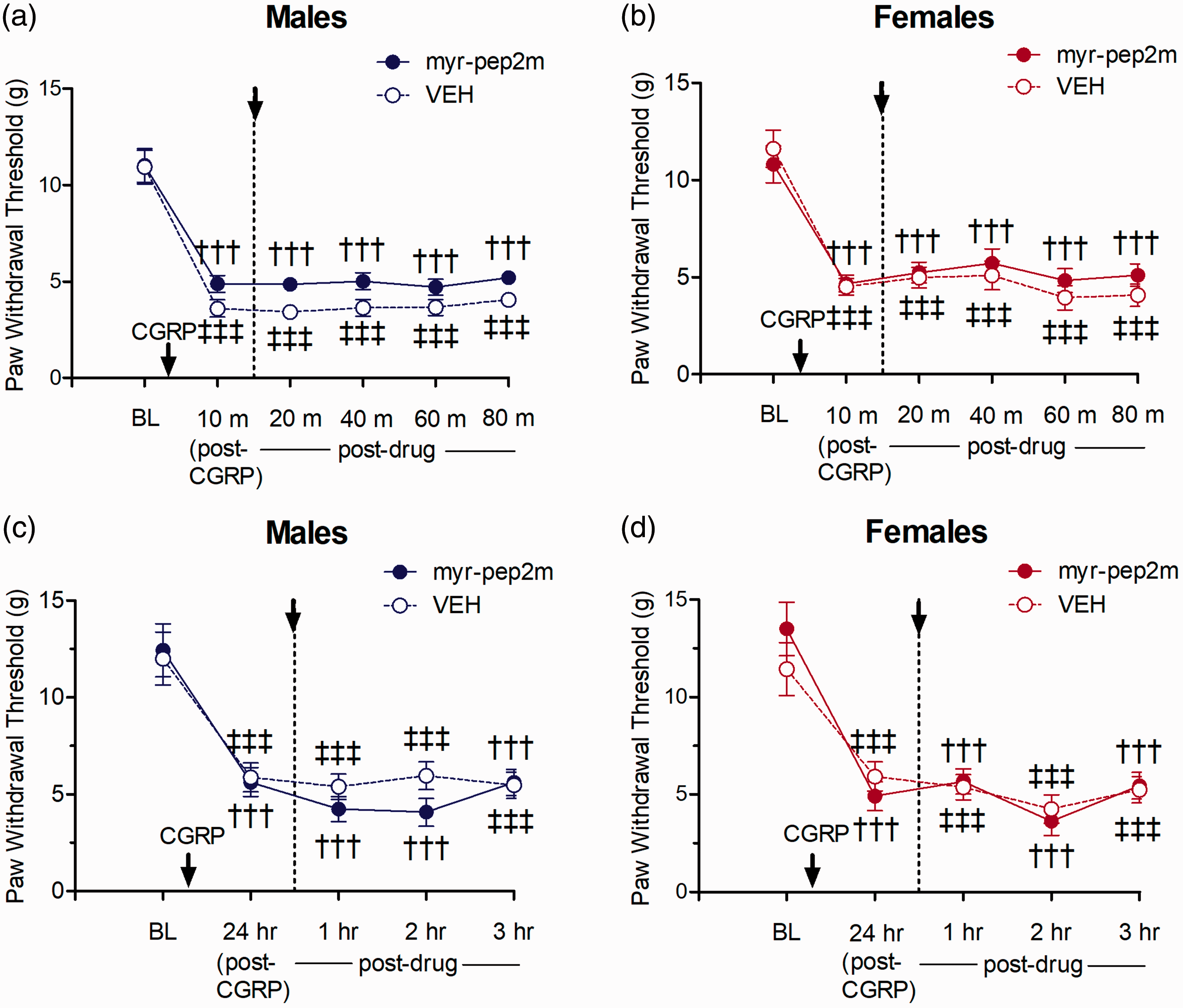
Effect of myr-pep2m (NSF/GluA2 inhibition) on acute mechanical allodynia induced following i.t. CGRP in (a) males and (b) females, or persistent mechanical allodynia induced 24 h following high-dose i.t. CGRP in (c) males and (d) females (n=6/group). Paw withdrawal thresholds did not differ between myr-pep2m- and vehicle control-treated female or male rats for either test.

Male and female responses to each drug following intramuscular acidic saline were compared by analyzing the area under the time-effect curve (AUC). There was a significant Sex
A higher dose of i.t. CGRP produced a long-lasting allodynia after administration, with PWTs significantly reduced compared to baseline measures for >24 h. Post-treatment of spinal myr-pep2m again did not increase the PWTs compared to vehicle control, for either males or females. Thus, no sex differences were found for the inhibition of interactions of NSF/GluA2 on persistent CGRP-induced allodynia (Figure 6(c) and (d)).
Sex differences in drug effects on i.m. acidic saline-induced allodynia
For experiments assessing intramuscular acetic saline-induced allodynia, we also calculated AUCs from the time course data for each drug treatment (ZIP, ICAP, Tat-GluA1(L2-3), myr-pep2m) to directly compare the responses of males and females to inhibition of aPKCs, PKCι λ, p62/GluA1 and NSF/GluA2. Comparison of male and female responses to ZIP, ICAP and Tat-GluA1(2L-3) indicated robust sex differences (p = 0.00013, p = 0.00014, p = 0.002, respectively), while there were no sex-related effects of interactions on allodynia detected after inhibition of NSF/GluA2 with myr-pep2m (p = 0.9871) (Figure 7).
Discussion
These findings provide novel insights into the contributions of atypical PKC isoforms and their downstream targets in the maintenance of hypersensitivity underlying persistent pain, as well as identify critical sex differences in the mechanisms underlying spinal nociceptive sensitization. Nearly all previous work related to PKMζ and pain has relied on non-specific inhibition of atypical PKCs by ZIP, with some investigations using viral overexpression of PKC/Mζ 13 or constitutive genetic ablation. 3 This was the first known selective inhibition of the atypical isoform PKCι/λ in spinal nociceptive sensitization, as well as the first assessment of its downstream target p62/GluA1, thus allowing for further examination of spinal nociceptive role of the different atypical PKC isoforms that are inhibited by ZIP.
Non-specific atypical PKC inhibition by ZIP resulted in a male-specific reversal of allodynia induced by repeated muscular injury, consistent with previous work in this same paradigm. 3 Interestingly, the hypothesized role of PKCι/λ in the maintenance of spinal nociceptive sensitization in females was not supported by these results, as female rats continued to exhibit allodynia following administration of inhibitors to both PKCι/λ and the downstream target p62/GluA1 in multiple pain tests. These results, in addition to previous work identifying PKMζ-dependent sex differences, 3 suggest that persistent hypersensitivity in females is likely to be maintained by factors independent of atypical PKC isoforms. Interestingly, inhibition of the PKMζ downstream target NSF/GluA2 using myr-pep2m reversed allodynia following i.m. acidic saline injury in both males and females. The analgesic effect of disrupting NSF/GluA2 in females is inconsistent with previous pharmacological and genetic manipulations of PKMζ, 3 suggesting that in female rats, NSF/GluA2 interactions may be the downstream target of something other than PKMζ.
Notably, the previous research that established the PKMζ-NSF/GluA2 pathway in the hippocampus relied exclusively on male animals and thus did not assess potential sex differences.32,33 However, differential utilization of PKMζ was demonstrated across the sexes in one study of long-term spatial memory, with males showing a significant increase in synaptic PKMζ expression positively correlated with memory retention scores, while females did not. 34 Interestingly, and similar to the present findings, despite the sex divergent role of PKMζ for memory retention, both female and male rodents demonstrated a positive correlation between retention test scores and synaptic GluA2 expression. 34 Based on the findings of this present study, the common interaction between NSF and the GluA2 subunit of AMPAR appears to be driven by divergent upstream mechanisms in males and females. Further investigation into female specific mechanisms is necessary to delineate sex-specific pathways, as well as to identify the point at which a mechanistic switch occurs; reflecting a divergence from a shared molecular pathway between the two sexes underlying sex-specific signalling. 35 Future studies focused on the interactions of NSF, and potential female-specific upstream mediators could assist to identify female-specific targets involved in the maintenance of centrally-mediated persistent hypersensitivity.
CGRP signalling was hypothesized to be a potential alternative upstream mediator of NSF/GluA2 that might be female specific. However, the lack of an anti-allodynic effect of myr-pep2m following i.t. CGRP (either acutely or at 24 h) did not support this. Brain-derived neurotrophic factor has been shown to stimulate PKMζ phosphorylation and the synthesis of PKMζ and PKCι/λ through the activation of PDK1/AKT/mTOR signalling at both spinal and cortical synapses, though notably only male animals have been assessed. 14 It is of interest to determine if this regulation is similar across the sexes, given the demonstrated sex differences established by this study and similar work by Nasir et al. 3 Additionally, hyperalgesic priming tests with i.m. acidic saline, using only female mice, supports the notion that priming is induced by a glutamate-/PKC-/ERK-/AMPAR-dependent pathway leading to synaptic plasticity at the spinal dorsal horn. 36 Further investigation into possible sex differences in PKC modulation of hyperalgesic priming would be of interest.
Inhibition of both PKCι/λ and the downstream target p62/GluA1 resulted in analgesic effects in males after i.m. acetic saline. This is consistent with the effects of inhibition of PKMζ with ZIP or constitutive PRKCZ knockout on nociceptive behaviours induced by i.m. acidic saline, 3 suggesting that both atypical PKC isoforms PKCι/λ and PKMζ may contribute to the maintenance of spinal nociceptive sensitization in males. Moreover, the analgesic effect of PKCι/λ inhibition is of particular interest as it highlights a potential mechanistic difference between the maintenance of spinal nociceptive plasticity and hippocampal plasticity, as in contrast to its effects on nociception, PKCι/λ was only shown to play a compensatory role in hippocampal late-LTP following genetic ablation of PKMζ and not in wild-type animals. 24 Identifying distinctions between spinal and cortical mechanisms would be helpful for future analgesic drug development by allowing for the discovery of agents that produce analgesia but do not disrupt memory formation or cortical function.7,37 Analgesic drug development has thus far been limited by the underlying mechanistic similarities between central sensitization and hippocampal plasticity and LTP. For example, NMDA receptor antagonists such as ketamine effectively reduce pain hypersensitivity, but severely impact cortical functioning. 37 Thus, due to the differential effects of its inhibitor on hippocampal LTP and spinal nociceptive sensitization, PKCι/λ may be a potential target of interest for future drug development for males.
There were demonstrated test-specific effects across the different pharmacological manipulations used in this study. Drugs were found to have no effect in the formalin test, while they demonstrated robust reversals of mechanical allodynia following i.m. acidic saline. The formalin test involves peripheral inflammation of the ipsilateral hind paw following formalin injection. Previous work has examined the influence of the nociceptive and inflammatory effects associated with varied concentrations of formalin on the effectiveness of drugs to reduce formalin pain by targeting central sensitization.38,39 Thus, it has been proposed that the peripheral inflammation and accompanying peripheral signalling to the spinal cord may continuously contribute to pain perception, masking any effects of reduced central sensitization, as previously demonstrated by reduced ZIP effects with increasing concentrations of formalin. 3 The lack of effect of PKCι/λ, p62/GluA1 or NSF/GluA2 inhibition in the formalin pain test is consistent with the previously demonstrated lack of analgesic effect of ZIP after chronic constriction injury of the sciatic nerve,12,16 spinal nerve ligation, 17 spared nerve injury 3 or plantar incision. 18 In contrast to the formalin test, and these other pain tests, i.m. acidic saline is used as a measure of remote allodynia; pain hypersensitivity which differs in location of expression from the site of tissue injury (i.e., plantar surface of the hind paw and gastrocnemius muscle, respectively), thus allowing for a behavioural investigation of centrally-mediated mechanisms spatially separated from ongoing noxious peripheral input. The findings presented here are consistent with ZIP effects after hyperalgesic priming involving IL6/PGE2, 13 brain-derived neurotrophic factor/PGE2, 14 IL6/plantar incision, 15 (in which priming and testing are temporally separated) and repeated injections of i.pl. and intracolonic capsaicin (which reflect both temporal and spatial separation, respectively). 3 Thus, the reported differences in inhibition of atypical PKCs and downstream targets across the two pain tests (formalin, i.m. acidic saline) indicate the influence of continued afferent input on the modulation of central sensitization. These findings provide further support for the notion that analgesia produced by inhibition of PKMζ is masked by ongoing peripheral input to the spinal cord, 3 suggesting that both atypical PKC isoforms contribute mainly to the maintenance of spinal nociceptive sensitization.
Investigation into the role of PKMζ in nociceptive sensitization has not been limited to the level of the spinal cord, with ZIP administration to the anterior cingulate cortex (ACC) establishing a key role of cortical atypical PKCs in spontaneous pain evoked by nerve injury. 40 Interestingly, spinal and cortical pain mechanisms related to atypical PKCs appear to differ in regard to the influence of peripheral input, as ZIP administration to the ACC is able to alleviate neuropathic pain,17,40 despite a number of studies demonstrating no effect after i.t. injections.3,12,16 As of yet, experiments examining pain plasticity in the ACC have not examined the contribution of PKCι/λ, and there has been limited investigation into female-specific mechanisms. However, recent translational work investigating neural pain memory demonstrated that the male-specific conditioned pain hypersensitivity was abolished by both i.t. and intra-cerebral injections of ZIP, interestingly demonstrating the influence of atypical PKCs on neuronal pain memory at both the spinal and cortical levels. Additionally, these authors found that the context-dependent pain hypersensitivity in male mice was eliminated by castration or by pharmacological blockade of the hypothalamic-pituitary-adrenal axis, indicating the influences of testosterone and stress, respectively. 41 Further study into how pain memories are generated and able to maintain hypersensitivities, particularly across the sexes, is a necessary and exciting new area for future pain research.
Overall, our results demonstrate novel behavioural evidence for the male-specific role for PKCι/λ and its downstream target p62/GLuA1, in addition to PKMζ, in pain states for which central sensitization is a key component. The sexually divergent pathways appear to converge at the interaction between NSF and the GluA2 subunit of the AMPA receptor. While the NSF/GluA2 interaction is thought to be downstream of PKMζ in males, these findings and previous PKMζ-dependent sex differences 3 suggest that females may rely on a thus far unidentified factor, independent of atypical PKCs, for the maintenance of persistent pain states. These behavioural findings will inform future investigation of basic mechanisms related to the maintenance of central plasticity, a critical step towards identifying targets for analgesic drug development.
Footnotes
Acknowledgments
The authors wish to thank Dr. Robert Farese, Department of Internal Medicine, James A Haley Veterans Hospital, University of South Florida, Tampa, FL, for supplying the ICAP.
Author Contributions
TJC and NG designed the study; NG and AL conducted behavioural experiments; NG and AL performed statistical analysis; NG prepared figures; NG wrote the paper and TJC and AL supervised experiments and edited the paper.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by grants from the Natural Sciences and Engineering Research Council of Canada (RGPIN/05605), the Canadian Institutes of Health Research (MOP-119279) and the Louise and Alan Edwards Foundation. NG was supported by a Fredrick Banting and Charles Best Canada Graduate Scholarship-Master’s (CGS-M).