
Abstract
Background
The amygdala plays a key role in fear learning and extinction and has emerged as an important node of emotional-affective aspects of pain and pain modulation. Impaired fear extinction learning, which involves prefrontal cortical control of amygdala processing, has been linked to neuropsychiatric disorders. Here, we tested the hypothesis that fear extinction learning ability can predict the magnitude of neuropathic pain.
Results
We correlated fear extinction learning in naive adult male rats with sensory and affective behavioral outcome measures (mechanical thresholds, vocalizations, and anxiety- and depression-like behaviors) before and after the induction of the spinal nerve ligation model of neuropathic pain compared to sham controls. Auditory fear conditioning, extinction learning, and extinction retention tests were conducted after baseline testing. All rats showed increased freezing responses after fear conditioning. During extinction training, the majority (75%) of rats showed a decline in freezing level to 50% in 5 min (fear extinction+), whereas 25% of the rats maintained a high freezing level (>50%, fear extinction−). Fear extinction− rats showed decreased open-arm preference in the elevated plus maze, reflecting anxiety-like behavior, but there were no significant differences in sensory thresholds, vocalizations, or depression-like behavior (forced swim test) between fear extinction+ and fear extinction− types. In the neuropathic pain model (four weeks after spinal nerve ligation), fear extinction− rats showed a greater increase in vocalizations and anxiety-like behavior than fear extinction+ rats. Fear extinction− rats, but not fear extinction+ rats, also developed depression-like behavior. Extracellular single unit recordings of amygdala (central nucleus) neurons in behaviorally tested rats (anesthetized with isoflurane) found greater increases in background activity, bursting, and evoked activity in fear extinction− rats than fear extinction+ rats in the spinal nerve ligation model compared to sham controls.
Conclusion
The data may suggest that fear extinction learning ability predicts the magnitude of neuropathic pain-related affective rather than sensory behaviors, which correlates with differences in amygdala activity changes.
Introduction
Interindividual differences in anxiety-depression-like conditions 1 as well as in pain and responsiveness to pain treatment2,3 are well documented, but mechanisms and biomarkers of vulnerability and resilience related to pain remain to be determined. Pain is a complex experience characterized by its strong negative affective component. Pain may share neurobiological mechanisms with other negative emotions such as fear, including plasticity in emotional networks.4,5 Fear learning and extinction play important roles in neuropsychiatric disorders such as anxiety disorders, posttraumatic stress disorder (PTSD), obsessive–compulsive disorder (OCD), and others.6–8 For example, impaired fear extinction (FE) has been linked to anxiety disorders and PTSD in animal models and humans9,10 and is being considered as a potential biomarker for interindividual differences in predicting vulnerability to anxiety disorders and PTSD in preclinical 11 and clinical studies. 12
Exploring FE ability as a predictor for interindividual differences in pain sensitivity and magnitude of pain symptoms in chronic pain was the goal of our preclinical study in outbred rats. Another area of interest was the involvement of the amygdala in this process. The amygdala plays a key role in fear learning and extinction13,14 and has emerged as an important node of the emotional-affective aspects of pain and pain modulation.15–17 Importantly, the amygdala has also been implicated in the interaction between pain and fear in humans,18,19 and amygdala size predicts the risk for chronic pain in humans. 20 In preclinical studies, the amygdala has been linked to fear-conditioned analgesia,21–23 but its role in pain prediction based on FE ability has not been determined.
Here, we subjected adult rats to cued fear conditioning, extinction learning, and extinction retention tests and correlated interindividual differences with pain-related behaviors in a neuropathic pain model (spinal nerve ligation (SNL)) compared to sham controls. For correlation with amygdala activity, we performed extracellular single-unit recordings of neurons in the central nucleus (CeA) in behaviorally tested and phenotyped rats. The CeA receives input from the spino-parabrachio-amygdaloid pathway that plays a role in nociceptive processing 24 and fear learning. 5 Neuroplastic changes in CeA neurons result in increased activity and pain-related behaviors in models of acute and chronic pain.15,16 The results have in part been communicated in abstract form. 25
Materials and methods
Animals
Male Sprague Dawley rats (180–320 g, corresponding to 6–12 weeks of age; Harlan Laboratories) were housed in a temperature-controlled room under a 12 h light/dark cycle. Water and food were available without restriction. On the day of the experiment, rats were transferred from the animal facility and allowed to acclimate to the laboratory for at least 1 h. All experimental procedures were approved by the Institutional Animal Care and Use Committee at Texas Tech University Health Sciences Center and conform to the guidelines of the International Association for the Study of Pain and of the National Institutes of Health.
Experimental protocol
Normal naive rats were subjected to fear conditioning and FE trials. One week later, a neuropathic pain model was induced or sham surgery was performed (see “Neuropathic pain model” section). Behavioral or electrophysiological studies were done four weeks after surgery when neuropathic pain-related behavioral and neuronal changes reach a stable plateau in this model. 26 In another set of control experiments, sensory and affective behaviors (mechanosensitivity, vocalizations, and open field test (OFT) performance) were measured two days before and two days after the fear conditioning/extinction trials to rule out any changes induced by the fear conditioning and extinction tests. The experimenter was blinded with regard to the FE phenotype (see fear conditioning and extinction) and neuropathic versus sham condition.
Neuropathic pain model
The well-established SNL model 27 was used, which generates stable and long-lasting neuropathic pain behaviors. Rats were anesthetized with isoflurane (3%–4% induction and 2% maintenance). Under anesthesia and using sterile techniques, the L5 spinal nerve was tightly ligated using 6–0 sterile silk. In the sham-operated group, the nerve was exposed but not ligated.
Behaviors
Fear conditioning and extinction
Auditory fear conditioning, extinction learning, and extinction retention tests were conducted using two chambers of a near infrared Video Fear Conditioning System (Med Associates Inc., Fairfax, VT) as in our previous studies.28,29 Conditioning chambers were situated inside a sound-attenuating isolation cabinet. The metal grid floor of the conditioning chambers was connected to a grid stimulator to administer aversive shocks. Two training chambers (context A and context B) were used which were distinct with visual, olfactory, tactile, dimensional, and lighting differences. On day 1, rats were habituated to the training chamber (context A) for 5 min followed by fear conditioning, which consisted of delivering a footshock (0.7 mA, 2 s; the unconditioned stimulus, US) during the final 2 s of an auditory stimulus (white noise, 80 db, 4.5 kHz, 30 s; the conditioned stimulus, CS). Two CS-US pairings were used (intertone interval, ITI, 120 s). On day 2, rats were habituated to context B for 5 min followed by extinction training (30 CSs, ITI 5 s). On day 3, rats were habituated to context B for 5 min followed by extinction retention (5 CSs, ITI 5 s). Behavior of the rats was recorded with a video camera mounted in the conditioning chambers. Freezing behavior was analyzed and quantified off-line using Video Freeze software (Med Associates Inc.) as the conditioned response (expressed in percentage for each 30-s period). Based on their FE learning ability, rats were classified into high (FE+) and low (FE−) FE groups and then randomly divided into four groups (sham FE+, sham FE−, SNL FE+, and SNL FE−). Four weeks after neuropathic (SNL) or sham surgery, behavioral assays (see the next paragraphs) and in vivo extracellular single unit recordings of CeA neurons (see “Systems electrophysiology” section) were performed.
Mechanosensitivity
Mechanical withdrawal thresholds were measured using von Frey monofilaments (3.61, 3.84, 4.08, 4.31, 4.56, 4.74, 4.93, and 5.18, corresponding to 0.4, 0.6, 1, 2, 4, 6, 8, and 15 g) applied to the plantar surface of the hind paw at the base of the third or fourth toe, the most sensitive area in the SNL model. An abrupt withdrawal of the foot during stimulation or immediately after stimulus removal was counted as a positive response. Paw withdrawal thresholds (expressed in g) were determined by the up-down method and the formula of Dixon as in our previous studies. 26 In some experiments, a plantar electronic von Frey anesthesiometer (IITC Life Science, Woodland Hills, CA) was used to measure mechanical thresholds. The tip was applied perpendicularly to the base of the third or fourth toe with increasing force until a flexion reflex was provoked, which was automatically recorded as the paw withdrawal threshold (in grams). The average of triplicate measurements at least 30 s apart was used. 30
Emotional responses
Vocalizations in the audible (20 Hz–16 kHz) and ultrasonic (25 ± 4 kHz) ranges were measured using a condenser microphone and a bat detector, respectively, connected to a filter and amplifier (UltraVox four-channel system; Noldus Information Technology, Leesburg, VA) as in our previous studies.26,31,32 Animals were anesthetized briefly with isoflurane (2%, precision vaporizer) and placed in a custom-designed recording chamber that permitted access to the hindlimbs for mechanical test stimuli. After habituation to the chamber, brief (15 s) innocuous (100 g/6 mm2) and noxious (500 g/6 mm2) mechanical stimuli were applied to the hind paw using a calibrated forceps with a force transducer to monitor the applied force (in grams). Durations of audible and ultrasonic vocalizations were analyzed for 1 min following the onset of the mechanical stimulus using Ultravox 2.0 software (Noldus Information Technology).
Anxiety-like behavior
Animal movements in the open and closed arms of an elevated plus maze (EPM, Columbus Instruments; arm length, 50 cm; arm width, 10 cm; and wall height, 40 cm) were detected with photocells and entries into the respective arm were measured for 15 min using a computerized analysis system (Multi-Varimex software; Columbus Instruments) as described previously.26,32,33 After acclimation to the lab for 1 h, the animal was placed onto the central area of the EPM, facing an open arm. The ratio of open-arm entries to the total number of entries (expressed as percentage) during the first 5 min was calculated as is standard EPM protocol. 34 In some experiments, the OFT 35 was used to measure exploratory behavior of the animal in an arena (70 cm × 70 cm) with acrylic walls (height, 45 cm) for 15 min, using a computerized videotracking and analysis system (EthoVisionXT 11 software, Noldus Information Technology). Duration in the center area (35 cm × 35 cm) was calculated for the first 5 min. Avoidance of the open arms in the EPM and of the center field in the OFT suggests anxiety-like behavior.
Depressive-like behavior
The well-established forced swim test (FST), also termed “behavioral despair,” was used to evaluate depressive-like behavior36,37 as in our previous studies. 32 Rats were placed individually in a plastic cylinder (50 cm in height × 20 cm in diameter) partially filled with water (30 cm; 23°C–25°C). The FST paradigm includes an initial 15-min pretest, followed 24 h later by a 5-min test. Movement was recorded for 5 min by a video surveillance system for off-line analysis (EthoVisionXT 11 software, Noldus Information Technology). Total duration of time spent immobile (no active movements) over the 5-min trial was used to assess depression-like behaviors. The cylinder was cleaned carefully after each test.
Systems electrophysiology
Extracellular single-unit recordings were made from neurons in the laterocapsular division of the CeA (CeLC) as described previously.26,38,39 Recordings were made in SNL or sham rats four weeks post-surgery.
Animal preparation and anesthesia
Rats were anesthetized with isoflurane (2%, precision vaporizer, Harvard apparatus). Core body temperature was maintained at 37°C by means of a homeothermic blanket system. The animal was mounted in a stereotaxic frame (Kopf Instruments) and a craniotomy was performed at the sutura frontoparietalis level to allow the insertion of the recording electrode (glass insulated carbon filament, 1–3 MΩ). Depth of anesthesia was assessed by regularly testing the corneal blink, hind paw withdrawal, and tail-pinch reflexes and by checking for abnormal breathing patterns. Anesthesia was maintained with isoflurane (2%, precision vaporizer, Harvard apparatus).
Single-unit recording and identification of CeA neurons
Stereotaxic recordings of individual CeLC neurons were made using the following coordinates: 2.2–3.1 mm caudal to bregma, 3.8–4.2 mm lateral to midline, and depth 7–8.5 mm. The recorded signals were amplified, band-pass filtered (300 Hz to 3 kHz), displayed on an analog oscilloscope, and processed by an interface (1401 Plus; CED). Spike2 software (version 4; CED) was used for spike sorting, data storage, and analysis of single-unit activity. Spike size and configuration were monitored continuously. Only those neurons with a spike configuration that matched a “preset” template and could be clearly discriminated from activity in the background throughout the experiment were included in the study. The spike template was created for each neuron during the initial baseline recording period (5 min), capturing the waveform within set limits of variability for parameters such as amplitude, duration, and rise time, using Spike2 software. Subsequent spikes were matched to that template (spike sorting). CeLC neurons were identified by monitoring background activity and responses to mechanical compression of the hind paw (innervation territory of L5) at innocuous (100 g/6 mm2) and noxious (500 g/6 mm2) intensities with a calibrated forceps. Neurons were selected that had a receptive field in the hind paw and responded more strongly to noxious than innocuous mechanical stimuli.
Neuronal activity analysis
Spike2 software was used to measure neuronal activity as spikes/s for 10 min (background activity in the absence of intentional stimulation) and then during mechanical test stimulation (compression of the paw for 15 s). For net evoked responses, background activity (spikes/15 s) preceding the stimulus was subtracted from the total number of spikes during stimulation (15 s). Spike2 software burst analysis script was used to analyze interspike interval (ISI) distribution and burst-like activity for each neuron during a 5-min period of recorded background activity (no test stimuli). Irregular burst-like activity was defined as follows: a silent period of at least 100 ms before the first spike in a burst that was followed by a second spike with an ISI ≤10 ms as described in our previous study. 26 Any subsequent spikes with a preceding ISI ≤10 ms were also considered to be part of a burst.
Verification of recording site
At the end of each experiment, the recording site was marked by injecting DC (250 µA for 3 min) through the recording electrode. The brain was removed and submerged in 10% formalin and potassium ferrocyanide. Tissues were stored in 30% sucrose before they were frozen sectioned at 50 µm. Sections were stained with hematoxylin and eosin, mounted on gel-coated slides, and cover slipped. Lesion/recording sites were identified under bright-field microscopy histologically and plotted on standard diagrams.
Statistical analysis
All averaged values are given as the mean ± SE. Statistical significance was accepted at the level P < 0.05. GraphPad Prism 5.0 software was used for all statistical analyses. Statistical analysis was performed on the raw data. For multiple comparisons, a one-way or two-way analysis of variance (ANOVA) (repeated measures where appropriate) was used with Bonferroni post hoc tests.
Results
Interindividual differences in FE learning ability of naive rats
Fear learning and FE were measured in 109 normal naive rats (see “Methods” section). On day 1, rats showed no or minimal freezing behavior during habituation in context A, indicating normal locomotor activity, but all rats developed increased freezing responses after two parings of CS (80 db white noise, 30 s) and US (0.7 mA foot shock, 2 s) during the conditioning session (Figure 1(a)). During FE training in context B (30 CSs) on day 2, however, two groups emerged that showed differences in the decline of freezing behavior in the absence of a foot shock (US) with regard to the time course and magnitude (Figure 1(b) and (c)). Eighty-two rats (75%) showed a quick decline of freezing to levels lower than 50% (per 30 s block), reflecting extinction learning ability (FE+), whereas 27 rats (25%) maintained a freezing level higher than 50%, suggesting low extinction learning ability (FE−). Percent freezing per 30-s block was significantly higher in the FE− group compared to the FE+ group (P < 0.0001, F1,4280 = 1210.0, two-way ANOVA; Bonferroni post hoc test results are shown in Figure 1(b)). During FE retention testing in context B (5 CSs) on day 3, FE+ rats showed a lower and quicker decline in freezing compared to FE− rats that maintained a significantly higher freezing level (Figure 1(d); P < 0.0001, F1,1605 = 175.6, two-way ANOVA; Bonferroni post hoc test results are shown in Figure 1(d)). Importantly, no difference in percent freezing was observed between FE+ and FE− groups during the habituation phase (Figure 1(a), (b), and (d)) and in the fear conditioning session (Figure 1(a)).
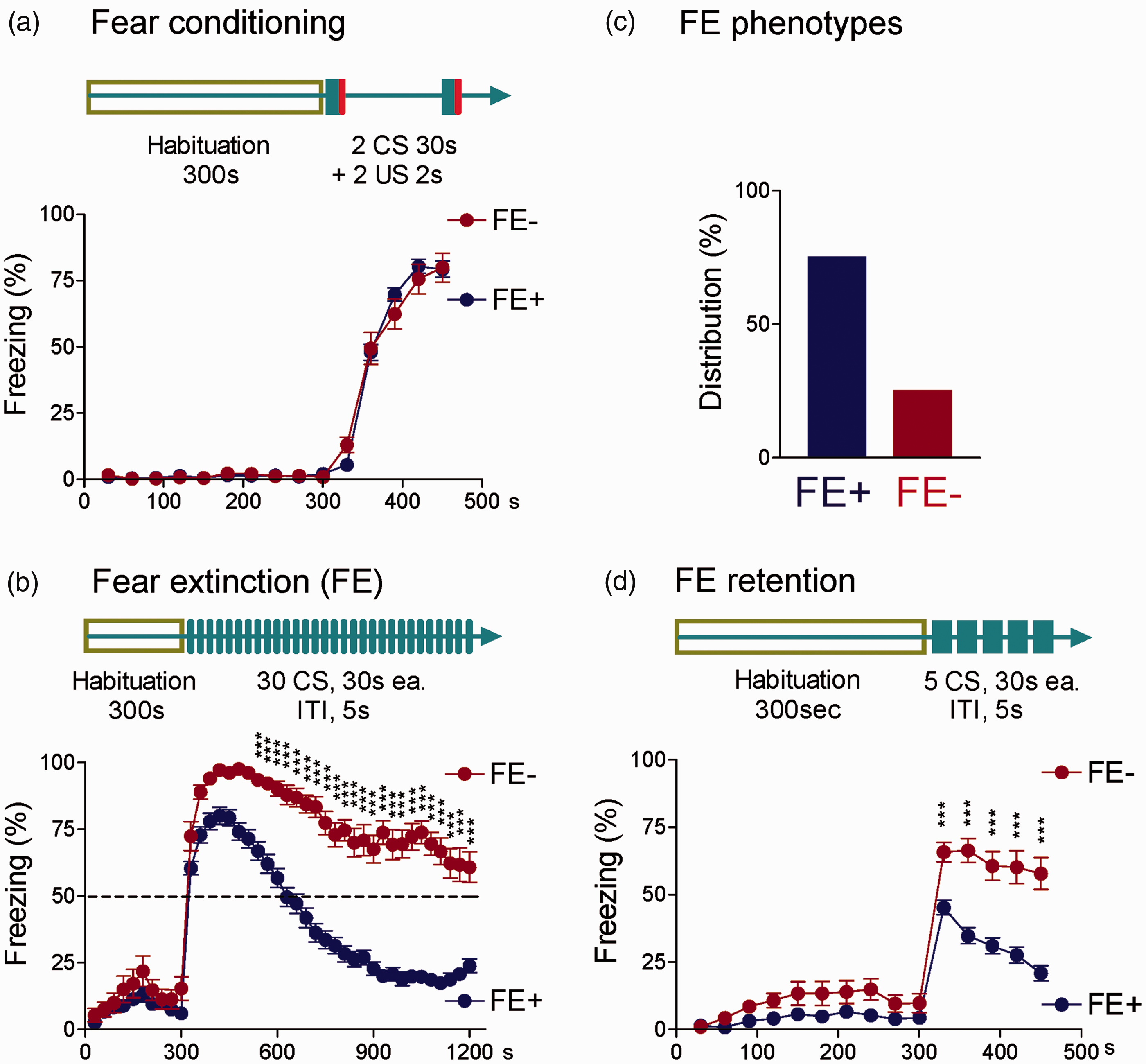
Interindividual differences in fear extinction learning ability in normal naive rats. Auditory fear conditioning (a), extinction (b), and extinction retention (d) tests were conducted using two distinct context chambers. (a) Day 1, rats were habituated to context A followed by fear conditioning (2 CS-US pairs, see Methods section). Diagram illustrates experimental protocol. Symbols in the line graph show freezing responses expressed in percent per 30-s block during fear conditioning with 2 CS-US pairings. (b) Day 2, rats were habituated to context B followed by extinction training (30 CSs, no US). Diagram illustrates experimental protocol. Symbols in the line graph show freezing responses to tone (CS) expressed in percent per 30-s block. (c) Bar histograms show distribution of rats with fear extinction (FE+) and rats with delayed and weak (freezing levels >50%) fear extinction (FE−). For details see Methods and Results section. (d) Day 3, rats were habituated to context B, followed by extinction retention testing (5 CSs, no US). Diagram illustrates experimental protocol. Symbols in the line graph show freezing response to tone (CS) expressed in percent per 30-s block. *, **, and *** indicate P < 0.05–0.001, Bonferroni post hoc tests following two-way repeated measures ANOVA (see “Results” section). ITI: intertone interval; FE: fear extinction.
Interindividual differences in pain-related behaviors of FE+ and FE− rats
Next, we examined whether interindividual differences in FE ability would translate into behavioral differences in a neuropathic pain model (SNL, see Methods) and/or in the sham control condition. To do so, FE+ and FE− rats were randomly assigned to the neuropathic group or sham group, and the following behavioral assays were done four weeks after surgery (nerve injury or sham): audible and ultrasonic vocalizations evoked by brief (15 s) noxious mechanical compression of the hind paw (emotional responses, Figure 2(a) and (b)), EPM test (anxiety-like behavior, Figure 2(c)), FST (depression-associated behavior, Figure 2(d)), and von Frey test (mechanosensitivity, Figure 2(e)).

Interindividual differences in pain-related behaviors of FE+ and FE− rats. (a and b) Duration (s) of audible and ultrasonic vocalizations, respectively, evoked by brief (15 s) noxious mechanical compression of the hind paw. Significant differences were found between FE+ (n = 11) and FE− (n = 13) neuropathic (SNL) rats but not between FE+ (n = 14) and FE− (n = 14) sham rats. *, **, and *** indicate P < 0.05–0.001, ANOVA with Bonferroni post hoc tests (see “Results” section). (c) Open-arm choice in the EPM was significantly lower in sham FE− (n = 10) than sham FE+ (n = 12) rats as well as in neuropathic FE− (n = 10) compared to neuropathic FE+ (n = 9) rats. Open-arm choice of both FE+ and FE− rats was significantly lower in the neuropathic pain model compared to sham control conditions. *P < 0.05; **P < 0.01, ANOVA with Bonferroni post hoc tests (see text). (d) Immobility (s) in the FST measured for 5 min was significantly greater in FE− (n = 9) than FE+ (n = 10) neuropathic rats, but there was no significant difference between FE− (n = 10) and FE+ (n = 8) sham rats. Neuropathic FE−, but not FE+, rats showed significantly greater immobility than their sham controls. *P < 0.05;***P < 0.001, ANOVA with Bonferroni post hoc tests (see “Results” section). (e) Mechanical thresholds (von Frey test) were not significantly different between FE− (n = 13) and FE+ (n = 42) sham rats and between FE− (n = 9) and FE+ (n= 28) neuropathic rats, but neuropathic FE− and FE+ rats had significantly lower mechanical thresholds than their sham controls. ***P < 0.001, ANOVA with Bonferroni post hoc tests (see “Results” section). Bar histograms show means ± SEM. EPM: elevated plus maze; FST: forced swim test; FE: fear extinction; SNL: spinal nerve ligation.
No significant difference in audible and ultrasonic vocalizations (Figure 2(a) and (b)) was found between sham FE+ rats (n = 14) and sham FE− rats (n = 14). In the neuropathic pain model, vocalizations of FE+ rats (n = 11) and FE− rats (n = 13) were significantly increased compared to sham controls (P < 0.05–0.001 as shown in Figure 2(a) and (b)). However, vocalizations of SNL FE− rats were significantly greater than those of SNL FE+ rats (P < 0.05). For the statistical analysis of vocalizations in the four experimental groups, ANOVA with Bonferroni post hoc tests was used (audible, F3,47 = 16.16; ultrasonic, F3,47 = 8.203).
Open-arm choice in the EPM (Figure 2(c)) was lower in FE− rats compared to FE+ rats in sham control conditions (FE−, n = 10; FE+, n = 12) as well as in the neuropathic pain model (FE−, n = 10; FE+, n = 9; P < 0.01), suggesting higher anxiety-like behavior of FE− rats. Open-arm choice of both FE+ and FE− rats decreased in the neuropathic pain model compared to sham control conditions significantly (P < 0.05). For statistical analysis of EPM performance in the four experimental groups, ANOVA with Bonferroni post hoc tests was used (F3,37 = 15.79).
In the FST (Figure 2(d)), no significant difference in immobility was found between sham FE+ rats (n = 8) and sham FE− rats (n = 10). In the neuropathic pain model, however, FE− rats (n = 9), but not FE+ rats (n = 10), showed significantly increased immobility compared to sham controls (P < 0.001), and so a significant difference emerged between SNL FE− rats and SNL FE+ rats (P < 0.05), suggesting that it is FE− rats, but not FE+ rats, that develop depression-associated behaviors. For statistical analysis of FST performance in the four experimental groups, ANOVA with Bonferroni post hoc tests was used (F3,33 = 15.48).
In the von Frey test of mechanosensitivity (Figure 2(e)), there was no significant difference between sham FE− rats (n = 13) and sham FE+ rats (n = 42) and between SNL FE− rats (n = 9) and SNL FE+ rats (n = 28), but mechanical thresholds of both FE− and FE+ rats in the neuropathic pain model were lower than those of their sham controls (P < 0.001), suggesting that both types of rats developed neuropathic hypersensitivity. For statistical analysis of mechanosensitivity in the four experimental groups, ANOVA with Bonferroni post hoc tests was used (F3,88 = 88.11).
Fear conditioning and extinction testing did not change pain-related behaviors
In a separate set of experiments, mechanosensitivity, emotional responses, and anxiety-like behaviors were measured two days before and two days after the fear conditioning/extinction trials to determine if the fear conditioning/extinction tests themselves had any effect on these behaviors (Figure 3). There was no significant difference in vocalizations of FE+ (n = 21) and FE− rats (n = 9) before and after fear conditioning and extinction trials (audible, F3.56 = 0.3379 and “ultrasonic, F3.56 = 0.1215”, Figure 3(a); ANOVA with Bonferroni post hoc tests). For repeated measurements of anxiety-like behaviors, we used the OFT rather than EPM, because retesting in the EPM is considered problematic. 34 No significant difference in time spent in the center of the OFT was found in FE+ (n = 21) and FE− rats (n = 9) before and after fear conditioning/extinction trials (F3.56 = 0.008120, Figure 3(b)). There was no change in mechanosensitivity of FE+ (n = 21) and FE− rats (n = 9) measured with the electronic von Frey anesthesiometer (F3.56 = 0.1910). There were also no significant differences between FE+ and FE− rats in these tests.

Comparison of behaviors before and after fear conditioning/extinction trials. Measurements were made in normal naive FE+ (n = 21) and FE− (n = 9) rats two days before (pre-FE) and two days after (post-FE) the fear conditioning/extinction trials. (a) Duration (s) of ultrasonic vocalizations evoked by brief (15 s) noxious mechanical compression of the hind paw. (b) Center duration in the OFT. (c) Mechanical thresholds measured with an electronic von Frey anesthesiometer. There were no significant differences between FE+ and FE− rats and between pre-FE and post-FE values in any of these tests (ANOVA; see Results section). Bar histograms show means ± SEM. OFT: open field test; FE: fear extinction. *, **, *** P < 0.05, 0.01, 0.001.
Interindividual differences in amygdala neuronal activity of FE+ and FE− rats
The amygdala plays a key role in fear and anxiety14,40,41 and in emotional-affective aspects of pain and pain modulation.15,16 Although distinct and overlapping elements of the amygdala circuitry involved in pain and fear remain to be determined in detail, the central nucleus (CeA) serves important output functions for both conditions. Therefore, we sought to determine if the FE+ and FE− phenotypes showed differences in CeA neuronal activity under control conditions and/or in the SNL model of neuropathic pain. Extracellular single unit recordings of CeA neurons in the laterocapsular division (CeLC) were made in four groups of rats four weeks after SNL or sham surgery: sham FE+, sham FE−, SNL FE+, and SNL FE− rats (Figure 4, individual neurons; Figure 5, summary). The CeLC is the target of nociceptive inputs from the parabrachial area and is referred to as the “nociceptive amygdala.”15,42 Recording sites in the CeLC are shown in Figure 6. CeLC neurons were selected that responded more strongly to noxious than innocuous stimuli as in our previous studies.26,43,44

Examples of individual amygdala (CeLC) neurons in FE+ and FE− sham and SNL rats. (a and b) Top, peristimulus time histograms show the number of action potentials (spikes) per second for individual neurons. Innocuous (100 g/6 mm2) and noxious (500 g/6 mm2) stimuli (compression of the hind paw) are indicated by horizontal lines (15-s duration). Bottom, joint ISI plots (previous ISI against the subsequent ISI) show burst activity of individual neurons indicated by the rectangular insets (dots within the dashed red lines represent the first spike in a burst). Each dot represents a burst event in the individual neuron shown in each of the four plots. (a) Individual CeLC neurons recorded in sham FE+ rats and in sham FE− rats. (b) Individual CeLC neurons recorded in SNL FE+ rats and in SNL FE− rats. Individual CeLC neurons (8) from different experimental groups (4); one neuron recorded per animal. ISI, interspike interval; FE: fear extinction.

Sample of CeLC neurons in FE+ and FE− sham and SNL rats. Background activity (in the absence of any intentional stimulation) (a) and responses of CeLC neurons to brief (15 s) innocuous (b), and noxious (c) compression of the hind paw with a calibrated forceps. For net evoked responses, background activity (spikes/15 s) preceding the stimulus was subtracted from the total number of spikes during stimulation (15 s). (d) Mean number of bursts (see dots in Figure 4 meeting the burst definition) in a 5-min period of recording background activity of neurons in each experimental group. Bursts/s were calculated for each neuron by dividing the number of bursts by 300 s (n[burst]/300 s). Bar histograms show means ± SEM. Recordings were made in sham FE+ rats (n = 9 neurons), sham FE− rats (n = 7 neurons), SNL FE+ rats (n = 8 neurons), and SNL FE− rats (n = 9 neurons). *, **, and *** indicate P < 0.05–0.001, ANOVA with Bonferroni post hoc tests (see “Results” section). FE: fear extinction; SNL: spinal nerve ligation. *, **, *** P < 0.05, 0.01, 0.001.
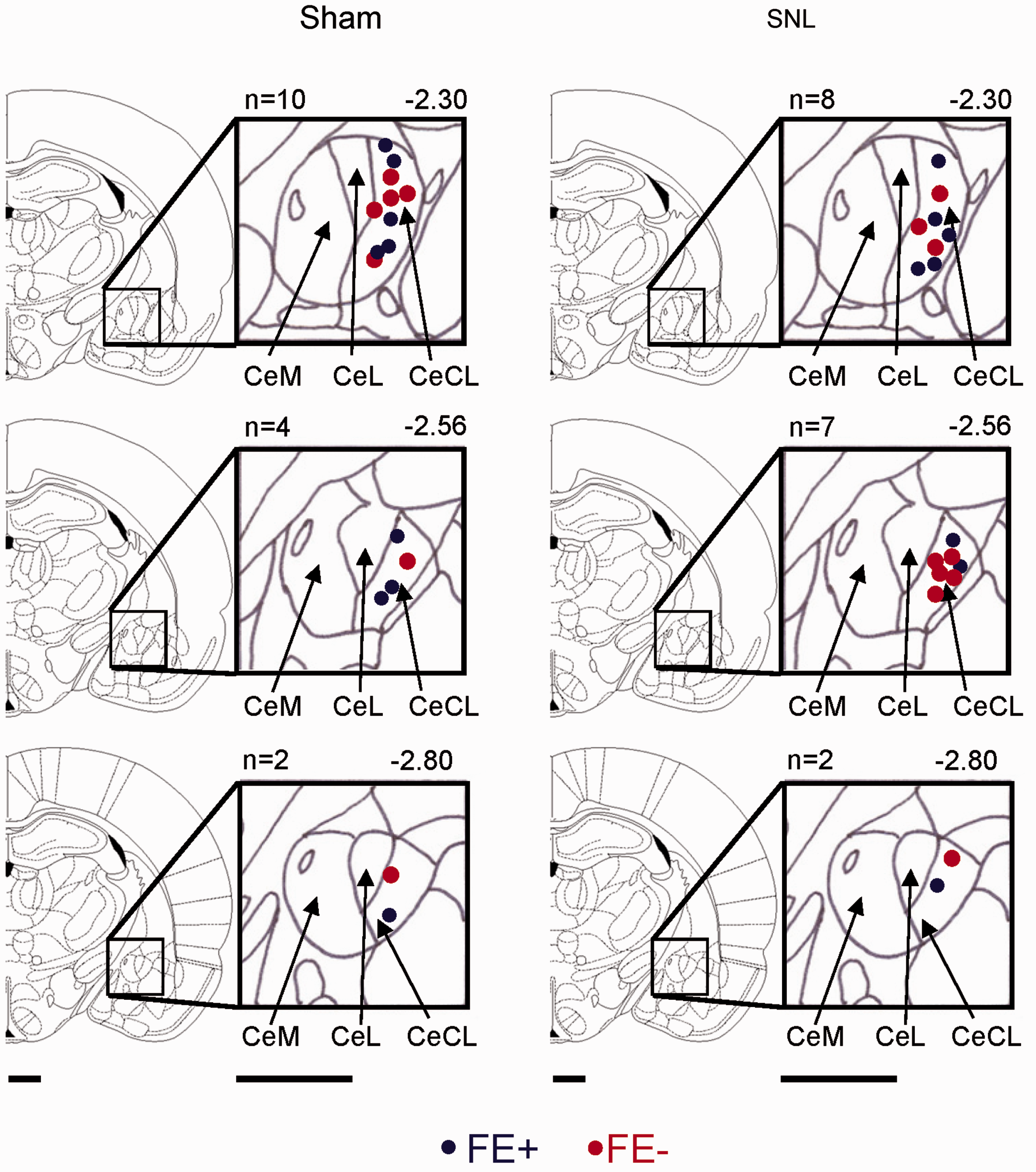
Histologically verified recording sites of CeLC neurons in FE+ and FE− sham and SNL rats. Diagrams show coronal brain sections at different levels posterior to bregma (−2.30 to −2.80). The medial (CeM), lateral (CeL), and latero-capsular (CeLC) subdivisions of the central nucleus of the amygdala (CeA) are shown next to each section. Blue symbols, neurons recorded in FE+ rats and red symbols, neurons from FE− rats. Scale bars are 1 mm. FE: fear extinction; SNL: spinal nerve ligation.
In the sample of CeLC neurons recorded in sham rats, no significant difference was found between neurons in FE+ (n = 9 neurons) and FE− (n = 7 neurons) rats with regard to background activity measured as spikes/s in the absence of any intentional stimulation and in their responses to brief (15 s) innocuous and noxious compression of the hind paw with a calibrated forceps (see “Materials and Methods” section) (P > 0.05, ANOVA with Bonferroni post hoc tests). For net evoked responses, background activity (spikes/15 s) preceding the stimulus was subtracted from the total number of spikes during stimulation (15 s). Background and evoked activity of CeLC neurons recorded in neuropathic FE− rats (n = 9 neurons) was significantly higher than in neuropathic FE+ rats (n = 8 neurons). The results of the statistical comparison of data from the four experimental groups using ANOVA with Bonferroni post hoc tests were as follows: background activity, P < 0.001, F3,29 = 17.56; responses to innocuous stimulation, P < 0.001, F3,29 = 20.27; responses to noxious stimulation, P < 0.05, F3,29 = 9.089 (Figure 5(a) to (c), respectively).
For a more refined analysis of background activity, we analyzed firing patterns as described in Materials and Methods section and in our previous work. 26 Irregular burst-like activity was defined as a silent period of ≥100 ms preceding the first spike in a burst followed by a second spike with an ISI ≤10 ms. Any subsequent spikes with a preceding ISI ≤10 ms were also considered as part of a burst. Joint interspike interval scatter plots (previous ISI against the subsequent ISI) showed more burst events (indicated for individual neurons by the rectangular insets in Figure 4) of CeLC neurons recorded in sham FE− rats (n = 7 neurons) compared to sham FE+ rats (n = 9) and in neuropathic FE− rats (n = 9 neurons) compared to neuropathic FE+ rats (n = 8 neurons). For each neuron, the number of bursts in a 5-min period of recorded background activity was calculated by dividing the number of bursts by 300 s (n[burst]/300 s). Mean number of bursts was then calculated for the sample of neurons in each experimental group. Burst-like activity of CeLC neurons was significantly higher in neuropathic rats than in sham controls. For statistical comparison of burst-like activity from the four experimental groups, ANOVA with Bonferroni post hoc tests was used (F3,29 = 18.73, see significance of individual comparisons in Figure 5(d)).
The data suggest that phenotypic differences between FE− and FE+ rats in neuropathic pain-related behaviors are reflected in differences in amygdala (CeA) activity.
Discussion
This study explored differences in FE ability of outbred rats as a predictor for interindividual differences in pain sensitivity and magnitude of pain symptoms in a neuropathic pain model. The key novelties here are the identification of distinct behavioral phenotypes based on their FE ability that showed different levels of neuropathic pain-related behaviors and amygdala neuronal activity. The rationale for selecting fear learning and extinction assays to identify interindividual differences in the development of chronic neuropathic pain in the preclinical setting includes the following.
Fear conditioning and extinction are well-established models of aversive learning and of adjustment or inhibitory control of these aversive associations.8,41 Interindividual differences such as fast and slow extinction phenotypes were found in outbred rats 45 that correlated with differences in extinction retention,45,46 anxiety-like behavior in the EPM, 46 and early life stress-induced anxiety-like behavior. 11 Importantly, these extinction-based phenotypes showed no differences in fear learning. Results of our study are consistent with these findings. Interestingly, interindividual differences in the modulation of fear responses have also been linked to vulnerability for anxiety disorders and post-traumatic stress disorder (PTSD) in humans,8,12,47,48 and there is evidence for delayed and/or reduced FE or extinction recall in patients with anxiety disorders, PTSD, and OCD.7,9,10,49,50
Interindividual differences are also well documented for pain and pain modulation. 3 Interactions of multiple biopsychosocial factors, including pain-related fear and avoidance, have been linked to pain persistence, which can be modified with extinction-related interventions.51,52 Pain and fear may share neurobiological mechanisms such as plasticity in emotional networks, 4 and the corticolimbic system in particular has emerged as a key player in pain modulation and prediction of chronic pain53,54 and in the pain–fear relationship. 19 Amygdala size and amygdala connectivity with the medial prefrontal cortex (mPFC) and nucleus accumbens network have been shown to predict chronic pain in humans. 20 Interactions between mPFC and amygdala also serve as the neurobiological substrate for FE.8,41,55
The amygdala plays a key role in fear and anxiety14,40,41 and in emotional-affective aspects of pain and pain modulation,15–17 and the central nucleus (CeA) serves output functions for both systems. The lateral-capsular divisions of the CeA are referred to as the “nociceptive amygdala”15,42 and receive nociceptive input from the spino-parabrachio-amygdaloid pathway 24 that plays a role in pain-related behaviors,15,16,56 anxiety-like behaviors, 57 as well as in fear learning. 5 Synaptic plasticity of CeLC neurons, including at the parabrachial-CeLC synapse,58,59 has been shown in different pain models, including formalin test, 60 knee joint arthritis, 61 colitis, 62 and nerve injury. 63 Pain-related neuroplasticity in this circuity results in increased neuronal activity in the amygdala output region in acute 64 and chronic26,65 pain models and drives pain-related behaviors and behavioral changes, including sensitivity, emotional responses, anxiety, and depression.15–17
Pain-related neuroplasticity in the CeA can develop because of an excitation-inhibition imbalance that may in part be due to impaired mPFC-driven feedforward inhibition of CeLC neurons.16,66,67 Brain slice physiology studies showed decreased glutamate-driven synaptic inhibition (feedforward inhibition) of CeLC neurons in an arthritis pain model.68,69 In that study, electrical stimulation of the external capsule was used, which may include mPFC inputs into the amygdala. 68 A more definitive optogenetic approach to activate mPFC axon terminals in the amygdala showed that infralimbic, but not prelimbic, mPFC-driven feedforward inhibition of CeLC neurons and synaptic activation of basolateral amygdala (BLA) neurons was decreased in brain slices from arthritic rats. 70 Conversely, systems electrophysiology studies found that pharmacologic activation of mPFC neurons inhibited the increased CeA neuronal activity in an arthritis pain model. 44 Decreased mPFC pyramidal cell activity in this pain model was caused by abnormally enhanced BLA-driven feedforward inhibition, resulting in cognitive deficits in a decision-making task.44,71,72
Decreased infralimbic mPFC activity has also been implicated in extinction deficits.73–76 During extinction, activation of the mPFC, particularly its infralimbic region, was found to inhibit CeA activity.55,77,78 Therefore, mPFC control of amygdala activity could play an important role in FE as well as in pain modulation, and differences in this cortico-limbic interaction could serve as a link between extinction deficits and pain vulnerability and severity. The close functional relationship between fear and pain modulation, and similar neurobiological mechanisms, despite potential differences at the cellular and synaptic levels, 21 led us test the hypothesis that FE learning ability can predict pain vulnerability, defined as the magnitude of behavioral changes in a pain condition.
In the present study, distinct behavioral phenotypes differed in their FE but not fear learning ability, which is consistent with data in the literature as mentioned earlier, and there were no baseline differences in mechanosensitivity, emotional-affective responses (vocalizations), or depression-like behavior (FST). Weak extinction ability correlated with anxiety-like behavior in the EPM (Figure 2(c)), but not OFT (Figure 3(b)), under sham control conditions. Greater anxiety-like behavior of the slow extinction phenotype in the EPM was reported in the literature 46 but not found in another study. 11 Differences between FE+ and FE− phenotypes in the EPM, but not OFT, may reflect the OFT modeling rather exploratory behavior influenced by “normal anxiety.” 35 FE+ and FE− phenotypes showed differences in the magnitude of neuropathic pain-related changes (increased vocalizations and development of depression-like behavior). These differential changes were not the result of the fear conditioning/extinction trials and/or the recovery of fear after extinction (“relapse”),79,80 because no differences were observed in mechanosensitivity (electronic von Frey) and averse affective behaviors (vocalizations, OFT) measured before and after these trials.
The electrophysiological analysis of CeLC amygdala neurons detected significant differences in background and evoked activity between FE+ rats and FE− rats in the neuropathic pain model but not in sham controls, when the firing rate (number of spikes/s) was used. A more fine grained analysis of firing patterns, however, detected differences in irregular burst-like firing of amygdala neurons between FE− rats and FE+ rats both in sham controls and in the neuropathic pain model, suggesting perhaps that phenotypic baseline differences may be more subtle but could be captured with a detailed analysis of synaptic and cellular processes. It should also be noted that the observation of neuronal activity changes in different behavioral phenotypes does not establish casualty, but the current study provides the rationale for more detailed and mechanistic analyses. Interindividual differences at the synaptic and cellular levels that underlie resilience in neuropsychiatric disorders in general remain to be determined, 81 and this is also true for pain and for mechanisms underlying neuronal activity differences observed here. They may include epigenetic factors affecting neurobiological systems as has been suggested for vulnerability phenotypes in stress-induced psychopathology. 82
Conclusion
The data may suggest a positive correlation between extinction learning ability and neuropathic pain control through a mechanism that involves the amygdala. Synaptic and cellular mechanisms remain to be determined.
Footnotes
Authors’ Contributions
JG and VN conceived the study, designed the experiments, and wrote the manuscript. JG, VY, TK, and PP carried out the experiments. All authors participated in the data analysis and interpretation of results and read and approved the final manuscript.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work is supported by the National Institutes of Health grants NS038261, NS081121, and NS106902.