
Abstract
Spinal nociceptive transmission receives biphasic modulation from supraspinal structures. Recent studies demonstrate that the anterior cingulate cortex facilitates spinal excitatory synaptic transmission and nociceptive reflex. However, whether the top-down descending facilitation can cause long-term synaptic changes in spinal cord remains unclear. In the present study, we recorded C-fiber-evoked field potentials in spinal dorsal horn and found that the anterior cingulate cortex stimulation caused enhancement of C-fiber-mediated responses. The enhancement lasted for more than a few hours. Spinal application of N-methyl-D-aspartate (NMDA) receptor antagonist D-AP5 abolished this enhancement, suggesting that the activation of the NMDA receptor is required. Furthermore, spinal application of methysergide, a serotonin receptor antagonist, also blocked the anterior cingulate cortex-induced spinal long-term potentiation. Our results suggest that the anterior cingulate cortex stimulation can produce heterosynaptic form of long-term potentiation at the spinal cord dorsal horn, and this novel form of long-term potentiation may contribute to top-down long-term facilitation in chronic pain conditions.
Keywords
Introduction
Spinal nociceptive transmission receives modulation from supraspinal structures such as the anterior cingulate cortex (ACC), midbrain periaqueductal grey (PAG), and rostral ventral medulla (RVM).1–4 Electrical or chemical stimulation on the supraspinal structures, especially the RVM, produces facilitatory or inhibitory effect on the spinal transmission.2–7 However, whether the descending modulation is able to cause long-term synaptic changes in spinal cord remains unclear.
In addition to the RVM, selectively activating the ACC also modulate peripheral nociceptive threshold.1,8 The ACC is well documented to play a critical role in chronic pain and related emotional disorders.9–13 In anesthetized rats, at most sites within the ACC, electrical stimulation of the ACC produced significant facilitation of spinal nociceptive tail flick reflex. Focal application of the agonist of metabotropic glutamate receptors (mGluRs) in the ACC also produced a facilitatory effect. 1 Consistently, in awaked mice, optogenetic activation of pyramidal neurons in the ACC reduced nociceptive thresholds, while activating inhibitory GABAergic interneurons had an inhibitory effect on nociception. 8 In recent studies, Chen et al.14–16 reported that the ACC-spinal cord projecting fibers made excitatory synapses with neurons in spinal dorsal horn. Stimulation of the ACC directly potentiated the spinal excitatory synaptic transmission and spinal neuronal Ca2+ responses. Selective activation of the ACC-spinal cord projecting neurons induced behavioral sensitization in pain responses.
In the present study using in vivo C-fiber-evoked field potential recording technique, we recorded a novel form of spinal long-term potentiation (LTP) induced by the ACC stimulation. The LTP could last for more than 2 h. Applying 2% lidocaine into the RVM did not affect this form of LTP. In addition, this LTP is both NMDA receptor dependent and serotonin dependent. Our findings further confirmed the direct top-down facilitation of spinal sensory transmission from the ACC in a RVM-independent way.
Materials and methods
Animal
Adult, male Sprague-Dawley albino rats weighing 250–350 g were used in experiments. Animals were randomly housed under a 12-h light-dark cycle (9 a.m. to 9 p.m. light), with food and water freely available, at least 1 week before carrying out experiments. The Animal Care and Use Committee of Xi’an Jiaotong University approved the research protocol.
Surgical preparation for in vivo field recording
Rats were anesthetized with an intraperitoneal injection of urethane (1.5 g/kg; Sigma Aldrich). The recording procedure of C-fiber-evoked field potential has been described previously. 17 Briefly, the rats were anesthetized with 20% urethane and placed in a stereotaxic frame. The body temperature was maintained at 37°C–38°C by a feedback-controlled heating blanket. A laminectomy was performed to expose the lumbar enlargement (L4 and L5 segments) of the spinal cord. The dura matter was incised longitudinally. The left sciatic nerve was exposed and stimulated by a bipolar silver chloride hook-electrode. All exposed nerve tissues were covered with warm paraffin oil in a pool made of skin flaps. An incision was made over the skull and the surface was exposed. A small hole was drilled above the contralateral side of the ACC and a concentric bipolar electrode was placed. Therefore, the final coordinates for stimulation would be 1.0 mm anterior to the bregma, 0.8 mm lateral to the midline, and 2.30 mm ventral to the surface of the skull for the ACC.
Electrophysiological recording and nerve stimulation
Field potentials were recorded at a depth of 100–500 μm from the dorsal surface of the spinal cord in ipsilateral lumbar enlargement with a glass microelectrode (filled with 0.5 M sodium acetate, impedance 0.5–1 MΩ). The glass pipette was driven by an electronically controlled microstepping motor. An A/D converter was used for digitization. The sampling rate was 10 kHz. To evoke field potentials in spinal dorsal horn, the test stimuli (0.5 ms duration per min) were delivered to the sciatic nerve, with the strength adjusted to 1.5–2 times of threshold for C-fiber response (5–7 V). The amplitudes of C-fiber-evoked field potentials were measured as the maximal distance from the baseline. The high-frequency stimulation (HFS, 100 Hz at the intensity of 100 μA, the latency of 100 μs, 100 pulses repeated every 5 s for 150 s) was applied on the ACC. For drug application, a small well was formed on the cord dorsum at the recording segments with 1.5% agar dissolved with 0.9% saline. Only one experiment was conducted in each animal.
Statistical analysis
Data are expressed as means ± S.E.M. Potentials recorded at 1 min intervals were averaged. The mean amplitudes of the averaged responses before saline or drug application served as baseline. The amplitude of C-fiber-evoked field potential was expressed as percentage of baseline.
Results
The ACC-spinal LTP in spinal dorsal horn neurons
Previous studies demonstrated that LTP of C-fiber-evoked field potentials can be induced in spinal dorsal horn by tetanic stimulation to the ipsilateral sciatic nerve.18,19 To investigate whether LTP of C-fiber-evoked field potentials may be induced by stimulating the ACC, we employed a stimulation protocol (100 Hz at the intensity of 100 μA, the latency of 100 μs, 100 pulses repeated every 5 s for 150 s, Figure 1(c)) in the ACC of anesthetized rats. Meanwhile, a small test stimulus was applied on the contralateral sciatic nerve per minute (Figure 1). C-fiber-evoked field potentials were recorded in superficial layers of spinal dorsal horn (Figure 1(b)).
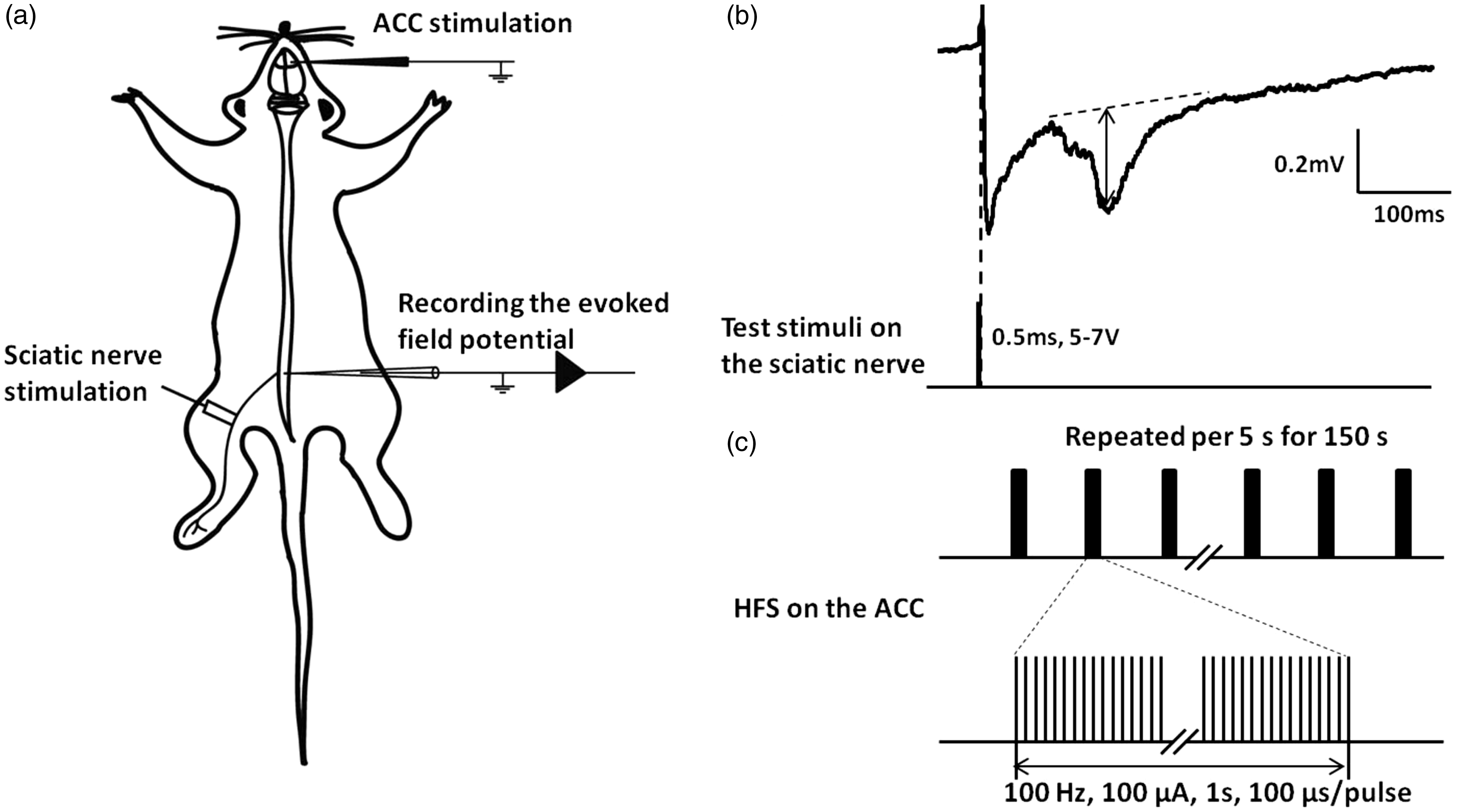
The ACC stimulation facilitates C-fiber-evoked field potential in the spinal dorsal horn. (a) Schematic diagram of the experimental arrangement. (b) The field potential (top) after stimulating the C-fiber in sciatic nerve by a single test stimulus (bottom). The amplitudes of C-fiber-evoked field potentials, measured by the maximal distance from the baseline (arrow shown in b). (c) High-frequency stimulation was applied in the deep layer of the ACC. ACC: anterior cingulate cortex.
LTP of C-fiber-evoked field potentials was induced by HFS (Figure 1(c)) in the ACC in all five tested rats (n = 5; see Figure 2). The amplitudes of C-fiber responses were significantly increased and reached to a stable level at 30 min after the ACC stimulation (136.4% ± 8.4% of the baseline, n = 5; p < 0.05, the amplitude was obtained at the last 10 min of the 2-h recording.). This potentiation lasted for more than 3 h. These results demonstrate that stimulation of the ACC is sufficient to induce stable LTP in C-fiber-evoked field potentials in spinal dorsal horn in vivo.

Activation of the ACC-induced LTP of C-fiber-mediated responses in spinal dorsal horn. (a) Representative sample and original traces of the C-fiber-evoked field potentials. HFS was applied in the ACC 30 min after baseline recording. Original traces of the field potentials, which were recorded before the ACC stimulation (a), 30 min after HFS (b), and 2 h after HFS (c), were shown. (b) The averaged amplitude of C-fiber-evoked field potentials in five rats showing that HFS of the contralateral side of the ACC induced a spinal heterotopic LTP of C-fiber-evoked field potentials that lasted for 3 h. (c) Microinjection of 2% lidocaine (0.5 μl) into bilateral RVM-enhanced C-fiber response for 25 min. Then the test stimulation intensity was decreased to reset the baseline for about 30 min, and HFS was delivered to the contralateral side of the ACC to induce spinal heterotopic LTP. ACC: anterior cingulate cortex.
The ACC-spinal LTP is independent of RVM
It is well known that the RVM is important for descending biphasic regulation of spinal nociceptive transmission.5–7,20 To test if the RVM activity is required for the ACC-induced spinal heterosynaptic LTP, we examined the effect of RVM blockade on the ACC-induced potentiation. After obtaining stable baseline C-fiber responses, we applied 2% lidocaine (0.5 μl) microinjection into bilateral RVM. As shown in Figure 2(c) (n = 5), blocking RVM significantly increased spinal C-fiber-evoked field potential at 5 min after lidocaine administration. This increase persisted for a long period of time and is likely due to the loss of tonic descending inhibition from the RVM. To prevent the saturation of C-fiber responses, we lowered the intensity of testing stimulation to reset the field potential baseline to pre-RVM injection level. After obtaining stable responses, we delivered HFS to the ACC. We found that the ACC stimulation induced a significant potentiation that persisted for at least 1 h (mean 138.7% ± 20.0% of pre-baseline. n = 5, p < 0.05). The results indicate that inhibition of RVM does not affect the ACC-spinal LTP.
Activation of NMDA receptors is required for the ACC-spinal LTP
The activation of NMDA receptors is proved to be necessary for C-fiber-evoked LTP.17,19,21 Spinal application of NMDA receptor antagonists before tetanic stimulation on sciatic nerve blocked the induction of spinal LTP. To investigate whether the top-down LTP is NMDA receptor dependent, we applied D-AP5 (200 μM), an NMDA receptor antagonist on the spinal cord 30 min before the HFS. The induction of LTP is abolished within 1 h (mean: 102.76% ± 8.58% of baseline. Figure 3, n = 6), indicating that the ACC-spinal LTP is NMDA receptor dependent.

NMDA receptors were necessary for the induction of top-down LTP. (a) Amplitudes of C-fiber-evoked field potential. The NMDA receptor antagonist D-AP5 (200 μM) was superfused to the spinal cord 30 min after baseline recording. HFS was applied in the ACC 30 min after drug application. Original traces of the field potentials, which were recorded before the D-AP5 application (a), before HFS (b), and after HFS (c), were shown. (b) The final averaged amplitude of C-fiber-evoked field potentials in six rats shown that D-AP5 blocked the induction of LTP within 1 h. ACC: anterior cingulate cortex.
Serotonin (5-HT) receptors are required for the ACC-spinal LTP
Since 5-HT is the key neurotransmitter of the descending projection from the RVM to the spinal cord and plays an important role in descending nociceptive transmission, 22 we also investigated the role of 5-HT for the ACC-spinal LTP. Since the ACC-spinal LTP is independent of RVM activity, we expect that the ACC-spinal LTP may not be affected by spinal application of methysergide. However, methysergide (antagonist of 5-HT, 30 μM) application 30 min before the ACC HFS also blocked the induction of LTP (mean 86.26% ± 2.82% of baseline. Figure 4, n = 3) within at least 2 h.

The induction of top-down LTP by the ACC stimulation requires the activation of spinal 5-HT receptors. (a) Sample and original traces of amplitudes of C-fiber-evoked field potential. The 5-HT receptor antagonist methysergide (30 μM) was superfused to the spinal cord 30 min after baseline recording. HFS was applied in the ACC 30 min after drug application. (b) The final averaged amplitude of C-fiber-evoked field potentials in three rats shown that methysergide blocked the induction of LTP. ACC: anterior cingulate cortex.
Discussion
In this study, we demonstrate that the ACC HFS induced a novel, heterosynaptic form of LTP in spinal cord dorsal horn of adult rats in vivo. Especially, this is a form of LTP that potentiates nociceptive C-fiber-evoked potentials in the spinal dorsal horn. The induction of this heterosynaptic LTP was independent to RVM, consistent with a recent report of the ACC top-down facilitation. Both NMDA receptors and 5-HT receptors are required. Future studies are clearly needed to investigate signaling mechanisms for this form of LTP in vivo.
LTP of C-fiber-mediated responses: Homo- and heterosynaptic
Two major forms of LTP have been reported in central nervous system: homo- and heterosynaptic LTP. Homosynaptic LTP is commonly investigated in brain regions such as hippocampus and the ACC. Heterosynaptic LTP, especially, under in vivo condition, is less investigated. 23 In spinal cord dorsal horn, homosynaptic LTP of C-fiber-evoked potential in superficial spinal dorsal horn was induced by stimulating the primary afferent C-fiber in the sciatic nerve. 17 Spinal application of NMDA receptor antagonist before HFS of the sciatic nerve blocked the induction of the spinal LTP. 17 A recent study suggests that the ACC projecting fibers form excitatory synapses onto spinal dorsal horn neurons.14–16 In the present study, we also found that the activation of NMDA receptor is required for the ACC-spinal LTP of C-fiber-evoked field potentials. Future studies using in vitro and in vivo approaches are needed to reveal synaptic mechanisms for the ACC-spinal LTP.
Independent of tonic descending inhibition from RVM
It is well known that the brainstem RVM is a key relay for descending inhibitory modulation from the midbrain.24,25 In the present study, we found that blocking the RVM by 2% lidocaine caused an increase in the spinal-evoked field potentials. This result is consistent with the previous findings of tonic descending inhibitory effect of the RVM on the C-fiber-evoked field potentials.26,27 Descending facilitatory systems are generally covered by tonic descending inhibition. In this study, we found that the ACC-spinal LTP can be still induced after the blockade of the RVM. This finding further confirms the existence of direct top-down facilitation pathway from the ACC to spinal cord.14,15 Early studies demonstrated that the PAG mostly exerts descending inhibition on spinal nociceptive transmission. 2 It is thus unlikely that ACC may affect spinal plasticity through the PAG projection.
Synaptic mechanism for heterosynaptic LTP
In Aplysia, a heterosynaptic modulatory pathway in the gill-withdraw reflex of Aplysia has been well documented. 28 There is direct monosynaptic connection from sensory neurons in the siphon to motor neurons in the gill. Tail stimulation activates modulatory interneuron that act on the terminals of the sensory neurons and triggers heterosynaptic long-lasting sensitization. In spinal dorsal horn, there were also some reports about heterosynaptic LTP. In spinalized animals, prolonged burst stimulation of primary afferent Aδ-fibers induces potentiation of C-fiber-evoked field potentials, 26 and this form of potentiation also requires NMDA receptor. In lamina I neurons, conditioning stimulation of primary afferent fibers with also triggered LTP at GABAergic synapses (LTPGABA). 29 This form of heterosynaptic LTP requires the activation of group I mGluRs.
In our study, spinal application of D-AP5 before the ACC stimulation blocked the induction of the spinal LTP. The results indicate that the activation of NMDA receptors is necessary. It is unclear that why methysergide also blocked the ACC-spinal LTP, because the ACC-spinal LTP is not affected by RVM blockade. We cannot completely rule out the possibility that RVM blockade was incomplete in the present study. It is possible that the ACC-spinal and the ACC-RVM-spinal pathways may work together in whole animal conditions. More studies are clearly needed to investigate these interactions in future.
In summary, we report a novel form of the ACC-spinal LTP in the spinal cord dorsal horn. This form of LTP will allow descending facilitation of spinal nociceptive transmission to persist for a long period of time. Considering that the ACC synapses are potentiated for a long period of time in chronic pain conditions,10,11,13 it is likely that the ACC excitation may lead to long-term enhancement of spinal sensory transmission including C-fiber-mediated nociceptive transmission. Understanding molecular mechanisms for such positive feedback at synaptic and circuit level will help to design better treatment for chronic pain in future.
Footnotes
Author Contributions
QYC, TC, and MZ designed the experiments. QYC performed experiments and analyzed data; QYC, TC, and MZ drafted the manuscript and finished the final version of the manuscript. All authors read and approved the final manuscript.
Acknowledgment
The authors would like to thank Yongyong Li for maintaining the recording system. The authors also thank Weijie Zeng and Jun Zhang for their constructions of the experiment.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was partially supported by grants from the CIHR project grant (PJT-148648) awarded to MZ.