
Abstract
Neuroligin1 is an important synaptic cell adhesion molecule that modulates the function of synapses through protein–protein interactions. Yet, it remains unclear whether the regulation of synaptic transmission in the spinal cord by neruoligin1 contributes to the development of postoperative pain. In a rat model of postoperative pain induced by plantar incision, we conducted Western blot study to examine changes in the expression of postsynaptic membrane of neuroligin1, postsynaptic density 95 (PSD-95), and α-amino-3-hydroxy-5-methyl-4-isoxazole propionate (AMPA) receptor GluA1 and GluA2 subunits in the spinal cord dorsal horn after injury. The interaction between neuroligin1 and PSD-95 was further determined by using coimmunoprecipitation. Protein levels of neuroligin1 and GluA1, but not GluA2 and PSD-95, were significantly increased in the postsynaptic membrane of the ipsilateral dorsal horn at 3 h and 1 day after incision, as compared to that in control group (naïve). A greater amount of PSD-95 was coimmunoprecipitated with neuroligin1 at 3 h after incision than that in the control group. Intrathecal administration of small interfering RNAs (siRNAs) targeting neuroligin1 suppressed the expression of neuroligin1 in the spinal cord. Importantly, pretreatment with intrathecal neuroligin1 siRNA2497, but not scrambled siRNA or vehicle, prevented the upregulation of GluA1 expression at 3 h after incision, inhibited the enhanced neuroligin1/PSD-95 interaction, and attenuated postoperative pain. Together, current findings suggest that downregulation of spinal neuroligin1 expression may ameliorate postoperative pain through inhibiting neuroligin1/PSD-95 interaction and synaptic targeting of GluA1 subunit. Accordingly, spinal neuroligin1 may be a potential new target for postoperative pain treatment.
Introduction
The mechanisms underlying postoperative pain may differ from those of inflammatory or neuropathic pain. 1 There is hence an overwhelming need to dissect the mechanisms underlying postoperative pain in order to develop better analgesics for postoperative pain therapy. The spinal cord dorsal horn is the first central relay station in the pain pathways. In a rat model of incisional pain, previous studies demonstrated that the dorsal horn neuronal circuits shift toward a state of hyperexcitability, characterized by the enhanced neurotransmitters release and the recruitment and aggregation of postsynaptic receptors in the excitatory synapses. 2 Neurotransmitters released from the presynaptic terminal act on receptors located on the post-synaptic membrane to convey information between neurons at synapses. 3 Protein complexes that link pre- and post-synaptic membranes serve an important function in regulating neural networks. Signaling can be facilitated by adhesion molecules, which cooperate across the synaptic junction.4–8 Synaptically localized cell adhesion molecules modulate the function of synapses through protein–protein interactions, alternating signaling cascades and synaptic receptor function. The most widely described cell adhesion molecules pairing is the interface between neurexins and neuroligins that are located at pre-synaptic and post-synaptic sites, respectively. At excitatory postsynapses, neuroligin1 binds the major scaffold protein postsynaptic density 95 (PSD-95), which interacts directly with NMDA glutamate receptors (NMDAR) and indirectly with AMPA glutamate receptors (AMPAR) through binding to the auxiliary subunit stargazin.9–14 The importance of neuroligins in nervous system function is highlighted by the fact that neuroligin knock-out mice show altered NMDA-mediated synaptic responses 15 and deficits in long-term potentiation. 16
Change in transsynaptic signaling may play a critical role in the development of postoperative pain. Our previous studies showed that the synaptic targeting of AMPA receptor GluA1 subunit through the interaction between stargazin and PSD-95 in dorsal horn neurons plays an important role in the development of postoperative pain. 17 The function of neuroligin1 is vital in the transmission of trans-synaptic signaling. Binding of neurexin/neuroligin to PDZ3 domain of PSD-95 contributes to the transmission of signals between postsynaptic and presynaptic membrane.18,19 However, whether neuroligin1-mediated transsynaptic signals are implicated in the development of postoperative pain remains unclear. We hypothesize that neuroligin1 may play an important role in the postsynaptic recruitment and aggregation of AMPA receptors in dorsal horn neurons and hence contributes to postoperative pain. Conversely, disrupting the neuroligin1-mediated transsynaptic signals in dorsal horn would ameliorate postoperative pain.
Materials and methods
Animals
Adult male Sprague-Dawley rats (weigh 280–300 g) were used for all studies. Rats were housed on a 12-h light/12-h dark cycle and maintained at 21°C ± 2°C with free access to food and water. All experiments were approved by the Ethical Committee of Beijing Friendship Hospital, Capital Medical University (15–1001, Beijing, China), and were performed in compliance with the guidelines for animal experimentation of the international association for the study of pain. To avoid selection bias, we assigned animals randomly to different experimental groups by a computer-generated randomization list. The sample size for our experiments was determined based on our previous studies. 17
Plantar incision
The plantar incision was performed as described previously. 20 Briefly, all rats were anesthetized with 1.5% to 1.8% isoflurane anesthesia. Then, we made a 1-cm longitudinal incision through plantar skin and fascia of the right hind paw of rats. The underlying flexor muscle was elevated with forceps and incised longitudinally. The muscle origin and insertion remained intact. The wounded skin was closed and covered with antibiotic ointment. Sham-operated rats underwent all procedures but no incision.
Behavioral test
The paw withdrawal threshold and cumulative pain score were carried out to evaluate pain behaviors by an experimenter blind to surgery and drug treatment conditions.
The paw withdrawal threshold was tested with calibrated von Frey filaments (Danmic, USA; Aesthesio). In brief, to measure the mechanical withdrawal threshold, we applied a series of filaments vertically to an area adjacent to the wound near the medial heel for 6 s or until the animal withdrew the paw. The positive responses included rapid lifting, shaking, or licking of the incised paw. The mechanical withdrawal threshold was determined by an up-down method as described previously.21,22 To measure the cumulative pain score, the animals were placed on an elevated mesh floor and observed closely for a 1-min period every 5 min for 1 h, and a score of 0, 1, or 2 was given based on the extent of weight bearing on the incised foot. Full weight bearing of the foot (score = 0) was present if the plantar aspect of the hindpaw was blanched or distorted by the mesh. If the foot was completely off the mesh, a score of two was recorded. If the plantar aspect of the hindpaw touched the mesh without blanching or distorting, a score of one was given. The sum of the 12 scores (0–24) established the cumulative pain score.20,23 At 3 h, 1 day, and 3 days after plantar incision, rats were subjected to behavioral testing and then sacrificed for tissue harvest.
Whole-cell homogenates and postsynaptic density fractionation of proteins
The dorsal lumbar spinal cord (L3–L6) ipsilateral and contralateral to the incised paw was separated and stored at −80°C. Tissues were homogenized in whole-cell buffer (50 mM Tris-HCl (pH 7.5), 2 mM EDTA, 2 mM EGTA, 1 mM DTT, 50 mM KF, 5 mM sodium pyrophosphate, 0.1 mM sodium vanidate, 5 × 10–5 mM okadaic acid, 5 mg/L pepstatin, 5 mg/L chymostatin, 5 mg/L leupeptin, 5 mg/L aprotinin, 2% SDS). The suspension was sonicated 8 to 10 times by ultrasonic probe until the suspension was no longer turbid. To avoid overheating, the EP tube was cooled in ice bath at intervals. We collected the supernatant that was used for protein concentration.
Postsynaptic density fractionation was carried out according to previous studies with minor modifications.24,25 Tissues were homogenized in homogenization buffer (10 mM Tris-HCl (pH 7.4), 5 mM NaF, 1 mM sodium orthovanadate, 320 mM sucrose, 1 mM EDTA, 1 mM EGTA, 0.1 mM phenylmethylsulfonyl fluoride, 1 mM leupeptin, and 2 mM pepstatin A) and centrifuged at 1000 × g for 20 min at 4°C. We collected the supernatant (S1, total soluble fraction) and discarded the pellet (P1, nuclei, and debris fraction). Then, S1 was centrifuged at 10,000 × g for 20 min to produce a pellet (P2) and supernatant (S2). The P2 was lysed hypo-osmotically in ddH2O (500 μl) and centrifuged at 25,000 × g to produce pellet 3 (P3). We collected the P3 (crude synaptosomal membrane fraction) to test the protein concentration.
Western blot
Proteins (60 μg) were loaded and separated by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and then transferred onto a polyvinylidene difluoride membrane. The membrane was blocked and then incubated with primary mouse antibody to neuroligin1 (1:1000, SYSY, Göttingen, Germany) at room temperature for 2 h. Then, the membrane was incubated with horseradish peroxidase-conjugated goat anti-mouse IgG (1:4000, ZSGB-BIO, China) for 1 h. The membrane was also blocked and incubated overnight with primary rabbit antibodies to GluA1 (1:1000, Millipore, Billerica, MA), GluA2 (1:1000, Millipore, Billerica, MA), or PSD-95 (1:500, Abcam, Cambridge, UK). Then, the membrane was incubated with horseradish peroxidase-conjugated goat anti-rabbit IgG (1:1000, KPL, Boston, MA). Finally, the membranes were exposed to reagents from the Enhanced Chemiluminescence Detection Kit (Thermo, Waltham, MA) and x-ray film for visualization of protein bands. The intensity of the protein bands was quantified with densitometry. The β-tubulin was used as the loading control.
Immunofluorescence
Four rats were sacrificed at 3 h after incision and were transcardially perfused with 10 mM phosphate-buffered saline followed by 4% formaldehyde. L3–L6 spinal cord segments were placed in 15% sucrose until the tissue sink to the bottom and then replaced with 30% sucrose for 48 h at 4°C. Then, the tissues transversely cut to a 20-μm thickness. Spinal cord sections were blocked and dual stained for neuroligin1 and PSD-95. The tissues were incubated overnight in monoclonal mouse anti-neuroligin1 (1:50, Santa, Dallas, TX) with rabbit anti-PSD-95 (1:500, Abcam, Cambridge, UK). The sections were then incubated for 1 h in fluorescein-conjugated affinipure goat anti-mouse IgG (1:100, Jackson, Lancaster, PA) and rhodamine affinipure goat anti-rabbit IgG (1:100, Jackson, Lancaster, PA) and mounted in VectaShield medium (Vector Laboratories, Burlingame, CA). Confocal microscopy was performed, and images were prepared with the confocal software and Adobe Photoshop software (San Jose, CA). All images were taken from the dorsal horn that is ipsilateral to side of injury (right side).
Coimmunoprecipitation
Coimmunoprecipitation was performed as described previously with minor modifications. 26 Total protein was extracted using non-denaturing cell lysis buffer (Applygen, Beijing, China). The affinity-purified mouse neuroligin1 antibody (2 μg, Santa, Dallas, TX) was added to the protein G sepharose beads and incubated for 3 to 4 h at 4°C with gentle rotation. The complex was centrifuged at 2000 r/min for 4 min. Then, the mixture was mixed with the total protein from spinal cord homogenates (500 μg) for overnight at 4°C. The immunoprecipitates were washed three times. The proteins were collected and the proteins were separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis and transferred onto polyvinylidene difluoride membrane. The membrane was blocked and then incubated overnight with primary antibodies to neuroligin1 (1:1000, SYSY, Göttingen, Germany) or PSD-95 (1:500, Abcam, Cambridge, UK). As a positive control (input), 60 μg of total protein was loaded onto the gel. The proteins were detected with horseradish peroxidase-conjugated secondary antibody and visualized by enhanced chemiluminescence and exposure to x-ray film. Band intensity was quantified with imaging software.
Intrathecal catheter implantation
A polyethylene catheter was prepared and sterilized before insertion. Rats were subjected to 5% chloral hydrate anesthesia (300 mg/kg, i.p.). After sterile preparation, a blunt guide cannula (20 Gauge) was advanced in the narrow space between L5 and L6 intervertebral space, and the correct intrathecal localization was confirmed by a tail-flick or a paw retraction and occasionally by backflow of spinal fluid. The catheter was threaded into the guide cannula and inserted approximately 3 cm beyond the tip of the guide cannula. The catheter was sutured and tunneled under the skin to the cervical region. After surgery, rats were housed in individual cages. On the day 2 of the implantation surgery, a 15 µl of 2% lidocaine followed by saline (10 µl) was given and an immediate motor paralysis of the hind part of the animal (within 15 s) lasting for 20 to 30 min indicated a correct intrathecal location. The rats without motor or postural deficits were selected for further experiments at least five days after intrathecal implantation. All injections were followed by a 10-μl saline flush to clear the catheter.
Neuroligin1 siRNA
Purified siRNA duplexes targeting rat neuroligin1 were chemically synthesized by GenePharma Co., Ltd (Shanghai, China). The following four different siRNA sequence pairs were used: siRNA 699, 5′-GGCCUGUCAUUCAGUUUCUTT -3′ and 5-AGAAACUGAAUGACAGGCCTT -3′; siRNA2362, 5′-GGUACCUCAUCUGCAUAAUTT -3′ and 5′-AUUAUGCAGAUGAGGUACCTT -3′; siRNA2497, 5′- GCAGGAUGAUCCCAAACAATT -3′ and 5′- UUGUUUGGGAUCAUCCUGCTT -3′; and siRNA1342, 5′- GCUGACUUUAUCCCAUUAUTT -3′ and 5′- AUAAUGGGAUAAAGUCAGCTT -3′. To identify the interference efficiency of the four siRNA duplexes and screen the optimal siRNA duplexes for in vivo experiments, we carried out cotransfection experiments in vitro. Human embryonic kidney 293T cells were cultivated under standard conditions as described previously. Cotransfection experiments were performed with a pcDNA3.1/neuroligin1-green fluorescent protein (GFP) plasmid (0.3 μg) and siRNA (0.8 μg). Selective neuroligin1 siRNA or scrambled siRNA was mixed in 200 μl of OptiMEMI low-serum media with 6 μl of Lipofectamine 2000 reagent (Invitrogen, Grand Island, NY). Complexes were allowed to form for 20 min at room temperature before being added to cells. Cells were incubated at 37°C in 5% CO2 with gentle rocking for 24 h or 48 h and then harvested. Protein expression was analyzed by Western blotting. The plasmid (NlGN1-GFP) lane was used as standard lane.
The intrathecal injections of vehicle, neuroligin1 siRNA, or scrambled siRNA (negative control, 3.3 μg in 10 μl) in transfection reagent (at a ratio of 1:5, w: v) were performed twice daily for three consecutive days in rats before plantar incision.17,27 The selected dose of siRNA was based on our preliminary data.
Statistical analysis
Quantitative analysis for expression time-courses of neuroligin1, GluA1, GluA2, and PSD-95 was performed by one-way analysis of variance (ANOVA) followed by all pair-wise multiple comparison procedures using the Bonferroni test. For coimmunoprecipitation experiments, comparisons of immunoprecipitated neuroligin1 and PSD-95 between control and the 3-h time point were analyzed statistically by Student’s t test for independent samples. For comparisons of input and immunoprecipitation on immunoblots of neuroligin1 and PSD-95 among control, sham-operated, and incision groups, we used ANOVA followed by Bonferroni multiple comparison tests. The data from cell culture and pain behavioral tests were also analyzed with ANOVA followed by Bonferroni multiple comparison tests. The two-tailed tests were used since we had open outcome expectations for these experiments. SPSS 18 (SPSS Inc., Chicago, IL) was used for data analysis. All data are expressed as mean ± SEM. P < 0.05 was regarded as significant.
Results
Plantar incision increased the protein levels of neuroligin1 and GluA1 in postsynaptic membrane fraction of ipsilateral dorsal horn, but did not enhance the total protein levels of neuroligin1 in whole-cell homogenates of ipsilateral dorsal horn
Western blot study showed that the levels of neuroligin1 and GluA1 were significantly increased in the postsynaptic membrane fraction of ipsilateral dorsal horn at 3 h and 1 day after plantar incision (P < 0.05), as compared to that in control group (Figure 1(a) to (c)). The increased neuroligin1 and GluA1 expression returned to the baseline level at three days after incision. There was no significant change in the expression of GluA2 and PSD-95 in the postsynaptic membrane fraction of ipsilateral dorsal horn after plantar incision (P > 0.05, Figure 1(a), (d), and (e)).
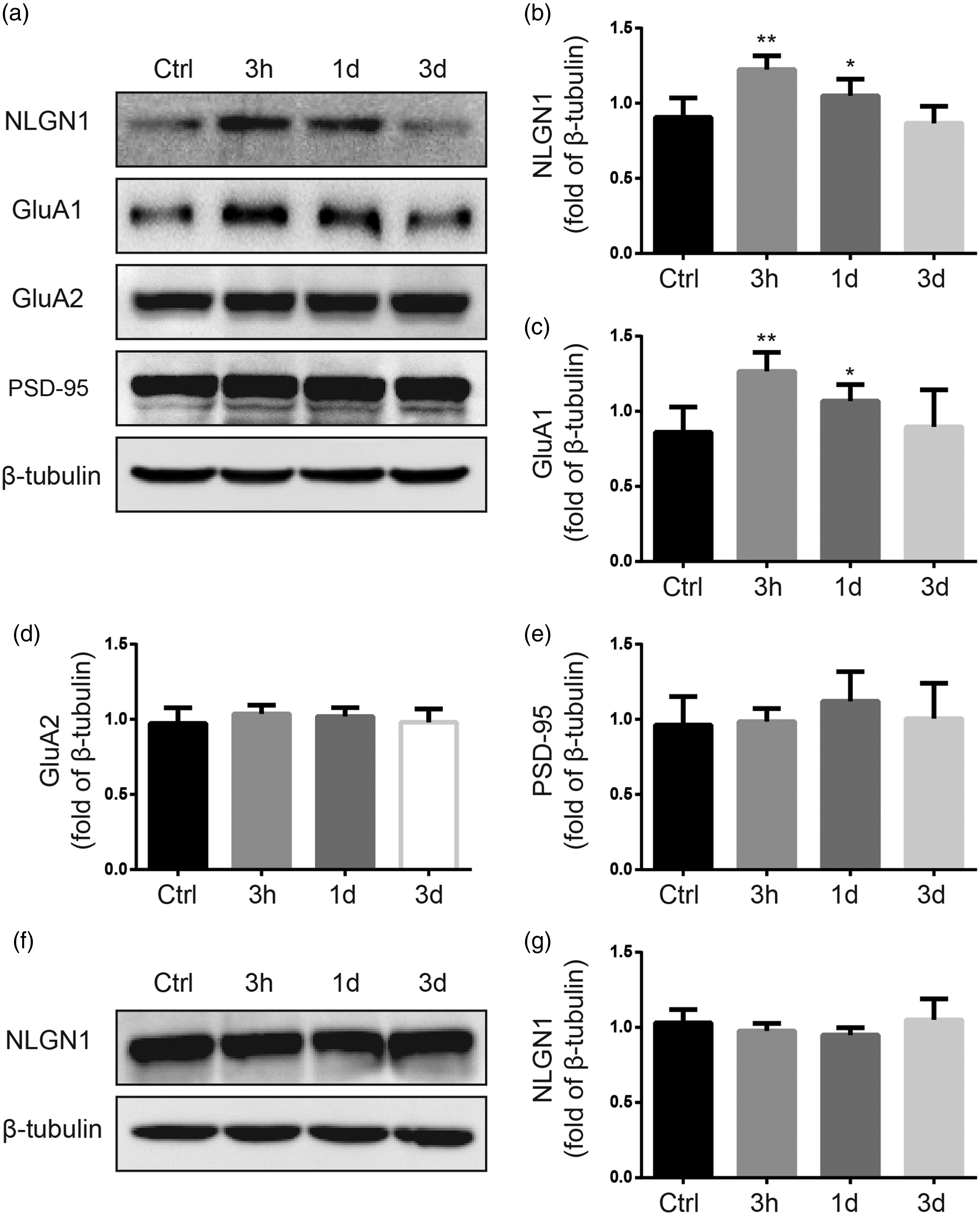
Plantar incision increased the expression of neuroligin1 and GluA1 in the postsynaptic membrane fraction of ipsilateral dorsal horn, but did not enhance the expression of total neuroligin1 in whole-cell homogenates of ipsilateral dorsal horn. (a) Representative Western blots show neuroligin1, GluA1, GluA2, and PSD-95 protein levels in the postsynaptic membrane fraction of ipsilateral dorsal horn in rats at 3 h, 1 day, and 3 days after plantar incision. (b) to (e) Quantitative analysis of band density shows that the expressions of neuroligin1 and GluA1, but not GluA2 and PSD-95 in the postsynaptic membrane fraction of ipsilateral dorsal horn, were significantly increased after incision. The β-tubulin was used as an internal reference. (f) Representative Western blot shows the expression of neuroligin1 in the whole-cell homogenates of ipsilateral spinal cord dorsal horn in rats at different time points after plantar incision. The β-tubulin was used as an internal reference. (g) Quantitative analysis of band density shows that incision did not significantly alter the expression of neuroligin1 in the whole-cell homogenates of the ipsilateral dorsal horn, as compared to that in the control group *P < 0.05, **P < 0.01, compared with the control group. n = 6 rats/time point. Ctrl: Control group (naive); NGNL1: neuroligin1; PSD: postsynaptic density.
In whole-cell homogenates of ipsilateral dorsal horn, the protein levels of neuroligin1 at 3 h, 1 day, and 3 days after incision were not significantly different from that of control group (Figure 1(f) and (g)). Additionally, in the contralateral dorsal horn, the level of neuroligin1 did not significantly change after incision (data not shown).
Neuroligin1 and PSD-95 were colocalized in the dorsal horn and showed an increased interaction after plantar incision
The neuroligin1 and PSD-95 interaction is indispensable for the transmission of trans-synaptic signals. Double-labeling immunofluorescence study showed that neuroligin1 and PSD-95 were highly colocalized in ipsilateral dorsal horn in rats at 3 h after plantar incision (Figure 2(a) to (f)).

Neuroligin1 and PSD-95 were colocalized in the dorsal horn and showed an increased interaction after plantar incision. Representative images of neuroligin1 ((a) and (d), green) and PSD-95 ((b) and (e), red) immunoreactivity in the dorsal horn of lumbar spinal cord (L5–L6). Colocalization (merge) is shown in yellow ((c) and (f)). Double-labeled neurons are indicated by arrows. (a), (b), and (c) are shown in lower magnification. (d), (e), and (f) are shown in higher magnification. Scale bars: 100 μm (a) and 50 μm (d). (g) Ipsilateral dorsal horn tissue was collected from rats at 3 h after plantar incision of the right hind paw and from control (naïve) rats. Representative immunoblots illustrate total neuroligin1 and PSD-95 protein (Input) and PSD-95 coimmunoprecipitated by neuroligin1 antibody (IP: anti-neuroligin1) or IgG (negative control). (h) Quantitative analysis shows the level of PSD-95 coimmunoprecipitated by neuroligin1 antibody. The interaction between neuroligin1 and PSD-95 was significantly enhanced at 3 h after incision. The PSD-95 was not coimmunoprecipitated with IgG. **P < 0.01 compared to the control group. n = 6 rats per group. NGNL1: neuroligin1; PSD: postsynaptic density.
We further determined whether incision changes the interaction between neuroligin1 and PSD-95 in dorsal horn. At 3 h after plantar incision, the amount of PSD-95 that was coimmunoprecipitated with neuroligin1 antibody was significantly increased in the ipsilateral dorsal horn, as compared to that of the control group (P < 0.01, Figure 2(g) and (h)). PSD-95 was not coimmunoprecipitated by IgG (Figure 2(g)), suggesting the specific interaction between neuroligin1 and PSD-95, which was enhanced in the dorsal horn after plantar incision. The interaction between neuroligin1 and PSD-95 in the contralateral dorsal horn was not significantly increased after incision (data not shown).
Validation of small interfering RNAs (siRNAs) for neuroligin1
Since siRNAs may induce selective and effective gene-silencing, we examined four siRNAs in cultured human embryonic kidney 293T cells to identify the effective one for inhibiting neuroligin1 expression. The siRNA699, siRNA2362, and siRNA2497 inhibited the expression of GFP-neuroligin1 at 24 h and 48 h after treatment (P < 0.01, Figure 3(a) and (b)). The siRNA2497 induced a greater inhibition than siRNA699 and siRNA2362. The siRNA1342 and scrambled siRNA (negative control) did not inhibit GFP-neuroligin1 expression. Accordingly, the siRNA2497 was chosen for the following in vivo study.
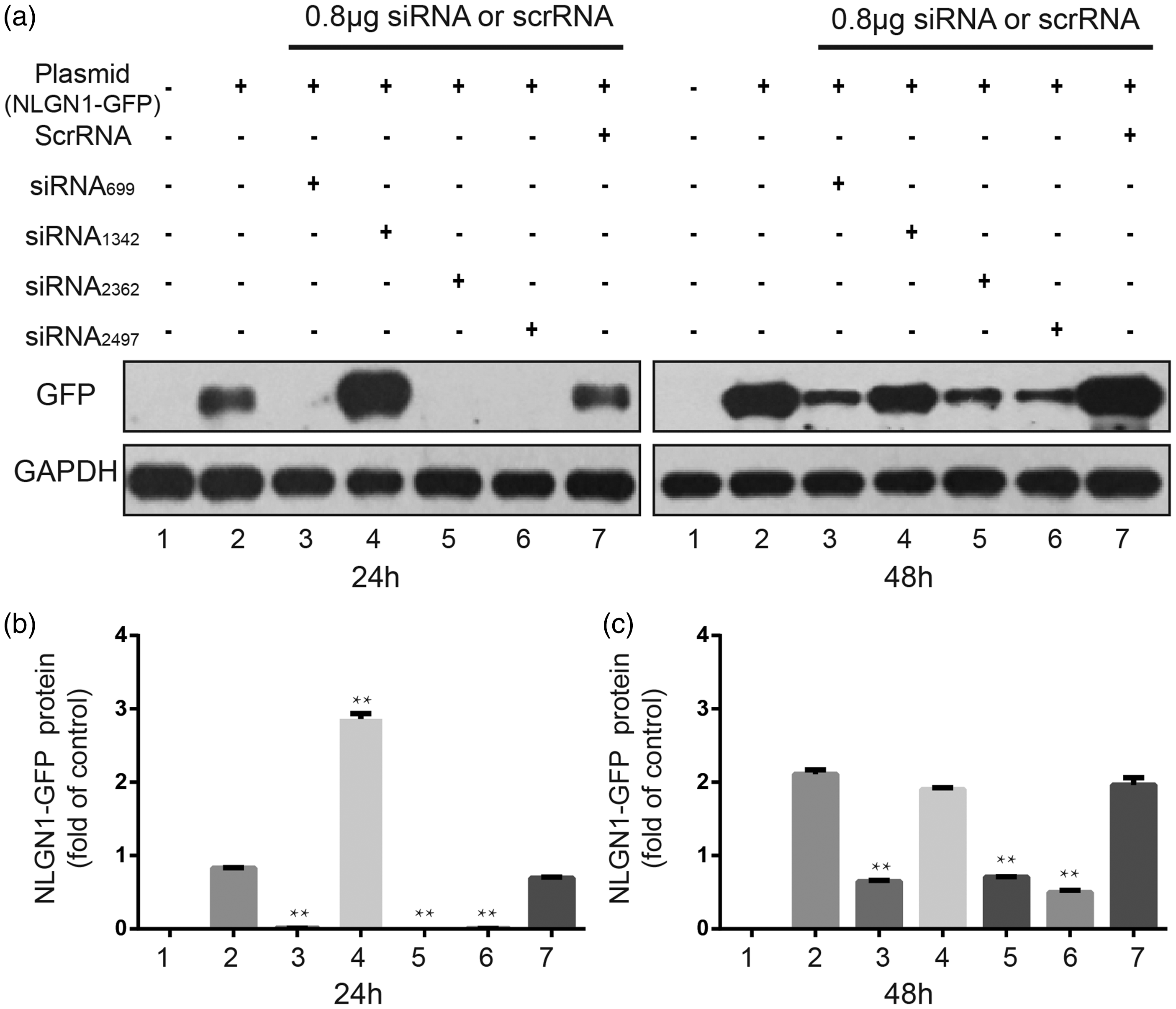
Identification of small interfering RNA (siRNA) for neuroligin1. (a) and (b) Representative immunoblots show the effects of four neuroligin1 siRNA constructs on the expression of neuroligin1-green fluorescent protein (GFP) fusion protein at 24 h and 48 h after treatment in human embryonic kidney 293 cells. (c) Quantitative analysis shows that the siRNA699, siRNA2362, and siRNA2497 decreased the expression of neuroligin1 in cultured cells at 24 h and 48 h. The siRNA2497 completely silenced neuroligin1 expression at 24 h and induced a greater inhibition than others siRNAs at 48 h. The scrambled siRNA did not affect neuroligin1 expression. **P < 0.01, compared to the control group. n = 6/group. Scr: scrambled; siRNA: small interfering ribonucleic acid; NGNL1: neuroligin1.
Intrathecal pretreatment of siRNA2497 attenuated postoperative pain in rats
We then used intrathecal delivery of siRNA2497 to determine whether downregulation of spinal neuroligin1 in vivo provides postoperative pain relief. Twenty-seven rats with intrathecal catheter were randomly divided into three experimental groups (n = 9/group) that received intrathecal vehicle, scrambled siRNA, or siRNA2497, respectively. Compared to naive and sham-operated rats, plantar incision significantly increased cumulative pain scores (P < 0.05, Figure 4(a)) and decreased the paw withdrawal threshold to mechanical stimulation (P < 0.05, Figure 4(b)). Compared to vehicle pretreatment, intrathecal pretreatment with siRNA2497 significantly reduced the cumulative pain scores and enhanced paw withdrawal threshold in rats at 3 h post-incision (P < 0.05, Figure 4). However, compared to control group, the cumulative pain scores remained higher and the paw withdrawal thresholds remained lower in the group with siRNA2497 pretreatment (P < 0.05). Pretreatment with scrambled siRNA did not change the cumulative pain scores and paw withdrawal thresholds, as compared to vehicle pretreatment (P > 0.05).
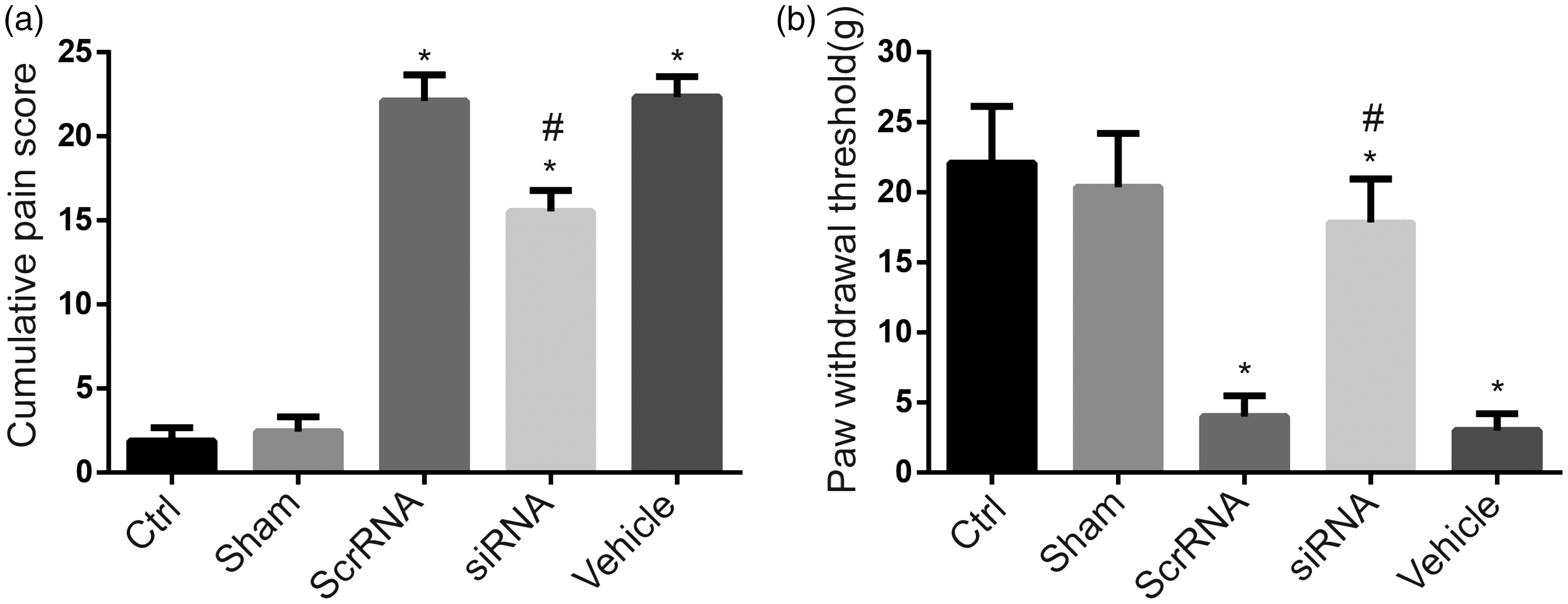
Intrathecal pretreatment with siRNA2497 decreased cumulative pain scores and mechanical hypersensitivity at 3 h after plantar incision. Pretreatment with intrathecal siRNA2497, but not scrambled siRNA, significantly reduced the cumulative pain scores and increased the paw withdrawal threshold of rats at 3 h post-incision as compared with vehicle pretreatment. *P < 0.05, compared to the control group; #P < 0.05, compared to the vehicle-pretreated group. n = 9 rats per group. Ctrl: control (naïve) group; siRNA: small interfering ribonucleic acid; Scr: scrambled.
Intrathecal siRNA2497 attenuated the upregulation of neuroligin1 and GluA1 expression after plantar incision
There was an increased expression of neuroligin1 and GluA1 in postsynaptic membrane of ipsilateral dorsal horn in scrambled siRNA-pretreated and vehicle-pretreated rats at 3 h after plantar incision, as compared to the control rats. The enhanced expression of neuroligin1 and GluA1 was significantly attenuated by siRNA2497 pretreatment (P < 0.05, Figure 5(a) to (c)). The expressions of GluA2 and PSD-95 in postsynaptic membrane were not significantly different between groups (Figure 5(d) and (e)).

Intrathecal pretreatment of rats with siRNA2497 inhibits the enhanced expression of GluA1 at postsynaptic membrane after plantar incision. (a) Representative Western blots show neuroligin1, GluA1, GluA2, and PSD-95 protein levels in the postsynaptic fraction of ipsilateral dorsal horn tissue from different groups. The β-tubulin was used as an internal reference. (b) Quantitative analysis of band density shows that pretreatment of siRNA2497 significantly suppressed the neuroligin1 expression in postsynaptic membrane 3 h after planter incision. (c) Pretreatment with intrathecal siRNA2497, but not scrRNA or vehicle, prevented the increase in GluA1 expression in the postsynaptic membrane of ipsilateral dorsal horn at 3 h after plantar incision. (d) and (e) Pretreatment with intrathecal siRNA2497 had no effects on the expression of GluA2 and PSD-95 in postsynaptic membrane. *P < 0.05, **P < 0.01, compared to the control group. n = 6 rats/group. Ctrl: control (naïve) group; siRNA: small interfering ribonucleic acid; Scr: scrambled; NGNL1: neuroligin1; PSD: postsynaptic density.
Intrathecal siRNA2497 inhibited the increased interaction between neuroligin1 and PSD-95 after plantar incision
The amount of PSD-95 that was coimmunoprecipitated by neuroligin1 antibody in ipsilateral dorsal horn of vehicle- and scrambled siRNA-pretreated groups was significantly greater than that of the sham injury group (P < 0.05, Figure 6(a) and (b)), suggesting an increased interaction between neuroligin1 and PSD-95 after plantar incision. Pretreatment with siRNA2497 significantly decreased the level of PSD-95 coimmunoprecipitated by neuroligin1 antibody (Figure 6(b)).

Intrathecal pretreatment with siRNA2497 attenuated the increased in neuroligin1–PSD-95 interaction after plantar incision. (a) Representative immunoblots illustrate PSD-95 coimmunoprecipitated by neuroligin1 antibody in the ipsilateral dorsal horn in rats that received different pretreatments. (b) Coimmunoprecipitation results show that pretreatment with siRNA2497 reduced the level of neuroligin1-PSD-95 interaction as compared to the control group. *P < 0.05, **P < 0.01, compared to the control group. n = 6 rats per group. Ctrl: control (naïve) group; IP: immunoprecipitation; NGNL1: neuroligin1; siRNA: small interfering ribonucleic acid; Scr: scrambled; PSD: postsynaptic density.
Discussion
Current study showed for the first time that plantar incision upregulated the expression of neuroligin1 and increased its interaction with PSD-95 in the postsynaptic membrane fraction of spinal cord dorsal horn. This change may subsequently lead to an increased targeting of AMPA receptor GluA1 subunit to the cellular membrane in dorsal horn neurons. Importantly, downregulation of neuroligin1 expression by intrathecal siRNA2497 before incision reduced the interaction between neuroligin1 and PSD-95 and synaptic targeting of GluA1 subunits. Furthermore, intrathecal neuroligin1 siRNA pretreatment also attenuated incision-induced guarding pain behavior and mechanical hypersensitivity.
AMPA receptors play an important role in fast excitatory synaptic transmission. 28 Previous studies showed that epidural or intrathecal administration of AMPA/kainate receptor antagonists produced analgesia in incisional pain model, suggesting an important role of spinal AMPA receptors in the development of postoperative pain hypersensitivity.29–31 Our group has been studying the regulation of spinal AMPA receptor (phosphorylation, subunits trafficking) in incisional pain sensitization. In an acute incisional pain model, we found that the phosphorylation of GluA1 subunits at Serine-831 and Serine-845 sites might be differentially regulated after incision and support a neurobiological mechanism of post-operative pain involved in phosphorylation of AMPA subunits GluA1-Ser-831, but not pGluA1-Ser-845. 32 Furthermore, we found that surgical tissue injuries enhanced the membrane translocation level of PKC gamma, but not PKC alpha, beta I, and beta II, and induced the trafficking of GluA1, but not GluA2 into neuronal plasma membrane. 33 Our group also demonstrated that surgical incision enhances the interaction between stargazin and AMPA receptor subunit GluA1, but not GluA2, and increases the surface delivery of GluA1 in dorsal horn neurons. Importantly, downregulation of stargazin with intrathecal siRNA significantly inhibited incision-induced interaction between stargazin and GluA1, and attenuated the surface delivery of GluA1 in dorsal horn. 17 Current findings show that plantar incision significantly increases the level of GluA1, but not GluA2 subunit at postsynaptic membrane fraction of the ipsilateral dorsal horn. 33 Under physiologic condition, most AMPA receptors are impermeable to Ca2+ because of the presence of the GluA2 subunit. 34 Changes in GluA1 and GluA2 subunit trafficking, which lead to rapid alterations in the composition of synaptic AMPA receptors, may play an important role in pain hypersensitivity that develops after tissue or nerve injury.35–37 Our current findings in a rat model of incisional pain suggest that an increase in Ca2+-permeable AMPA receptors in dorsal horn neurons in response to plantar incision may contribute to postoperative pain by strengthening the excitatory synaptic transmission at spinal level. It has been reported that acute peripheral inflammatory stimulation by CFA or 5% formalin can induce spinal long-term potentiation very early after stimulation onset (within 3 h after subcutaneous injection of CFA or formalin into plantar). 38 This will further provide evidence to demonstrate the functional significance of the present study. Previous studies also showed that neurogenic inflammation induced by capsaicin, carrageenan, and formalin injection also increased the membrane insertion of GluA1, but not GluA2 subunits in the dorsal horn,35,39–42 which is in line with current findings.
Neurexins and neuroligins are important transmembrane adhesion proteins in the regulation of synapse function.11,43–46 The expressions of neuroligin1 and neuroligin2 in the central nervous system are restricted to excitatory and inhibitory synapses, respectively.18,47–49 Neuroligin1 is enriched at the postsynaptic densities of excitatory synapses containing the scaffolding protein PSD-95. In contrast, neuroligin2 is primarily located at glycinergic and GABAergic inhibitory synapses, where it colocalizes with the scaffolding protein gephyrin. 6 Because of their specific synaptic localization and function, neuroligins control the balance between excitatory and inhibitory synapses.18,50,51 Under hyperalgesic states, synaptic plasticity in the dorsal horn shifts the balance toward excitation. 52 However, less is known whether neuroligin1 dysfunction is implicated in the imbalanced excitatory/inhibitory synaptic transmission in dorsal horn during postoperative pain. In the current study, we provide novel evidence that plantar incision increases the expression of neuroligin1, at postsynaptic membrane fraction, but not the whole homogenates of the ipsilateral dorsal horn after plantar incision. Recent studies have shown that nerve injury did not alter the neuroligin1 expression in the dorsal horn.47,52 It has been demonstrated that changes in synaptic strength can alter neuroligin1 intracellular transport. Competitive PSD-95 peptide and neuroligin 1 C-terminal mutagenesis can specifically alter expression of neuroligin1 on membrane and interfere its internalization. 11 Our results suggest that the trafficking of neuroligin1 to postsynaptic membrane in dorsal horn neurons may occur after plantar incision, but the underlying mechanisms remain to be investigated.
Although the level of PSD-95 in postsynaptic membrane fraction was unchanged after plantar incision, coimmunoprecipitation revealed that incision enhances its interaction with neuroligin1. According to previous studies,53–55 pre-existing PSD-95 “slots” promoted the synaptic recruitment of Stargazin/AMPARs complexes in long-term potentiation. Our study showed that plantar incision enhanced the expression of neuroligin1 in postsynaptic membrane and the binding between neuroligin1 and pre-existing PSD-95. The total levels of PSD-95 in postsynaptic membrane were unchanged after planter incision. Additionally, double-labeling immunofluorescence study indicates that neuroligin1 and PSD-95 are colocalized in dorsal horn neurons at 3 h post-incision. This morphological evidence suggests a physiological basis for the interaction between neuroligin1 and PSD-95 in the same cell. Lin et al. 52 also showed the colocolization of neuroligin1 and PSD-95 in dorsal horn neurons in neuropathic rats. At excitatory synapses, neuroligin-1 binds to the third PDZ (PSD-95/Dlg/ZO-1) homology domain of the major scaffold protein PSD-95, which interacts directly with NMDA receptors and indirectly with AMPA receptors through binding to the auxiliary subunit stargazin.9–14 Our previous study demonstrated that the interaction between PSD-95 and stargazin can regulate the clustering of AMPA receptor GluA1 subunits in the dorsal horn in response to incisional noxious stimuli. 17 Neuroligin1 has been implicated in diverse forms of synaptic plasticity.56,57 Overexpression of neuroligin1 increases evoked synaptogenesis and excitatory postsynaptic currents,15,58 indicating that the enhanced expression of neuroligin1 may contribute to the facilitated excitatory synaptic transmission. Taken together, these findings suggest that plantar incision enhances the expression of neuroligin1, which leads to the increased interaction between neuroligin1 and PSD-95 in postsynaptic dorsal horn neurons. Subsequently, PSD-95 may indirectly enhance the synaptic targeting of GluA1 subunits through the interaction with stargazing. 17 Together, these findings support the critical role of neuroligin1/PSD-95 interaction in the incision-induced synaptic targeting of spinal AMPA receptor GluA1 subunits. As we have not examined in a chronic postsurgical pain model (e.g., skin/muscle incision and retraction model), the interpretation of current findings is limited to the early or acute postoperative pain condition.
Neuroligin1 is involved in chronic pain, such as neuropathic pain. 52 Current findings suggest that neuroligin1-/PSD-95-mediated synaptic targeting of spinal AMPA receptor GluA1 subunits may also play an important role in the development of incisional pain sensitization. We think that the increased interaction between neuroligin1 and PSD-95 may contribute to the development of central sensitization, and may not only present in chronic pain conditions. But the mechanism underlying the transition of post-operative incision pain from acute phase to chronic phase remains unclear.
The suppression of neuroligin1 by siRNA produced a significant pain inhibition after incision, suggesting that neuroligin1 is an important player in the development of postoperative pain. Meanwhile, knockdown of neuroligin1 expression with siRNA attenuated the increased interaction between neuroligin1 and PSD-95 and inhibited the synaptic targeting of GluA1 subunits after incision. However, the expressions of PSD-95 and GluA2 subunits at postsynaptic membrane fraction were unchanged. The expression ratio of synaptic GluA1 to GluA2 may greatly affect the efficacy and strength of AMPA receptor-mediated excitatory synaptic transmission.59–62 GluA1-deficient mice exhibited fewer Ca2+-permeable AMPA receptors and a decrease in AMPA current density in dorsal horn neurons, which was associated with a decreased inflammatory hyperalgesia.63,64 In contrast, GluA2-deficient mice exhibited an increased spinal Ca2+-permeable AMPA receptors and an increased inflammatory hyperalgesia. 65 Thus, neuroligin1 siRNA may cause a reduction in the number of Ca2+-permeable AMPA receptors at postsynaptic dorsal horn neurons, which, in turn, decreases synaptic strength and leads to attenuated spinal nociceptive transmission. A recent study in mice lacking 4E-binding protein 1 showed that an enhanced expression of neuroligin1 is associated with the increased excitatory synaptic input into dorsal horn neurons and facilitation of synaptic potentiation. 66 Furthermore, genetic suppression of neuroligin1 normalized the increased excitatory synaptic activity and reversed mechanical hypersensitivity. 66 Therefore, neuroligin1-mediated trans-synaptic signaling is a critical to the development of pain hypersensitivity.
In summary, our findings suggest that neuroligin1-/PSD-95-mediated synaptic targeting of AMPA receptor GluA1 subunits may be involved in postoperative pain. Targeting neuroligin1-mediated trans-synaptic signaling at spinal level may provide a new therapy to inhibit postoperative pain.
Footnotes
Author Contributions
WY and TM conceived the project and supervised all experiments. GRJ and WY designed the project. GRJ, LXY, and XZJ performed the animal models, conducted behavioral experiments and siRNA experiments. LHL, SYQ, and MDX carried out the western, coimmunoprecipitation, and immunofluorescence. GRJ, LHL, LXY, GY, LJF, and WY analyzed the data. GRJ, WY, and GYT wrote the manuscript. All authors read and approved the final manuscript.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the National Natural Science Foundation of China (81400909, 81571065, 81428008, 81171181, Beijing, China) and Natural Science Foundation of Beijing (7152056, Beijing, China).