
Abstract
N-acetylaspartylglutamate (NAAG) is the third most prevalent and widely distributed neurotransmitter in the mammalian nervous system. NAAG activates a group II metabotropic glutamate receptor (mGluR3) and is inactivated by an extracellular enzyme, glutamate carboxypeptidase II (GCPII) in vivo. Inhibitors of this enzyme are analgesic in animal models of inflammatory, neuropathic and bone cancer pain. NAAG and GCPII are present in the locus coeruleus, a center for the descending noradrenergic inhibitory pain system. In the formalin footpad model, systemic treatment with GCPII inhibitors reduces both phases of the inflammatory pain response and increases release of spinal noradrenaline. This analgesic efficacy is blocked by systemic injection of a group II mGluR antagonist, by intrathecal (spinal) injection of an alpha 2 adrenergic receptor antagonist and by microinjection of an α-amino-3-hydroxy-5-methylisoxazole-4-propionic acid (AMPA) receptor antagonist directly into the contralateral locus coeruleus. Footpad inflammation increases release of glutamate in the contralateral locus coeruleus and systemic treatment with a GCPII inhibitor blocks this increase. Direct injection of GCPII inhibitors into the contralateral or ipsilateral locus coeruleus reduces both phases of the inflammatory pain response in a dose-dependent manner and the contralateral effect also is blocked by intrathecal injection of an alpha 2 adrenergic receptor antagonist. These data support the hypothesis that the analgesic efficacy of systemically administered GCPII inhibitors is mediated, at least in part, by the contralateral locus coeruleus via group II mGluR, AMPA and alpha 2 adrenergic receptors.
Keywords
Introduction
N-acetylaspartylglutamate (NAAG) is the most prevalent peptide co-transmitter in the mammalian nervous system.1,2 NAAG is inactivated by an extracellular glial peptidase, glutamate carboxypeptidase II (GCPII). Inhibitors of GCPII elevate synaptic levels of NAAG3–5 resulting in NAAG activation of presynaptic type 3 metabotropic glutamate receptors (mGluR3) 2 and the inhibition of transmitter release.3,5–8 NAAG also has been found to act as an antagonist at N-methyl-D-aspartate (NMDA) receptors 9 that contain NR2A and 2B subunits but not other NMDA receptors.10,11 Inhibitors of GCPII and GCPII knock-out mice have been studied in several animal models of clinical disorders12–16 leading to the hypothesis that activation of group II metabotropic glutamate receptors by elevated levels of NAAG has clinical potential. In the present paper, we further test this hypothesis in an animal model of inflammatory pain.
IP NAAG peptidase inhibitor delivery studies (a) and contralateral and ipsilateral NAAG peptidase inhibitor delivery studies (b).

Material and methods
This research was performed according to a protocol approved by the Institutional Animal Care Committee of Kumamoto University. Male Sprague–Dawley rats (250–300 g; Japan SLC, Inc., Shizuoka, Japan) were housed in groups of two, maintained on a 12-h dark–light cycle, and permitted food and water ad libitum. Animals were handled on arrival and housed for at least three days before testing. Animals were euthanized immediately after behavioral or microdialysis studies.
Drugs
The GCPII inhibitors (S)-2-[3-[(S)-1-carboxy-3-methylbutyl]ureido]pentanedioic acid (ZJ43, molecular weight = 304.3) 28 and 2-(phosphonomethyl) pentanedioic acid (2-PMPA, molecular weight = 314) 29 were from Reagents4Research LLC (Hangzhou, CN). The group II metabotropic glutamate receptor (mGluR2/3) antagonist, LY341495, 30 was from Tocris Cookson Ltd (Bristol, UK). The alpha 2 adrenergic receptor antagonist, idazoxan, and the AMPA receptor antagonist, 6-cyano-7-nitroquinoxaline (CNQX), were from Sigma (St. Louis, MO).
LC cannulae placement for microinjection
Implantation of the injection cannulae into the LC was performed under halothane anesthesia. Rats were placed in a stereotaxic apparatus and stainless steel 26 G thin wall guide cannulae (C315G, Plastics One, VA) were stereotaxically placed at the level of the ipsilateral or contralateral LC through a burr hole (AP: −10.0 mm, L: 1.4 mm, H: 8.0 mm from Bregma). Guide cannulae were affixed to the skull with stainless steel screws and cranioplastic cement. To confirm whether the effects of injected drugs were site specific, we examined the effects of drugs that were injected via the guide cannula that was placed at 2.0 mm caudal and 1.6 mm lateral to the original cannula (AP: −12.0 mm, L: 3.0 mm, H: 8.0 mm from Bregma). Mefenamic acid (32.5 mg, Daiichi-Sankyo, Tokyo, Japan) was orally administered two times/day at 0–2 days after surgery for postoperative pain control. The formalin tests were performed seven days after cannulae implantation. All animal displayed normal feeding and drinking behaviors post-operatively. Rats showing neurological deficits after cannulae implantation were not studied.
For the LC microinjections, drugs were delivered 10 min before formalin treatment in a total volume of 0.5 µl over a period of 60 s using a microsyringe pump (EP-60, EICOM, Japan) and 30 G stainless steel internal cannula (C315G, Plastics One, VA) connected via a polyethylene tube to a 10 -µl Hamilton syringe.
At the completion of the experiment, 0.5 µl of India ink was injected through the internal cannula 10 min before rats were euthanized. The brains were fixed with formalin, and coronal tissue sections were Nissl stained to confirm the proper injection site (Figure 7). Only the rats whose microinjection site was located within LC were included in the results.
Behavioral test
ZJ43 and 2-PMPA were injected systemically (IP) or directly into contralateral or ipsilateral LC 10 min before the formalin injection into the footpad. To test the mechanisms of action of the NAAG peptidase inhibitors, one of the following drugs was injected 10 min before the injection of the GCPII peptidase inhibitor: type 2/3 metabotropic glutamate receptor (mGluR2/3) antagonist, LY341495 (300 µg IP); alpha 2-adrenoreceptor antagonist, idazoxan (30 µg intrathecal [IT]); AMPA receptor antagonist, CNQX (0.3 µg microinjection into the contralateral LC).
Formalin test
In the formalin test, 50 µl of 5% formalin was injected subcutaneously (SC) into the dorsal surface of the hind paw with a 26-gauge needle under brief halothane anesthesia. Within 1 min after formalin injection, spontaneous flinching of the injected paw could be observed. Flinching was readily discriminated as a rapid and brief withdrawal or flexion of the injected paw. This pain-related behavior was quantified by counting the number of flinches for 1 min periods at 5 min intervals from 0 to 60 min after the injection. Two phases of spontaneous flinching behavior were observed: an initial acute phase (phase 1: during the first 6 min after the formalin injection) and a prolonged tonic phase (phase 2: beginning about 10 min after the formalin injection).
Intrathecal microdialysis and assay of noradrenaline level in the lumbar spinal cord
Rats were anesthetized with halothane and placed in a stereotaxic apparatus for surgical implantation of a microdialysis probe. The intrathecal microdialysis probe (exposed tip, 10 mm, Cut-off of 50 kDa; EICOM, Japan) was passed caudally 7.5 cm from the cisterna magnum to the lumbar enlargement. After surgery, each rat was individually housed and the probe was perfused at 1 µl/min with artificial cerebrospinal fluid. The rat was allowed to recover for 24 h before footpad formalin injection.
Microdialysis was carried out in conscious and freely moving rats. During the experiment, the probe was perfused at 2 µl/min with artificial cerebrospinal fluid, and dialysate samples were collected every 15 min. Three baseline fractions were collected before the 2-PMPA or saline injection. Fifteen minutes after the pretreatment of 2-PMPA (50 mg/kg IP) or saline, formalin (5%, 50 µl) was injected into the dorsal side of the left hind-paw and dialysate samples were collected during the formalin test. Samples were subsequently analyzed for noradrenaline.
Noradrenaline in the microdialysate samples was analyzed by reverse phase high performance liquid chromatography (HPLC) and electrochemical detection (ECD-300, EICOM, Japan). A reversed-phase column (EICOMPAK, CA-5ODS, 2.1 × 150 mm, EICOM) was used and the mobile phase for detection of noradrenaline composed of 0.1 M phosphate buffer solution (PBS) and 50 mg/ml EDTA·2Na with methanol (95:5 v/v) in water adjusted to pH 6.0 with 400 mg/l 1-octanesulfonic acid (sodium salt). The flow rate was 0.23 ml/min (EP-300, EICOM). The column temperature was maintained at 25℃ and the applied potential was set at +450 mV (ATC-300, EICOM). Quantification was obtained from standard curves.
LC microdialysis and assay of glutamate level
Rats were anesthetized with 10% chloral hydrate (0.3 mg/kg) and placed in a stereotaxic apparatus for surgical implantation of a guide cannula. The guide cannula (EICOM) was positioned to the ipsilateral and contralateral LC according to the coordinates (AP: −10.0 mm, H: 6.8 mm, L: 1.4 mm). The guide cannula was secured to the skull with dental cement anchored by two stainless steel screws. After surgery, each rat was individually housed and allowed to recover for 24 h before microdialysis and footpad injection.
Microdialysis was carried out on conscious and freely moving rats. Rats were lightly anesthetized with isoflurane to facilitate manual insertion of the microdialysis probe into the guide cannula. The stylet in the guide cannula was replaced with the microdialysis probe (exposed tip, 2.0 mm; cut-off of 50 kDa; EICOM). The probe was perfused at 2 µl/min with artificial cerebrospinal fluid. After at least 2 h to reach equilibration, dialysate samples were collected every 15 min. Three baseline fractions were collected before 2-PMPA or saline injection and the average of these values was taken as the “100% of control” value for the glutamate levels for each rat. Pretreatment with 2-PMPA (50 mg/kg IP) or saline occurred 15 min before formalin injection (5%, 50 µl) into the dorsal side of the left hind-paw during microdialysis. At the completion of each experiment, rats were euthanized and brains were removed, fixed in 10% formalin and 30 µm coronal sections prepared to confirm probe placement.
Glutamate in the microdialysate samples was derivatized with o-phthalaldehyde (OPA, Sigma, USA) and analyzed by reverse phase HPLC and electrochemical detection (EICOM). A reversed-phase column (Zorbax, SB-C18, 4.6 × 250 mm, 5 µm, Agilent) was used with a mobile phase for detection of glutamate composed of 0.1 M citric acid-sodium acetate and 0.05 mM EDTA with 12% methanol (v/v) in water adjusted to pH 3.8 with 1 M citric acid. The flow rate was 1.0 ml/min. The column temperature was kept at 30℃ and the applied potential was set at +850 mV. Quantification was obtained from standard curves prepared over a glutamate range of 0.1–20 μM.
Immmunohistochemistry and antibodies
The NAAG-specific antisera were prepared and immunohistochemistry was performed as previously described.31,32 Briefly, polyclonal NAAG antisera were purified by negative-affinity adsorption against protein-bound N-acetylaspartate and formalin-fixed rat liver tissue. Rats were transcardially perfused with 6% 1-ethyl-3-(3-dimethylaminopropyl) carbodiimide (EDAC) and 6% DMSO with 1 mM N-hydroxysuccinimide. Brains were post fixed with 4% paraformaldehyde, saturated with 30% sucrose, frozen and sectioned (20 µM). Endogenous peroxidase was blocked by 30 min incubation with 50:50 methanol water containing 3% H2O2. Sections were treated with 2% normal goat serum prior to incubation with purified anti-NAAG rabbit serum (1:2000). Antibodies were visualized with peroxidase-labeled avidin-biotin complex (Vectastain, Vector Labs, Burlingame, CA) and developed with H2O2 as substrate for diaminobenzidine as chromogen. Control tissue sections treated with NAAG-BSA blocked antibody exhibited no specific reaction product above background. No specific staining was observed in sections that were fixed with formalin rather than EDAC. These antibodies fail to cross react with glutamate, N-acetylaspartate, N-acetylglutamate, aspartate or GABA coupled to BSA with EDAC.31,32 Images were acquired on an Olympus BX51 microscope and DP71 camera and were adjusted for contrast, brightness and white balance using PC based software (Adobe Systems).
Retrograde neuron tracing
Rats were anesthetized by isoflurane. Standard sterile technique was performed throughout the surgery. The skin was shaved and an incision was made over the thoracic spine at T10-L2 level. After the incision of the dermal and subdermal tissues at the midline, paravertebral muscles were dissected bluntly exposing the lamina bilaterally. Complete laminectomies were performed, exposing the spinal cord at T12-L1. The retrograde tracer (50 nl, Fluorospheres, carboxylate-modified microspheres, 0.04 µM, red-orange fluorescent, Molecular Probes, OR) was injected unilaterally into the dorsal funiculus by using an auto nanoliter injector (Nanoject II, Drummond Scientific Company, PA) through a glass micro pipette (tip diameter: 20 µM) at T12-L1 level spinal cord. The wounds were then closed with 3/0 silk sutures. After seven days of survival to allow for sufficient transport of tracers, tissue sections were prepared as described below and examined under confocal laser scanning microscopy (FV 1200, Olympus, Tokyo, Japan).
Preparation of tissue sections
The rats were anesthetized deeply with halothane and transcardially perfused with saline to wash blood out of the cerebral vessels, followed by 4% paraformaldehyde in 0.1 M PBS. The brain was removed immediately, post-fixed overnight at 4℃ in the same fixative, and then stored in 30% sucrose in PBS overnight. Coronal sections were cut with a microtome at 50 µm.
DBH immunohistochemistry
After immersion in PBS containing 5% normal horse serum and 0.3% Triton X-100 for 1 h, tissue sections containing the LC were incubated with rabbit antibody to dopamine β-hydroxylase (DBH) (1:1000; Merck Millipore, Darmstadt, Germany) diluted with PBS containing 5% normal horse serum and 0.3% Triton X-100 for 24 h at room temperature. The sections were then incubated at room temperature for 3 h with Alexa Fluor 488 donkey anti-rabbit IgG (1:100; Molecular Probes) in PBS containing 5% normal horse serum and 0.3% Triton X-100 and cover slipped with SlowFade Diamond Antifade mountant (Molecular Probes). Sections were photographed under confocal laser scanning microscopy.
Statistical analysis
The time-response data are presented as the mean flinches (±SEM) per minute for the periods between 1–2 and 5–6 min after formalin treatment and then for 1 min periods at 5 min intervals up to 60 min. The data from phase 1 (0–6 min) and phase 2 (10–60 min) observations were considered separately. In each case, the cumulative instances of formalin-evoked flinches during the phase 1 and phase 2 were calculated for each rat. For the ZJ43 dose response study, one-way analysis of variance (ANOVA) was used with Tukey post-hoc test. The effects of 2-PMPA injected into either contralateral or ipsilateral LC to the formalin injected side were compared with those of saline.
Microdialysis data are expressed as percentage of basal values (calculated as means of the three samples before injections). The basal concentrations of either lumbar enlargement noradrenaline or LC glutamate in the dialysates, uncorrected for recovery, were 0.35 ± 0.25 nM or 1.17 ± 0.18 µM, respectively. The values were expressed as a % of baseline level for each rat and the mean and standard error were determined for each treatment group. All data were given as mean ± standard error of the mean (SEM) and not corrected for “recovery” of the dialysis procedure. Two-factor ANOVA for repeated measurements was used to examine the possibility of significant differences (p < 0.05) between groups followed by a Student’s t-test for every time-point.
Results
NAAG in the LC
In the rat brainstem, immunoreactivity for NAAG in the LC was among the strongest of all neuronal groups (Figure 1(a)). The cell bodies of the principal neurons of the LC express strong NAAG immunoreactivity suggesting that the peptide is coexpressed with noradrenaline in the descending spinal pathway (Figure 1(b)). Additionally, small interneuron-like cells in the LC also express NAAG immunoreactivity, a result that suggests that the peptide might regulate the activity of the principal neurons within the LC.
NAAG immunohistochemistry in the rat brainstem. Highly purified antibodies to protein-coupled NAAG show NAAG localization in the brainstem (a). Box in (a) indicates the area of higher magnification in (b). IV: 4th ventricle; LC: locus coeruleus; MeV: mesencephalic nucleus of the trigeminal nerve. Bar: 800 µm (a) and 100 µm (b).
Analgesic effect of IP-injected 2-PMPA
Consistent with our previous studies,19,22 IP injection of 50 mg/kg 2-PMPA attenuated the formalin-induced agitation behavior as compared with IP saline injected rats (p < 0.01) (Figure 2). The effect of 2-PMPA was blocked by IP injection of 1 mg/kg of the group II mGluR antagonist LY341495 (p < 0.01); 1 mg/kg of LY341495 itself had no significant effect on the formalin-induced agitation behavior.
Effect of IP injection of 50 mg/kg 2-PMPA in the rat formalin test. 1 mg/kg LY341495 and 2-PMPA were injected IP 20 min and 10 min, respectively, before the formalin injection. IP 2-PMPA attenuated the formalin-induced behavior as compared with saline treated rats (p < 0.001). The effect of 2-PMPA was antagonized by IP LY341495 (p < 0.01). IP LY341495 had no effect as compared with saline treated rats (p > 0.1). Ordinate: number of flinches per min; abscissa: time after formalin administration (min). Data in this and subsequent figures are presented as the mean ± SEM.
Inflammation-induced glutamate release in the LC
Formalin injection into the rat footpad stimulated a significant increase in glutamate levels in the contralateral, but not the ipsilateral, LC (p < 0.05 by two-way ANOVA, Figure 3(a)). This increase was substantially reduced by preinjection with 50 mg/kg 2-PMPA (IP) (p < 0.05 by two-way ANOVA, Figure 3(b), same data are plotted for saline contralateral LC in Figure 3(a) and (b)). Since basal and stimulated extracellular levels of glutamate are nearly 10-fold greater than NAAG,
3
this decrease in glutamate levels can be attributed to a decrease in synaptically released glutamate rather than a decrease in glutamate released from NAAG. 2-PMPA (IP) had no significant effect on the basal level of glutamate in the ipsilateral LC (p > 0.1, Figure 3(c), same data are plotted for saline ipsilateral LC in Figure 3(a) and (c)).
Microdialysis sampling for glutamate release in the LC. Glutamate levels are expressed as a percent of the three baseline samples for each animal. (a) Effect of formalin on glutamate release in ipsilateral and contralateral LC. There was a significant effect on glutamate levels in the 45–90 min dialysate samples. Formalin significantly increased glutamate release in the contralateral LC but not the ipsilateral LC. (b) Effect of 2-PMPA (50 mg/kg, i.p.) or saline on formalin-induced glutamate release in contralateral LC. There was a significant main effect on glutamate levels in the 45–115 min dialysate samples. 2-PMPA significantly decreased formalin-induced glutamate release in the contralateral LC as compared with saline group. The same saline contralateral LC data were used in (a) and (b). (c) Effect of 2-PMPA (50 mg/kg, i.p.) or saline on formalin-induced glutamate release in ipsilateral LC. 2-PMPA had no effect on the release of glutamate in the ipsilateral LC after formalin injection.
Inflammation induces release of spinal noradrenaline
Rats injected in the footpad with formalin + IP saline had a greater level of spinal noradrenaline release than rats treated with IP saline but not formalin (p = 0.006, Figure 4(a)). Preinjection of 50 mg/kg 2-PMPA (IP) significantly increased the release of noradrenaline that followed formalin treatment (Figure 4(a), p < 0.001). IP 2-PMPA alone also induced a smaller but significant increase in spinal noradrenaline release relative to the IP saline alone treatment (p < 0.001).
Role of noradrenaline in the lumber CSF during rat formalin test. Upper panel (a) shows the microdialysis data for noradrenaline release into the lumber CSF during formalin test. Either formalin injection itself or 2-PMPA (50 mg/kg, IP) itself enhances noradrenaline release equally. IP injection of 2-PMPA 15 min before the formalin injection-induced significant noradrenaline release as compared with 2-PMPA itself or formalin itself. Samples were collected over 15 min intervals. Ordinate: % of the three baseline samples for each animal; abscissa: time after drug administration (min). Lower panel (b) shows the effect of IT idazoxan (alpha 2-adrenergic receptor antagonist) on the behavioral response to IP 2-PMPA in the rat formalin test. 2-PMPA was injected IP 10 min before the formalin injection. IP saline + IT idazoxan decreased the phase 1 flinching response but not the phase 2 response in the rat formalin test relative to IP saline alone. Ordinate: number of flinches per min; abscissa: time after formalin administration (min).
The analgesic effect of IP 2-PMPA on phase 2, but not phase 1, of the formalin footpad test was antagonized by IT idazoxan (phase 1: p = 0.73; phase 2: p = 0.0002 by t-test, Figure 4(b)). IT idazoxan with saline had a significant effect on phase 1, but not phase 2 of the formalin-induced agitation behavior induced by paw formalin injection as compared with IP saline injected rats (phase 1: p = 0.003; phase 2: p = 0.112 by t-test). While these data suggest that spinal noradrenaline mediates the effect of systemic 2-PMPA on the phase 2 response, these data do not permit interpretation of its role in the phase 1 response inasmuch as IT idazoxan itself blocked formalin-induced phase 1 agitation.
2-PMPA-induced analgesia blocked by CNQX
Injection of the AMPA receptor antagonist CNQX into the contralateral LC antagonized the analgesic effect of IP injected 2-PMPA (50 mg/kg, IP) on the phase 2 response (p < 0.002) (Figure 5). CNQX into the ipsilateral LC had no significant effect on the formalin-induced agitation behavior as compared with rats with saline injected into LC (phase 1: p = 0.920; phase 2: p = 0.694; data not shown).
Effect of AMPA antagonist in the contralateral LC. The AMPA receptor antagonist, CNQX (0.3 µg), was injected into the LC contralateral to the formalin treated footpad 10 min before 2-PMPA (50 mg/kg, IP). Formalin was injected into the footpad (zero min on figure) 10 min after 2-PMPA. CNQX blocked the analgesic effect of 2-PMPA on the phase 2 response (p < 0.01) but did not affect the phase 1 response.
Retrograde tracing from spinal cord to DBH positive neurons in the LC
Fluorospheres injected unilaterally at the T12-L1 level of the spinal cord were observed in DBH positive neurons both ipsilateral and contralateral to the tracer injected side of the spine indicating that spinal nociceptive neurons were innervated by both ipsilateral and contralateral LC neurons (Figure 6).
Retrograde tracer image in LC 7 days after injection into spinal cord. The retrograde tracer was injected unilaterally into the lumbar dorsal funiculus by using an auto nanoliter injector. The retrograde tracer was observed in DBH positive neurons (LC neurons) ipsilateral and contralateral to the tracer injected side. Injection site in the LC. Spread of 0.5 µl of India ink injected through the internal cannula placed at AP: −10.0 mm, L: 1.4 mm and H: 8.0 mm from Bregma. Section was Nissl’s stained to confirm the injection site. LC was located within the India ink spread region. Scale bar: 200 µm
Analgesic effects of ZJ43 and 2-PMPA injected into LC
Microinjection of the GCPII inhibitor ZJ43 into the LC (Figure 7) either contralateral or ipsilateral to the formalin-injected side significantly decreased the early (phase 1, contralateral: p < 0.05; ipsilateral: p < 0.001) and late (phase 2, contralateral: p < 0.005; ipsilateral: p < 0.001) phase responses (Figure 8) in a dose-dependent manner (Figure 9). When 150 µg of ZJ43 was injected via a guide cannula that was placed 2.0 mm caudal and 1.6 mm lateral (AP: −12.0 mm, H: 8.0 mm, L: 3.0 mm from Bregma) to the LC cannula (AP: −10.0 mm, H8.0 mm, L1.4 mm from Bregma), ZJ43 had no effect on the flinching behavior induced by paw formalin injection as compared with the effect of ZJ43 microinjected via a LC cannula (Figure 8).
Effect of 150 µg of ZJ43 and 150 µg of 2-PMPA microinjected into either contralateral LC or ipsilateral LC to the formalin injected side. ZJ43 or 2-PMPA or saline were microinjected into LC 10 min before the formalin injection. 300 µg (1 mg/kg) of LY341495, a type 2/3 metabotropic glutamate receptor (mGluR2/3) antagonist, was injected IP 10 min before the ZJ43 or 2-PMPA microinjection to verify the effect of ZJ43 or 2-PMPA was mediated by the activation of mGluR3. For the LC microinjection, drugs were injected at AP: −10.0 mm, H: 8.0 mm, L: 1.4 mm from Bregma. Formalin injection-induced two phases of hindpaw flinching in the saline injected control rats. Microinjection of ZJ43 or 2-PMPA into either contralateral LC or ipsilateral LC reduced the formalin-induced response and the effect of ZJ43 or 2-PMPA injected into contralateral LC, but not ipsilateral LC, was reversed by IP LY341495. Either ZJ43 or 2-PMPA injected through a guide cannula placed at 2.0 mm caudal and 1.6 mm lateral to the original LC cannula had no effect on the formalin-induced response. ZJ43 dose response in the LC. Dose-response curves for ZJ43 injection into the LC either contralateral or ipsilateral to the formalin injection site presenting the cumulative instances of formalin-induced flinching behavior during phase 1 and phase 2 of the formalin test (closed circle). Open circle represents sum of flinches by saline treated rats (control). ZJ43 or saline was microinjected into LC 10 min before the formalin injection. Each point represents the mean of five rats. ZJ43 reduced phase 1 and phase 2 response in a dose-dependent manner at the dose between 1.5 and 150 µg (by one-way ANOVA). * p < 0.05 by Tukey post-hoc test as compared with the phase 1 or phase 2 responses in the saline treated control rats (open circle).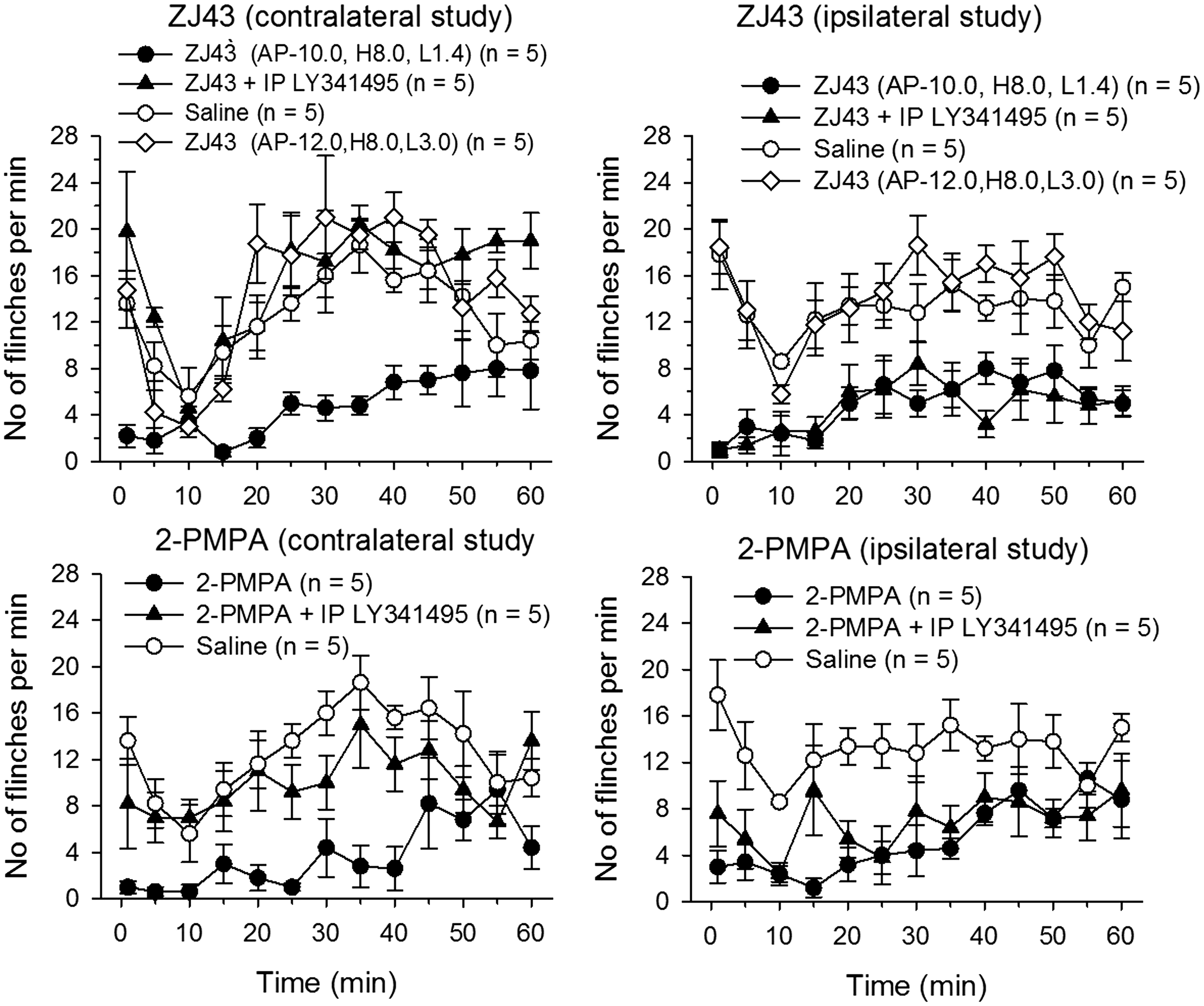

150 µg of 2-PMPA injected into LC either contralateral or ipsilateral to the formalin-injected side also significantly attenuated the both phase 1 and phase 2 flinching behaviors (phase 1: contralateral: p < 0.001; ipsilateral: p < 0.001), (phase 2: contralateral: p < 0.001; ipsilateral: p < 0.001) (Figure 8).
Effects of ZJ43 and 2-PMPA injected into contralateral LC, but not into ipsilateral LC, were completely antagonized by IP injection of the group II metabotropic glutamate receptor antagonist LY341495 (Contralateral: ZJ43: phase 1: p < 0.005, phase 2: p < 0.0005; 2-PMPA: phase 1: p < 0.01, phase 2: p < 0.05; Ipsilateral: ZJ43: phase 1: p > 0.3, phase 2: p > 0.8; 2-PMPA: phase 1: p > 0.05, phase 2: p > 0.4, by t-test) (Figure 8).
Spinal (intrathecal, IT) injection of the alpha 2 adrenergic receptor antagonist idazoxan reversed the analgesic effects of ZJ43 and 2-PMPA injected into contralateral LC, but not into ipsilateral LC (Figure 10; Contralateral: ZJ43 vs. ZJ + idazoxan: phase 1: p < 0.05, phase 2: p < 0.0005; 2-PMPA vs. 2-PMPA + idazoxan: phase 1: p < 0.05, phase 2: p < 0.005; Ipsilateral: ZJ43 vs. ZJ + idazoxan: phase 1: p > 0.7, phase 2: p > 0.6; 2-PMPA: phase 1: p > 0.4, phase 2: p > 0.05, by t-test). Idazoxan had no significant effect on ZJ43 or 2-PMPA injected into the ipsilateral LC.
Effect of intrathecal (IT, spinal) injection 30 µg of alpha 2-adrenergic receptor antagonist, idazoxan, on the analgesic effects of 150 µg of ZJ43 and 150 µg of 2-PMPA microinjected into either contralateral LC or ipsilateral LC to the formalin injected side. ZJ43 or 2-PMPA or saline were microinjected into LC 10 min before the formalin injection in the footpad. 30 µg of idazoxan was injected IT 10 min before the ZJ43 or 2-PMPA microinjection to verify the effect of ZJ43 or 2-PMPA was mediated by the activation of spinal alpha 2-adrenoreceptors. Analgesic effects of ZJ43 or 2-PMPA microinjected into LC contralateral, but not ipsilateral, to the formalin injection site were antagonized by IT idazoxan.
Discussion
Confirming an earlier study, 4 systemic treatment with a GCPII inhibitor reduces both phases of the response to inflammatory pain in the formalin footpad model. Consistent with the hypothesis that this effect is mediated by an increase in NAAG activation of mGluR3, this effect is blocked by the group II mGluR antagonist LY341495 (Figure 2). Previous microdialysis studies demonstrated that inhibitors of GCPII elevate extracellular NAAG levels3,5 and inhibit the release of neurotransmitters, including glutamate and GABA.3,5–8,22 NAAG and mGluR3 are widely distributed within the pain perception pathway and NAAG peptidase inhibitors introduced centrally into the spinal cord (intrathecal), third ventricle, contralateral periaqueductal gray, and rostral ventromedial medulla and peripherally into the footpad, reduced the first and second phase responses to formalin treatment.4,17–19,22
CNQX in the contralateral LC clearly discriminates between the roles of AMPA receptors as mediators of 2-PMPA effects on phase 1 and 2 responses to formalin treatment (Figure 5). The phase 1 response to systemic 2-PMPA seems unaffected by the antagonist in the LC while the phase 2 response is significantly reduced (p < 0.01). While the circuitry mediating the phase 1 response is not clearly resolved, the spinal cord is considered to have a central role. However, GCPII inhibition in other brain areas reduces both phases of the formalin response.19,22 More importantly, these data not only demonstrate that the contralateral LC contributes to the analgesic effects of systemic GCPII inhibition but that AMPA receptors in the LC have a role in this process.
Noradrenergic neurons in the LC mediate analgesia via axon terminals in the spinal dorsal horn. 33 Both the left and right LC have a significant influence on unilateral nociception in formalin test in rats, but the contralateral LC is reported to have a lesser effect. 33 In order to determine if this pathway is involved, the mechanism underlying the analgesic effects of systemic treatment with GCPII inhibitors,4,17,18 spinal noradrenaline levels were assessed in the inflammatory pain model. The finding that 2-PMPA increased flux of spinal noradrenaline following induction of inflammatory pain (Figure 4) supports the hypothesis that the LC contributed to the efficacy of the systemically administered peptidase inhibitor. Consistent with this conclusion, the analgesic effect of IP 2-PMPA was blocked by spinal treatment an alpha 2 adrenergic receptor antagonist, idazoxan (Figure 4). To test this directly, the two structurally different inhibitors were injected into the contralateral LC. Both drugs produced a group II mGluR-dependent inhibition of both phases of the response to inflammatory pain (Figures 7 and 8) via a process that was reversed by spinal application of idazoxan, a result that supports a role for alpha 2 adrenergic receptor mediation of the analgesic action of GCPII inhibition (Figure 10). A caveat to this interpretation of the idazoxan data is the ability of this drug to also act as imidazoline receptor agonist, an activity that could lead to non-competitive inhibition of NMDA receptors. 34 These data demonstrate that inhibitors of NAAG hydrolysis in the contralateral LC mediate analgesia via activation of noradrenergic LC neurons and support the hypothesis that systemic administration of these inhibitors achieves at least one element of their analgesic efficacy via this descending pathway.
The ipsilateral LC is in the primary ascending and the contralateral LC is in the primary descending inhibitory pain pathway. 33 Both innervate the dorsal spinal cord bilaterally (Figure 6) and GCPII inhibition in both LCs produced an analgesic-like effect in response to unilateral footpad inflammation (Figure 8). However, the mechanisms of action of the inhibitors differ between ipsilateral and contralateral LC. Inhibition in the LC contralateral to the formalin-treated footpad was reversed by systemic administration of the group II mGluR antagonist, a result that suggests NAAG activation of mGluR3. Consistent with the model of NAAG activation of presynaptic mGluR3 with consequent reduction in glutamate release,2,35 systemic administration of the inhibitor also reduced the inflammation-induced increase in glutamate levels in the contralateral LC. Similar inhibition of treatment-induced increases in transmitter release was obtained in animal models of stroke, traumatic brain injury and schizophrenia.3,5,6,22 However, given the multiple inputs to and transmitters acting within the LC, 33 it is difficult to model these results in the contralateral LC with decreased glutamate activity inasmuch as microinjection of the AMPA receptor antagonist in the contralateral LC reversed the effect of systemic 2-PMPA (Figure 5). This action of CNQX in the LC is similar to its efficacy in blocking the analgesic efficacy of gabapentin in the formalin inflammatory pain model. 26
While GCPII inhibition in the ipsilateral LC reduced the response to formalin treatment, this action was not sensitive to the group II antagonist (Figure 8), and thus was not mediated by NAAG activation of mGluR3. These data rule out a direct role for NAAG activation of a group II receptor in the ipsilateral LC in mediating analgesic action of systemically administered inhibitors since the systemic effect of the inhibitors is group II mGluR-dependent. Additionally, the ipsilateral LC differed from the contralateral in that formalin treatment did not elevate glutamate levels and these levels were not reduced by systemic treatment with 2-PMPA (Figure 3). Given the finding that NAAG also acts as an antagonist at some but not all subtypes of NMDA receptors,9,10,11,36 it might be that the reduced flinching response induced by ipsilateral microinjection of ZJ43 and 2-PMPA was mediated by NAAG acting as an antagonist at NMDA receptors in the LC containing 2 A and 2B subunits. However, the role of glutamate receptors in the LC is not clear. For example, in a study on the analgesic effects of LC treatment with estrodiol in the rat formalin model, 37 treatment with the NMDA antagonist (APV) and the AMPA antagonist (CNQX) in the ipsilateral LC did not significantly moderate the effect of estradiol on flinching behavior.
However, the failure of spinal administration of the alpha 2 receptor antagonist idazoxan to block the efficacy of ZJ43 and 2-PMPA that were microinjected into the ipsilateral LC (Figure 10) implies that this analgesic action is not the result of activation of noradrenergic neurons in the ipsilateral LC as would be suggested by NAAG antagonism of excitatory receptors in the LC. Inasmuch as the ipsilateral LC also provides an element in a secondary ascending pain projection pathway, 34 it may be that the efficacy of the GCPII inhibitors in the ipsilateral LC is due to NAAG antagonism of NMDA receptors 36 in this element of the ascending circuit.
Conclusion
Systemically administered GCPII inhibitors induce analgesia in this inflammatory pain model via a process that is blocked by a group II mGluR antagonist and by microinjection of an AMPA receptor antagonist directly into the contralateral LC. The mechanism underlying analgesia includes reduction of inflammation-induced elevation of glutamate release in the contralateral LC and increased release of noradrenaline at LC synapses in the spinal cord. Direct administration of GCPII inhibitors ZJ42 and 2-PMPA into the LC contralateral to the site of inflammation induced analgesia via a group II mGluR-dependent mechanism. Microinjection of the inhibitors into the ipsilateral LC also reduced the response to inflammation by a process that is not affected by a group II mGluR antagonist and thus is unrelated to group II mGluR activation by NAAG. These data provide additional proof of the concept that GCPII inhibition is a promising, novel treatment for inflammatory pain.
Footnotes
Authors’ contributions
Takahiro Nonaka: Design experimental protocol. Acquire and analyze the data; Toshihiko Yamada: Acquire and analyze the data. Tatsuhiro Ishimura: Acquire and analyze the data. Daiying Zuo: Acquire and analyze the data. John R Moffett: Acquired and analyzed data. Tatuso Yamamoto: Design experimental protocol and analyze the data. Co-drafted the manuscript. Joseph H Neale consulted on the experimental design and co-drafted the manuscript.
Declaration of Conflicting Interests
The author(s) have no conflicting interests associated with this research.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Tatsuo Yamamoto was supported by the Grant-in-Aid for Scientific Research (C). JHN supported by grants from the NIH and generous gifts from Nancy and Daniel Paduano.