
Abstract
Ethanol extracts of Cnidii Fructus, the dried fruits of Cnidium monnieri (L.) Cusson, have been externally applied in the treatment of Trichomonas vaginalis. However, the precise identity of the major constituents responsible for activity against T. vaginalis is unknown, but there is probability they are coumarin derivatives. In this study, the anti-Trichomonas activity of 4 major coumarin derivative constituents of Cnidii Fructus, namely, osthole, xanthotoxin, isopimpinellin, and bergapten, was characterized in terms of the resulting kinetics of growth and morphology of T. vaginalis upon treatment. The results demonstrated that osthole and xanthotoxol had significant trichomonacidal ability, while isopimpinellin and bergapten displayed low or no inhibitory efficacy toward T. vaginalis parasites. Our study suggests that the coumarin derivatives osthole and xanthotoxol can be potentially used as a basis for the development and design of new drugs for application in alternative or synergistic therapy against T. vaginalis.
Introduction
Trichomonas vaginalis (T. vaginalis) is a flagellate protozoan that causes the most prevalent non-viral or bacterial sexually transmitted disease, trichomoniasis. 1 This sexually transmitted disease (STD) increases the risks of infertility, 2 premature birth, 3 and enhance the risk of contracting cervical cancers 4 and other viral sexually transmitted diseases. 5 At present, trichomoniasis can be cured via oral administration of 5-nitroimidazole drugs, such as metronidazole (MTD). 6 However, patients’ partial noncompliance and intolerance to adverse reactions during metronidazole therapy result in recurrent and persistent infections. Persistent and recurrent infections, in turn, necessitate a higher metronidazole dosage for treatment; meanwhile, adverse reactions raise the risk of patient intolerance and, consequently, incomplete therapeutic courses and treatment failure. 6 In addition, a growing number of metronidazole-resistant vaginalis infections have been reported, leading to difficulty in treatment. 7 Therefore, there is an urgent need to exploit new reagents against T. vaginalis, preferably distinct chemical structures from 5-nitroimidazole derivatives, as an alternative or synergistic therapy with metronidazole reagents.8,9 Thus far, several anti-parasitic reagents have been developed from natural products and their derivatives.10-12 Moreover, many extracts or compounds isolated from natural resources all over the world have been investigated their susceptibility to T. vaginalis.8-10 In this study, we focused on the investigation of traditional Chinese plants to discover potential compounds against T. vaginalis parasites.
Among all recorded traditional Chinese plants, Cnidii Fructus (C. Fructus), the dried fruits of C. monnieri (L.) Cusson, has been widely applied in treating dermatological and vaginal itching or eczema.13,14 In addition, on the basis of the current Chinese pharmacopoeia, the ethanolic extract of Cnidii Fructus can be included in the preparation of lotion-based pharmaceutics and externally administered in clinics with other traditional Chinese medicine, such as Sophora flavescens Ait. and Prunus persica (L.) Batsch, to treat T. vaginalis.
15
However, even though its anti-Trichomonas activity has been demonstrated, the main bioactive constituent has not been clarified. According to numerous studies, coumarin derivatives are the main and most active constituents in C. Fructus ethanolic extracts.15-17 Among them, 4 compounds, namely, osthole, xanthotoxol (XTT), isopimpinellin (IPP), and bergapten (BGP), whose chemical structures are shown in Figure 1, are found at relatively high levels in C. Fructus.18-20 The contents of osthole and xanthotoxol (XTT) in C. Fructus are 1.5%–2.5% and 0.065%–0.11%, respectively, whereas C. Fructus contains 0.087%–0.15% isopimpinellin (IPP) and 0.05%–0.14% bergapten (BGP).
21
Herein, the anti-Trichomonas activities of these 4 main coumarin derivatives were independently investigated, and compared with those of metronidazole as a reference. The chemical structures of the coumarin derivatives in Cnidii Fructus: (A) osthole, (B) xanthotoxol, (C) isopimpinellin, and (D) bergapten.
Materials and Methods
Materials
Crude Cnidii Fructus drug was purchased from 10 local traditional Chinese medicine (TCM) markets in China and Taiwan and identified by Chien-Chih Yu. Metronidazole was obtained from the United State Pharmacopeial Convention, Inc. (North Bethesda, MD, USA). The osthole and bergapten (BGP) compounds were acquired from Yoneyama Yakuhin Kogyo Co. Ltd. (Osaka, Japan) and MilliporeSigma (St. Louis, MO, USA), respectively, whereas the xanthotoxol (XTT) and isopimpinellin (IPP) compounds were purchased from Extrasynthesis (Genay Cedex, France). The organic solvents, including ethanol and ethyl ether, were obtained from MerckMillipore (Burlington, MA, USA).
Preparation of the Cnidii Fructus Ethanolic Extract Paste
The C. Fructus extract paste was prepared following a previous report. 22 In brief, the C. Fructus samples (10 kg) were weighed and extracted with 100 L of ethanol following 2 rounds of heated reflux. The C. Fructus extracts were then filtered and combined. The combined alcoholic extract was collected and concentrated under reduced pressure using a rotary evaporator. The concentrated alcoholic extract was redissolved into hot petroleum ether and boiled. When the mixture cooled and delaminated, the upper layer was taken and concentrated into a C. Fructus paste form.
Kinetic Growth Curve of the Components of C. Fructus Against T. vaginalis
T. vaginalis, which had been pre-cultured for 40 h, was first incubated in 10 mL of medium at 37°C. Different concentrations of metronidazole, C. Fructus extract paste, and osthole, xanthotoxol (XTT), bergapten (BGP), and isopimpinellin (IPP) solutions were prepared in T. vaginalis growth medium, and 50 μL of these solutions were independently used in treatment of T. vaginalis. At predetermined time points (6, 12, 24, 30, 36, 48, 54, 60, and 72 h), 40 μL of the treated and control T. vaginalis suspensions was taken and blended with 100 mL of Isoton II buffering saline. The number of T. vaginalis was determined using a Coulter counter (Beckman Coulter Inc., Brea, CA, USA).
Toxic Assessment of the Components From C. Fructus Against Trichomonas vaginalis
T. vaginalis, incubated in 10 mL of medium at 37°C, was independently treated with 50 μL of metronidazole, C. Fructus extract paste, and the components of osthole, xanthotoxol, bergapten, and isopimpinellin. The concentrations of each compound and extract paste were previously adjusted to 0.25 mg/mL with the growth medium. After 48 h of incubation, a drop of T. vaginalis suspension in the medium was deposited onto a glass slide and the morphology of the T. vaginalis with or without treatment was optically observed using an inverted biological microscope. Simultaneously, 1 mL of T. vaginalis in the medium with or without treatment was collected and analyzed via flow cytometry (Beckman Coulter Inc., Brea, CA, USA).
Results
Kinetic Growth Curve of Trichomonas vaginalis With or Without Treatment
C. Fructus ethanolic extracts were applied in the treatment of trichomoniasis to characterize the effects on T. vaginalis. Herein, first, we confirmed the anti-T. vaginalis activity of the C. Fructus ethanolic extract paste by determining the kinetic growth curves of the T. vaginalis parasites with or without treatment. The kinetic growth curves were acquired by incubating the T. vaginalis parasites in the medium with or without C. Fructus extracts, where the number of parasites was monitored for 72 h using a Coulter counter, as shown in Figure 2. In Figure 2, the kinetic growth curves of T. vaginalis without any treatment, representing the control group, can be distinguished into 4 phases of progression: the lag phase, the logarithmic phase, the maximum stationary phase, and the death (decline) phase.
23
These phases occur after 0-20 h, 20-48 h, 48-60 h, and >60 h of incubation, respectively. Within the first 48 h incubation (in the lag and logarithmic phase), T. vaginalis displayed initiation of multiplication and the populations rapidly grew; after 48 h, T. vaginalis parasites reached maximum growth with populations (the maximum stationary phase) and the growth rate decreased, while at 60 h post-incubation, the populations of the T. vaginalis parasites declined, indicating that the growth curves were entry into the death (decline) phase. However, when the T. vaginalis parasites were treated with C. Fructus extract paste, the dose-dependent manner inhibition of growth or trichomonacidal effect was observed. When 25 mg/mL of the extract paste was applied, the growth of the T. vaginalis parasites was retarded, showing an inhibitory effect; as the extract concentration changed to 2.5 or 12.5 mg/mL, the kinetic growth curve revealed a trichomonacidal effect. The results in Figure 2 clearly indicate that C. Fructus ethanolic extract paste contains some substances against T. vaginalis parasites. Therefore, the main components of the ethanolic extracts were primarily investigated for their anti-T. vaginalis ability. The kinetic growth curves of the Trichomonas vaginalis parasites without or with 2.5, 12.5, and 25 mg/mL of C. Fructus extract.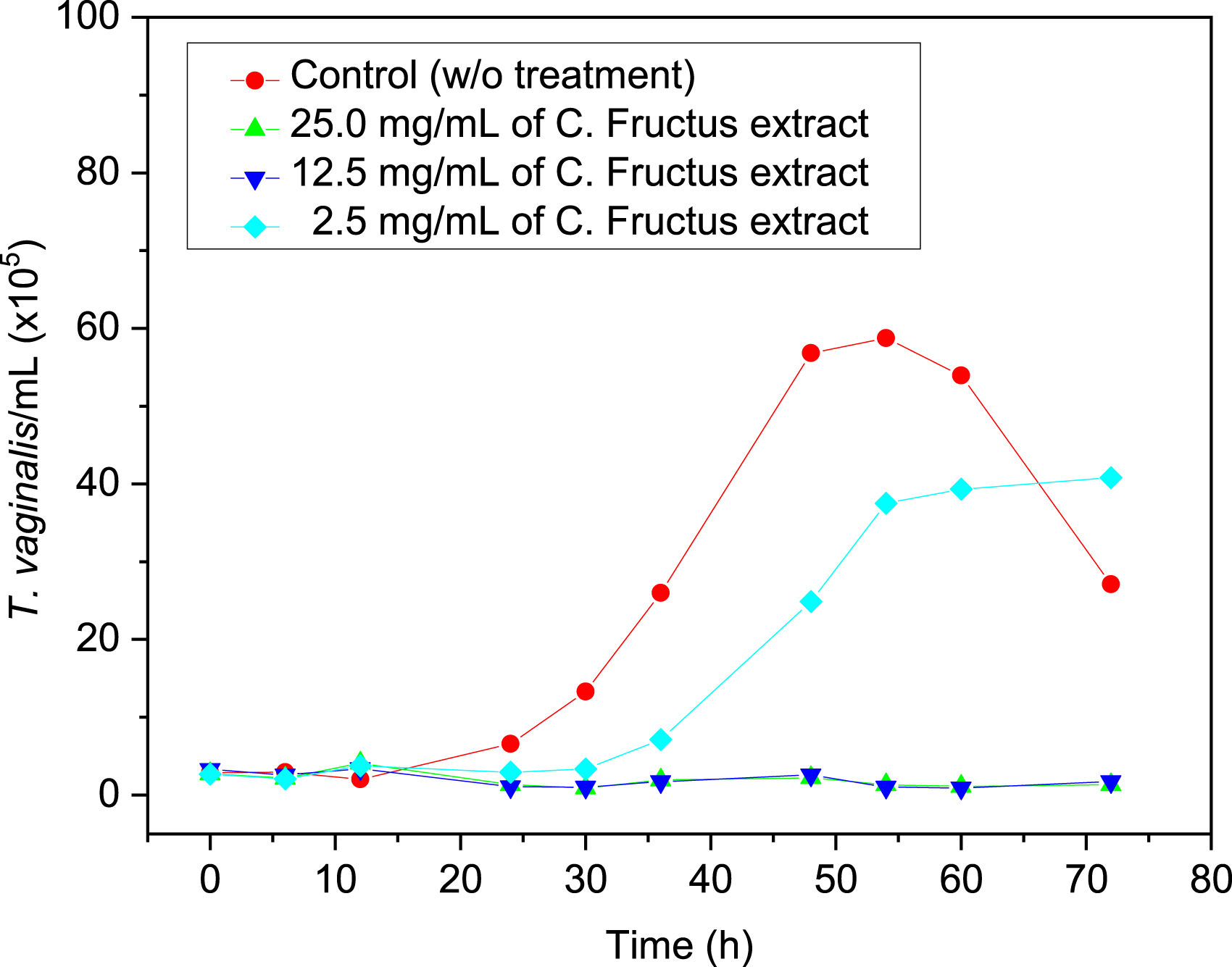
In order to further clarify the main constituents in C. Fructus against T. vaginalis parasites, the kinetic growth curves were determined for the T. vaginalis parasites treated with 4 main bioactive components, osthole, xanthotoxol, isopimpinellin, and bergapten, whereas their concentrations were adjusted according to their respective contents in C. Fructus. Meanwhile, as a comparison, T. vaginalis parasites were also treated with metronidazole, which is a 5-nitroimidazole derivative and is widely applied against T. vaginalis in clinics. As Figure 3A shows, the data for the number of T. vaginalis parasites treated with metronidazole reveal a trichomonacidal effect that is enhanced with increasing treatment concentration. When the treatment concentration reached to 5x10-3 mg/mL, an obvious trichomonacidal effect was observed in the growth curves. However, when the T. vaginalis parasites were treated with the components derived from C. Fructus, the kinetic growth curves were distinct. As Figure 3B displays, when .05 mg/mL of osthole was applied to the T. vaginalis parasites, a retardation in growth was revealed, representing an inhibitory effect on the parasites. However, when the treatment concentration of osthole increased to 0.5 mg/mL, that was approximately equal to the concentration of the osthole in 25 mg/mL of C. Fructus ethanolic extract, complete prevention of proliferation occurred, indicating trichomonacidal ability. These kinetic growth curves are similar to those of parasites treated with another constituent, that is, xanthotoxol. As Figure 3C shows, a trichomonacidal effect was discovered in those parasites treated with 0.05 mg/mL of xanthotoxol, whereas the inhibitory effects were witnessed in parasites when 0.25 mg/mL of xanthotoxol was given. Though, these concentrations were respectively 2 and 10-fold higher than the levels in C. Fructus ethanolic extract. In contrast to these results, when the T. vaginalis parasites were treated with bergapten, the kinetic growth curves did not alter, as shown in Figure 3D. Furthermore, as Figure 3E shows, when isopimpinellin, the other main component in the C. Fructus extract paste, was applied to the T. vaginalis parasites at a dosage higher than 0.05 mg/mL, proliferation of the parasites was subtly inhibited, while the growth trend was approximate to that without any treatment. The results clearly demonstrate the low inhibitory effects of bergapten and isopimpinellin against T. vaginalis. In addition, the results demonstrated that the osthoel was the main active constituent of C. Fructus ethanolic extracts against T. vaginalis parasites, while xanthotoxol, whose concentration was low in C. Fructus ethanolic extracts was one of the candidates for T. vaginalis parasites inhibition. The kinetic growth curves of the Trichomonas vaginalis parasites, independently treated with (A) metronidazole (MTD), (B) osthole, (C) xanthotoxol (XTT), (D) bergapten (BGP), and (E) isopimpinellin (IPP).
In Vitro Toxicity Assessment of the Main Constituents in C. Fructus Toward T. vaginalis
The in vitro trichomonacidal activities of the investigated compounds and the C. Fructus ethanolic extract were further estimated using flow cytometry. In brief, the T. vaginalis parasites were cultured in the medium or treated with the C. Fructus extract paste and the main components for 48 h. Afterward, the morphology of the T. vaginalis parasites was determined using an optical microscopy and flow cytometry and compared to that of parasites without any treatment (as a control group), and the optical microscopic images are presented in Figure 4. In Figure 4A, the parasites without any treatment exhibited complete morphologies, and they could be detected using forward scattering (FSC) and side scattering (SSC) in flow cytometry. However, when the T. vaginalis parasites were treated with C. Fructus extract paste for 48 h, deformation and destruction were observed, as shown in Figure 4B. Their deformation led to significant movement of the population in the flow cytometric analysis. The results correspond to the kinetic growth curves after C. Fructus extract treatment and again confirm the trichomonacidal activity of the C. Fructus extract paste. Furthermore, the anti-Trichomonas activity of the other components and metronidazole were further examined. The morphology of the Trichomonas vaginalis parasites. The parasites (A) without any treatment or (B) treated with 12.5 mg/mL of the C. Fructus ethanolic extract paste were incubated for 48 h and the morphology was observed via optical microscopy (200×) and detected with flow cytometry.
In order to precisely investigate the anti-Trichomonas activity of these components containing C. Fructus ethanolic extracts, their concentrations and metronidazole (MTD) reagents were adjusted to 0.25 mg/mL for comparison. After incubation with T. vaginalis parasites for 48 h, as Figure 5A shows, the metronidazole treatment led to the death of a huge number of the T. vaginalis parasites with disrupted morphology, leading to a shift in the parasite population on the basis of the flow cytometric analysis. Moreover, treatment with 0.25 mg/mL of osthole and xanthotoxol (XTT), 2 of the main components in Cnidii Fructus, led to similar results. As Figure 5B and 5C indicates, respectively, the same concentrations of osthole and xanthotoxol (XTT) resulted in the deformation and morphological destruction to the incubated T. vaginalis parasites; hence, flow cytometry was also used to detect the movement of these populations. However, on the basis of the flow cytometry analysis in Figure 5D and 5E, for the other 2 components, namely, bergapten (BGP) and isopimpinellin (IPP), there was not change in parasite populations after treatment. After treatment with bergapten for 24 h, the T. vaginalis parasites displayed intact morphology, in comparison of those without any treatment (Figure 5A), while subtle deformation after isopimpinellin (IPP) treatment was observed in the T vaginalis parasites. These results correspond to their kinetic growth curves and further confirm that osthole and xanthotoxol have trichomonacidal effects, whereas isopimpinellin exhibited inhibitory effects on the T. vaginalis parasites. The morphology of the Trichomonas vaginalis parasites following treatment with the compounds. The T vaginalis parasites were independently treated with 0.25 mg/mL of (A) metronidazole (MTD), (B) osthole, (C) xanthotoxol (XTT), (D) bergapten (BGP), and (E) isopimpinellin (IPP). At 48 h post-incubation, the morphology of the T. vaginalis parasites was examined using an optical microscopy (200 ×) and flow cytometry.
Discussion
In recent investigations, diverse plant compounds with chemical structures differing from that of 5-nitroimidazole were isolated, and the susceptibility of T. vaginalis parasites to these compounds was tested.8-10 To date, several chemical backbones and their anti-Trichomonas activity have been identified.8-10 For example, Potadar et al demonstrated that the β-glycoside constituent berberine, extracted from Berberis aristate, has an inhibitory effect on the growth of T. vaginalis parasites. 24 Tiwari et al illustrated that 0.05 mg/mL of saponins achieves trichomonacidal ability in 12 h, 25 whereas alkaloid derivatives such as candimine and lycorine, extracted from Hippeastrum morelianum and the members of the Amaryllidaceae family, respectively, also possess anti-Trichomonas activity.26,27 Furthermore, essential oils from herbs have been reported to have superior anti-Trichomonas activity.8-10 For instance, Dai et al indicated that Lanxangia tsao-ko (Amomum tsiao-ko) shows anti-T. vaginalis activity. They further identified that the anti-Trichomonas activity is associated with the major oil component geraniol, while the efficacy is still weaker than that of metronidazole. 28 Chiekh-Ali et al showed that in comparison to metronidazole, Aframomum sceptrum essential oil, which includes β-pinene and caryophyllene oxide as major active components, exhibits better trichomonacidal efficacy. 29 These reports clearly highlight the feasibility of discovering a lead compound in anti-T. vaginalis parasites from natural products.8-10 In fact, partial lead compounds are currently being developed via chemical modification to reinforce their anti-Trichomonas activity.30,31
In this study, we identified the anti-Trichomonas efficacy of C. Fructus ethanolic extracts from their kinetic growth curves and morphology, as shown in Figure 2B and 4B. However, on the basis of the kinetic growth curves of the parasites, the efficacy of C. Fructus displayed weaker anti-Trichomonas activity, in comparison to that of metronidazole (Figures 5A and 3A). This could be explained on the basis that not all of the major components in the C. Fructus ethanolic extracts have anti-Trichomonas activity, as we observed upon testing of individual components. On the basis of our verification, as shown in Figures 3 and 5, among the 4 coumarin derivatives in the C. Fructus ethanolic extracts, only osthole and xanthotoxol enabled the efficient inhibition of T. vaginalis growth, whereas the other 2 components, bergapten and isopimpinellin, displayed no or relatively low anti-Trichomonas activity, respectively. Moreover, on the basis of multiple studies, the active osthole content was approximately 1.5%–2.5%, whereas the content of active xanthotoxin was only 0.065%–0.11% in C. Fructus. 21 Therefore, as the concentration of the active ingredients osthole and xanthotoxol was adjusted to the same as metronidazole, the anti-T. vaginalis activity could be clearly observed, as shown in Figures 3B, C and 5B, C). These results suggest that osthole and xanthotoxol are potential new drug candidates for an alternative or synergistic treatment against T vaginalis. To the best of our knowledge, this is the first report to identify the major active components of C. Fructus ethanolic extracts against T. vaginalis parasites. Furthermore, our study is the first to report the anti-T. vaginalis activity of coumarin derivatives, suggesting that this chemical structure is worthy of application as a lead compound for further chemical modifications or in considering new drug designs. However, the pharmacological mechanism of the 2 coumarin derivative compounds against T. vaginalis is unclear and necessitates further investigation to support their use in future applications.
Conclusions
In this study, we confirmed the anti-T. vaginalis activity of Cnidii Fructus ethanolic extracts. We further identified that the coumarin derivatives osthole and xanthotoxin are the 2 major constituents that are active against Trichomonas parasites. Thus, this study affords 2 new candidates for the development of new drugs in the treatment of Trichomoniasis.
Footnotes
Acknowledgments
The authors sincerely appreciated China Medical University for the financial supports.
Author Contributions
C.-C. Y. identified the crude traditional Chinese herbs and prepared the C. Fructus ethanolic extracts. The experiments were conception by C.-C. Y. and the details were designed by T.-M. C. The experiments were performed by the research group of T.-M. C. The manuscript was written by Y.-T. C. under the supervision of C.-C. Y.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the China Medical University (CMU109-MF-54 and CMU110-MF-44).