
Abstract
Introduction
Ionizing radiation has been widely applied to brain disease diagnosis and radiotherapy to treat brain tumor and to prophylactically prevent metastasis of peripheral cancer cells to the brain. However, acute or fractional accumulative ionizing radiation may result in neurodegenerative disorders including cognitive impairment, depression, sensory disturbance, and poor motor coordination.1,2 Previous studies have shown that irradiation could lead to cognitive impairment and depression by reducing neurogenesis in hippocampus after the acute radiation exposure.3-6 Although many neuroprotective agents have been reported to have significant efficacy on radiation-induced cognitive dysfunction, amifostine is still the only approved radioprotectant during radiotherapy. 7 Our previous study has indicated that the oral administration of Chinese herbal medicine (CHM) epimedium after acute radiation exposure could significantly improve the hippocampal neurogenesis and radiation-induced depression and cognitive impairment in BALB/c mice. 5 Flavonoids, the active components of epimedium and many other CHMs, can reduce oxidative stress significantly. 8 This implicated that CHMs containing flavonoids may have great advantages as radio-neuro-protectants because of their anti-oxidative and immunoregulatory effects.
Lycium barbarum berry, also named Goji berry and wolfberry, which is rich of Lycium barbarum polysaccharides (LBPs), has been used as food supplement and herbal medicine in China for thousands of years and has been increasingly accepted as anti-oxidative and anti-aging fruit in Western countries.9,10 Its components, including flavonoids and many glycosylated derivatives of dicaffeoylspermidine, dicaffeoylspermine, and kukoamine, have been studied intensively.11,12 The mixture of those glycosylated derivatives in Lycium barbarum berry, the LBPs, can improve retinal function and reduce retinal neuron damage,13,14 reduce blood lipid and glucose,15,16 boost immune system to anti-allergy and anti-cancer, 9 protect against radiotherapy or chemotherapy induced tissue damages, 17 prevent neuron loss, and enhance neurogenesis. 18 Flavonoids can pass through the blood-brain barrier 19 to play their roles in anti-oxidation, anti-inflammation, and neuro-protection. 20 The dicaffeoylspermidine derivatives showed significant anti-oxidative and neuroprotective effects. 21 The radio-neuro-protective effects of kukoamines may have the similar anti-oxidative and anti-inflammatory pathways. 22 In addition, kukoamine A may down regulate the NMDA receptors to prevent the NMDA-induced neurotoxicity in vitro. 23 As a nutrient rich “superfood,” Lycium barbarum berry can be used as a daily diet without serious accumulative toxins or side-effect.10,24 Its potential neuroprotective effects after radiation exposure should be evaluated. In the present study, we studied the radio-neuro-protective effects of Lycium barbarum berry extract on neurobehavioral changes and hippocampal neuron loss in the mouse model of acute ionizing radiation exposure.
Materials and Methods
Lycium barbarum Berry Extraction
Five hundred grams dried Lycium barbarum berries (Zhongning, Ningxia, China) were crushed and decocted in 3 L boiled distilled water for 2 h. The decocted liquid was filtrated and the residue was decocted again in 2 L water. The 2 filtrates were combined and concentrated in rotary evaporator under 60°C to obtain 350 g thick paste which was stored under 4°C. The paste containing the water-soluble components of Lycium barbarum berry was dissolved in saline at the highest concentration of .5 g/mL (10 g/kg) before oral administration with the volume of .2 mL per 10 g body weight of mouse. Based on their efficacies on preventing animal body weight loss and behavioral changes after administration of 3 different dosages including 2.5, 5,, and 10 g/kg, the highest dosage of 10 g/kg of Lycium barbarum berry water extraction was chosen for further study.
Experimental Animal
Male specific-pathogen-free (SPF) BALB/c mice were purchased from Hubei experimental animal research center licensed with the number SCXK (Hubei) 2015-0018, and accommodated in local animal room to 8-week-old and body weight 22 ± 2 g before irradiation. Animals were divided into four groups: (1) sham X-ray exposure group (Sham) of 10 mice with oral saline administration after sham exposure; (2) experimental control group (Exp-Ctrl) of 10 mice with 5.5 Gy X-ray whole body irradiation followed by oral saline administration; (3) positive experimental control group (Ami-Exp) of 10 mice with the amifostine pretreatment (at 100 mg/kg, batch number: J0311A, Dalian Meilun Biotech. Co., China) at 30 min before 5.5 Gy X-ray whole body irradiation; and (4) experimental group (Exp-Lyc) of 10 mice with 5.5 Gy X-ray whole body irradiation followed by the oral administration of the Lycium barbarum berry extract (10 g/kg per day) 2 h after radiation exposure on the first day, and the Lycium barbarum berry extract treatment was continued for 4 weeks. The body weight was recorded after the irradiation. Behavioral tests were conducted four weeks after radiation exposure and the mice were then sacrificed for the brain sample collection.
Behavioral Tests
To test if irradiation induced depression and spatial memory impairment and if Lyc could improve those changes, tail suspension, forced swimming, open field, and Morris water maze tests were used as scheduled in Figure 1A. Behavioral tests were conducted in a quiet room with dim light and constant temperature (20-25°C) using SuperMaze animal behavior record and analysis system (Shanghai Xinruan InfoTech Co, China). The animals have been moved into the behavioral test room after the last oral administration. One day after, tail suspension and forced swimming immobility times were recorded for 4 min after 2 min environmental adaptation. The open field size of 50 cm (length) × 50 cm (width) × 30 cm (height) was divided into 25 boxes to record the total movement distance and the time spent in the central area (central 9 boxes) for 10 min. The animal was placed on the central point of open field to begin the test. The Morris water maze experiments were done as described previously.
5
The hidden platform escape latency was recorded for 5 days. On the day 6, after removing the platform, the times that the mice passed though platform area and the total resident time that the mice spent in the platform quadrant in 60 s were recorded. The experimental time schedule (A) and the effect of Lycium barbarum berry extract on the body weight of radiation exposed BALB/c mice (B). The body weight of the mice in the sham exposure group (Sham) increases steadily, but it decreases significantly after acute X-ray irradiation with 5.5 Gy in the experimental control (Exp-Ctrl), mice treated with Lycium barbarum berries (Exp-Lyc) and amifostine (Ami-Exp) at 1 week after the exposure (F (3, 36) = 61.85, P < .0001). From the second week, amifostine pretreatment (Ami- Exp) and oral Lycium barbarum berry extract treatment after exposure (Exp-Lyc) reverse the animal weight loss (F (3, 36) = 134.0, P < .0001) and these animals have similar body weight to Sham group at the end of the fifth week (F (3, 36) = 244.0, P < .0001). However, Exp-Ctrl group has a significant weight loss from 2 to 5 week(s) after radiation exposure when compared to the other 3 groups (P < .05, n = 10 in each group).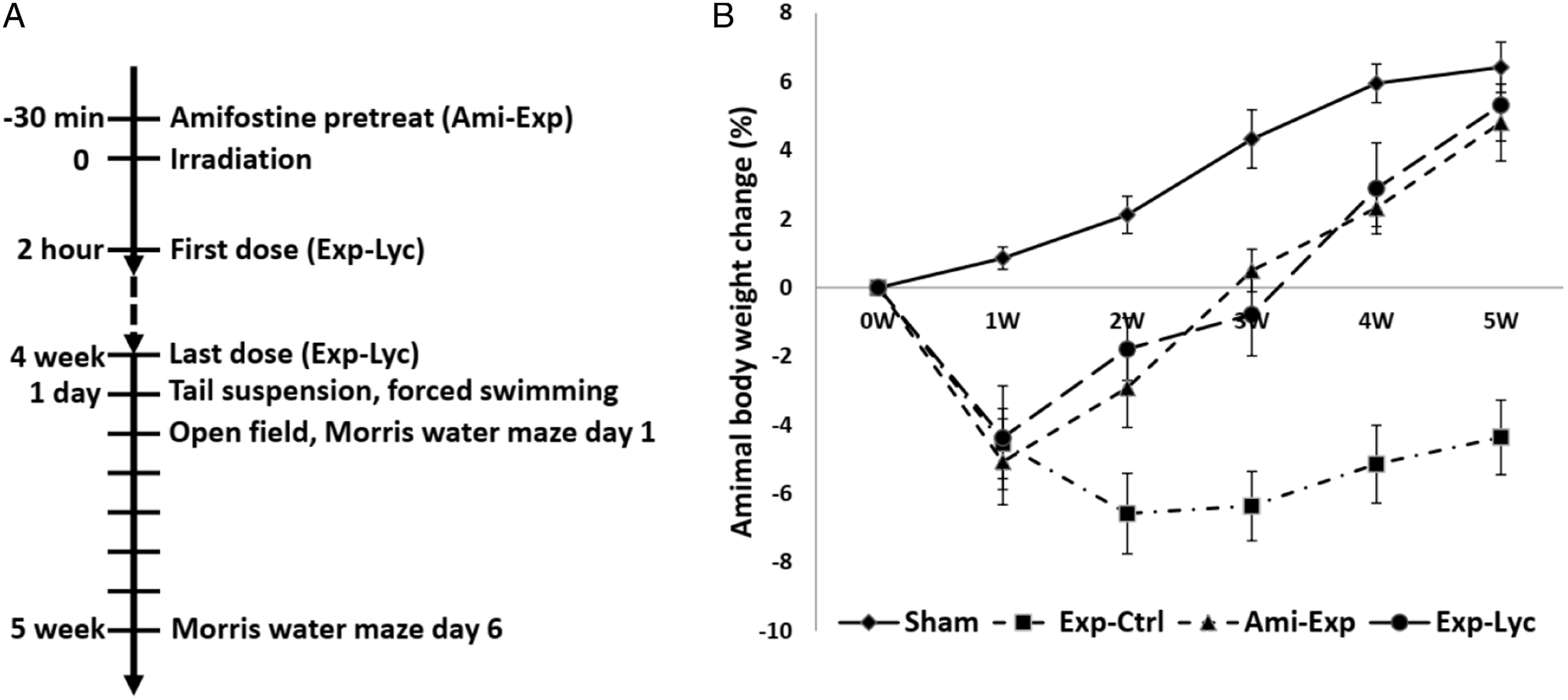
Immunohistochemistry
The mice were anesthetized with 1% pentobarbital after behavioral tests. Animals were perfused transcardially with .9% saline to wash out the blood followed by 4% paraformaldehyde. The brain was removed, postfixed with the same fixative overnight, and then stored in 30% sucrose solution prepared with .1 M phosphate buffer (pH: 7.4). For immunohistochemistry, sagittal brain sections with 40 μm thickness were cut and a set of 3 serial sections was prepared and sections collected were placed individually in different wells of a 24 well tissue culture dish for the control, NeuN, calbindin (CB), and parvalbumin (PV) immunohistochemical reaction. The freely floating sections were treated in 4% normal goat serum for 2 h at room temperature. All sections were then washed in .1 M phosphate-buffered saline (PBS) containing NeuN (1:100) (Invitrogen, United States), CB (1: 2000), and PV (1:1000) (Swant, Switzerland). After incubation, sections were washed in PBS and placed for 1 h in biotinylated goat anti-rabbit IgG (Vector Laboratories, Burlingame, CA, United States) diluted 1:2000 in PBS/Triton X-100. After 2 washes in PBS, they were placed in avidin– biotin complex (ABC) reagent (Vector Laboratories) in PBS/Triton X-100 for 1 h, washed in PBS and reacted in a solution of .12% H2O2 and .05% 3,3-diaminobenzidine (DAB) (Sigma, United States) in Tris buffer (TB) for 15 min, then mounted, dehydrated, coverslipped, and photographed by using image analysis system.
Cell Counting and Statistical Analysis
NeuN immunopositive neurons in the hilus of the dentate gyrus, CB immunopositive interneurons in the area of CA1 strata radiatum lacunosum moleculare and stratum oriens, PV immunopositive interneurons in the CA1 stratum pyramidum, and the stratum granulosum of the dentate gyrus in the dorsal hippocampus were counted by using Leica DM4 B upright microscope and LAS X software (Leica Microsystems, Germany). The 5 sagittal brain sections from every 3 alternative sections of the dorsal hippocampus of each animal were used for immunopositive cell counting. Five animals of each group were used for cell counting. All the data sets have passed the normal distribution tests by using Prism 8. One-way ANOVA followed by the Bonferroni post-hoc test for multiple comparisons was used for statistical analysis and statistical significance was considered when the adjusted P < .05. All the data were showed as mean ± SD.
Results
Body Weight
The animal body weight changes were recorded for 5 weeks since the day of irradiation (Figure 1B). X-ray irradiation with 5.5 Gy (Exp-Ctrl) induced a significant body weight loss in the first week after exposure compared to the sham radiation exposure mice (Sham) (F (3, 36) = 61.85, P < .0001). Oral administration of Lycium barbarum berry extract at the dosage of 10 g/kg per day (350 g extract paste from 500 g dried Lycium barbarum berries) (Exp-Lyc) prevented animal weight loss from the second week after irradiation, similar to amifostine pretreatment (Ami-Exp) with intraperitoneal injection at 100 mg/kg 30 min before irradiation (F (3, 36) = 134.0, P < .0001). Both therapeutic strategies in Exp-Lyc and Ami-Exp groups effectively improved the animal weight gain to the normal level in the Sham group by the end of the fifth week after irradiation. At this time point, animals in Exp-Lyc and Ami-Exp groups were significantly heavier than those in Exp-Ctrl (F (3, 36) = 244.0, P < .0001), but had no significant weight difference compared to the Sham group (P > .05).
Behavioral Tests
The average tail suspension (Figure 2A, F (3, 36) = 8.267, P = .0003) and forced swimming (Figure 2B, F (3, 36) = 12.27, P < .0001) immobility time in 4 min of those radiation exposed mice with saline treatment were significantly longer than the sham exposed mice. Amifostine or Lycium barbarum berry treatment significantly reduced immobility time when compared to irradiated animals treated with saline (Figures 2A, B, P < .05). No significant difference in immobility time was observed among those exposed animals treated with amifostine or Lycium barbarum berry and the Sham group (P > .05). Open field experiment showed that the total travel distance in the Exp-Ctrl group within 10 min was less than the Sham group, amifostine, or Lycium barbarum treated mice, indicating both treatments significantly improved animal locomotor activity (Figure 2C, F (3, 36) = 10.85, P < .0001). Similarly, Lycium barbarum berry also reduced the time animals stayed in the central area of test field when compared to radiation exposed control mice (Figure 2D, F (3, 36) = 13.14, P < .0001). Morris water maze experiments showed that the average platform escape latency time of radiation exposure mice (Exp-Ctrl) was significantly longer than the Sham, Ami-Exp, or Exp-Lyc group on the day 5 (Figures 3A, B, F (3, 36) = 35.53, P < .0001). In addition, the average platform crossing time and platform quadrant resident time of the Exp-Ctrl group on day 6 were less than the Sham, Ami-Exp, or Exp-Lyc group (Figure 3C, F (3, 36) = 29.24, P < .0001). Amifostine or Lycium barbarum berry treatment reduced the escape latency and increased the platform crossing time and platform quadrant resident time significantly, indicating that both treatments could improve the spatial memory of mice with radiation exposure. These data of behavioral tests demonstrated that the oral administration of Lycium barbarum berry extract could effectively prevent or treat the radiation damages in the motor activity and cognitive function of mice. Tail suspension, forced swimming, and open field tests. Similar to the amifostine pretreated (Ami-Exp) group, Lycium barbarum berry extract (Exp-Lyc) significantly reduces the immobility times in tail suspension (A, F (3, 36) = 8.267, P = .0003) and forced swimming (B, F (3, 36) = 12.27, P < .0001) tests. Open field test shows the average total travelling distance of the mice in radiation exposure control group (Exp-Ctrl) is significantly less than those of the mice in the other 3 groups (C, F (3, 36) = 10.85, P < .0001). However, the average central area staying time of Exp-Ctrl mice is significantly longer (D, F (3, 36) = 13.14, P < .0001). Asterisks * indicate P < .05. The values are the mean ± SD calculated from the data of 10 mice in each group. Morris water maze test. Morris water maze test shows that the average escape latency of the Exp-Ctrl group is significantly longer than those of the Sham group, the Ami-Exp or Exp-Lyc group in the training trials on the day 5 (A, B, F (3, 36) = 35.53, P < .0001). The average time spent in the platform quadrant and the times of animal crossing platform area of the Exp-Ctrl group are significantly lower than those of other 3 groups on day 6 (C, F (3, 36) = 29.24, P < .0001). Asterisks * in panel C indicate P < .05 when compared with the Exp-Ctrl group. The values are the mean ± SD calculated from the data of 10 mice in each group.

Immunohistochemistry
NeuN, CB, and PV immunopositive neurons in the target areas of dorsal hippocampus were counted from 5 slices of each mouse brain. The number of NeuN immunopositive neurons in the hilus of the dentate gyrus was significantly reduced after irradiation with 5.5 Gy (Figure 4, F (3, 36) = 26.04, P < .0001, Exp-Ctrl compared to Sham, P < .05). Both amifostine pretreatment and Lycium barbarum berry extract treatment prevented hilar neuronal loss when compared to the Exp-Ctrl (Figure 4, Ami-Exp or Exp-Lyc compared to Exp-Ctrl, P < .05). Similarly, the number of CB positive neurons in the strata radiatum lacunosum moleculare (Figure 5E, F (3, 36) = 16.88, P < .0001) and stratum oriens (Figure 5F, F (3, 36) = 16.47, P < .0001), and the number of PV positive interneurons in the CA1 stratum pyramidum (Figure 6M, F (3, 36) = 27.75, P < .0001) and the stratum granulosum of the dentate gyrus (Figure 6N, F (3, 36) = 6.78, P = .0037) were also reduced by radiation exposure, indicating the radiation induced neuron loss in these areas. Oral administration of Lycium barbarum berry effectively increased the numbers of CB and PV immunopositive cells in the corresponding areas (Figures 5, 6, Exp-Lyc compared to Exp-Ctrl, P < .05). These immunochemistry results demonstrated that the Lycium barbarum berry could effectively prevent the loss of NeuN, CB, and PV immunopositive cells in the hilus and stratum granulosum of the dentate gyrus, strata radiatum lacunosum moleculare, stratum oriens, and pyramidum of CA1 area of the mouse hippocampus. NeuN immunohistochemistry. A, B, C, and D show the NeuN immunopositive neurons in the hilus of the dentate gyrus (the triangle area indicated in each slice) of the representative slices from the Sham, Exp-Ctrl, Ami-Exp, and Exp-Lyc groups, respectively. E, Statistical analysis shows the number of NeuN immunopositive neurons in the hilus of the Exp-Ctrl group (pyramidal neurons in CA3c were excluded) is significantly fewer than the Sham group (F (3, 36) = 26.04, P < .0001). The Ami-Exp or Exp-Lyc group has more NeuN immunopositive neurons in the hilus than the Exp-Ctrl group. Asterisks * indicate P < .05 when compared with the Exp-Ctrl group (n = 5 in each group, cell number is counted in 5 sagittal brain sections from every 3 alternative sections of the dorsal hippocampus of each mouse, and indicated as the mean ± SD). Calbindin (CB) immunohistochemistry. A, B, C, and D show the CB immunopositive interneurons in the strata radiatum, lacunosum moleculare (SRLM) and oriens (SO) in CA1 areas (the areas indicated in the slice) of the slices from the Sham, Exp-Ctrl, Ami-Exp, and Exp-Lyc groups, respectively. E and F show the numbers of CB immunopositive interneurons in the CA1-SRLM (F (3, 36) = 16.88, P < .0001) and CA1-SO (Panel F, F (3, 36) = 16.47, P < .0001) areas of the Exp-Ctrl group are statistical significantly fewer than the Sham group. Both Ami-Exp and Exp-Lyc groups have more CB immunopositive interneurons in these areas than the Exp-Ctrl group. Asterisks * indicate P < .05 when compared with the Exp-Ctrl group (n=5 in each group, cell number is counted in 5 sagittal brain sections from every 3 alternative sections of the dorsal hippocampus of each mouse, and indicated as the mean ± SD). Parvalbumin (PV) immunohistochemistry. The areas indicated in the slices A, D, G, and J show the PV immunostaining interneurons in the CA1 stratum pyramidum (CA1-SP, B, E, H, and K) and the stratum granulosum of the dentate gyrus (DG-SG, C, F, I, and L) from the Sham, Exp-Ctrl, Ami-Exp, and Exp-Lyc groups, respectively. The picture D shows significant reduced numbers of PV immunopositive interneurons in the CA1-SP (E and M, F (3, 36) = 27.75, P < .0001) and the DG-SG (F and N, F (3, 36) = 6.78, P = .0037) of the Exp-Ctrl group when compared to the Sham group. The numbers of PV immunopositive interneurons in the CA1-SP (M) and the DG-SG (N) of the Ami-Exp (G, H, I) or Exp-Lyc group (J, K, and L) are significantly more than the Exp-Ctrl group. Asterisks * indicate P < .05 when compared with Exp-Ctrl group (n = 5 in each group, cell number is counted in 5 sagittal brain sections from every 3 alternative sections of the dorsal hippocampus of each mouse, and indicated as the mean ± SD).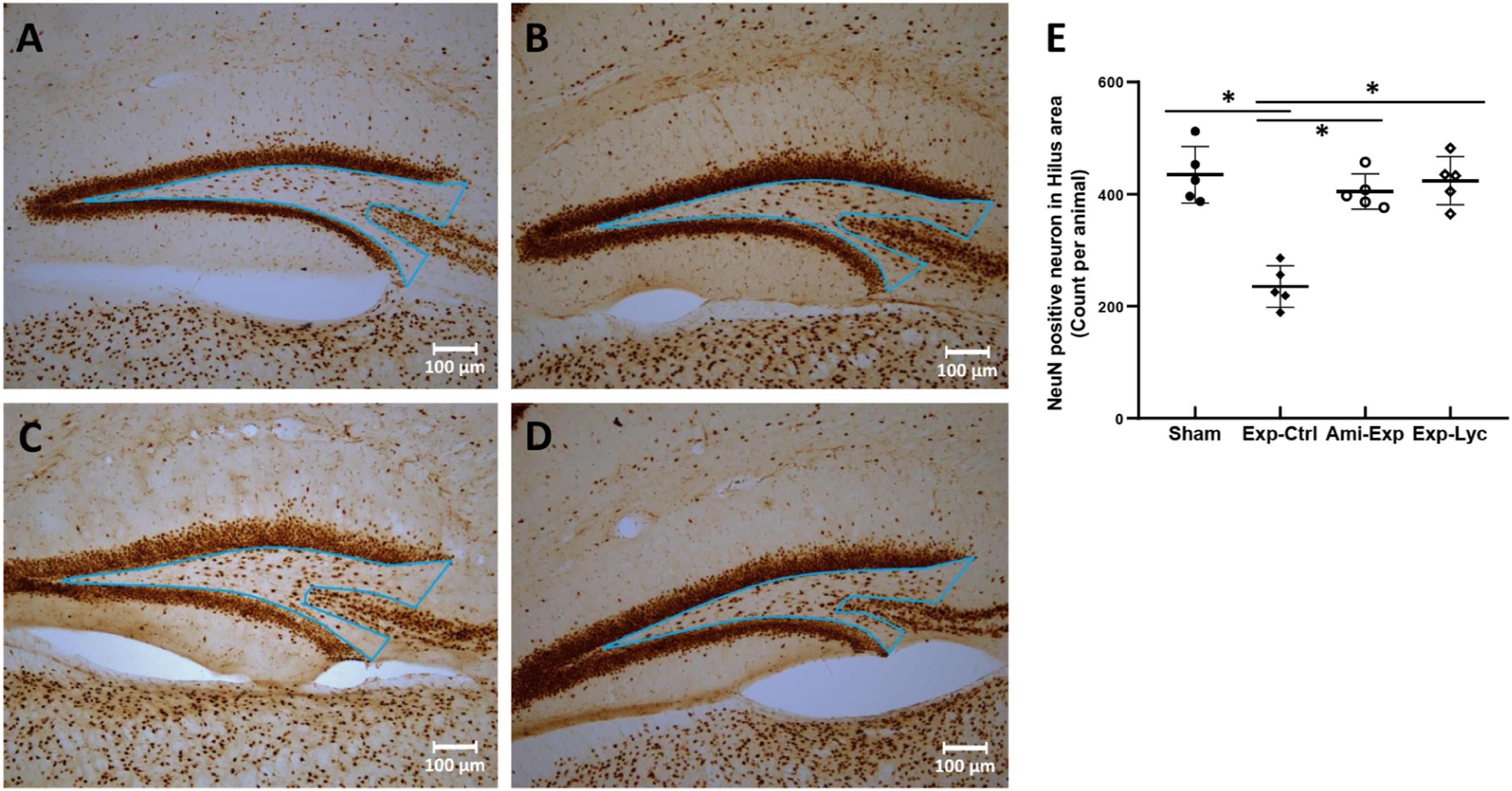


Discussion
Lyc Improves Radiation-Induced Body Weight Loss
Radiotherapy with ionizing radiation may lead to body weight loss in brain or neck cancer patients 25 and animals exposed to acute high dose radiation. 5 Lycium barbarum berry has been increasingly accepted as an anti-oxidative, immune-boosting, and neuroprotective food supplement which may strengthen the body resistance to damages, pathogens, and even ageing. 10 In the present study, oral administration of Lyc effectively prevented weight loss of the mice X-ray-irradiated with 5.5 Gy, indicating that this traditional herbal medicine should have a role in controlling radiation damages. To further study the radioprotective role of Lyc, we investigated its effect on the neurobehavioral performance and neuroprotection on the hippocampal interneurons and hilar cells in the dentate gyrus.
Lyc Improves Radiation-Induced Behavioral Impairments
Our data from open field, forced swimming, and tail suspension tests have shown that mice irradiated with 5.5 Gy suffered less locomotor activity and depression. These results confirmed that the radiation dosage we applied was sufficient to induce the neurobehavioral impairments of the exposed mice. 5 Morris water maze test has suggested the radiation induced spatial cognitive dysfunction. 26 Four-week oral administration of Lyc could effectively prevent those radiation induced behavioral and cognitive impairments. It has been reported that LBP could significantly improve the dextromethorphan-induced depression in the rat model. 27 In the APP/PS1 double transgenic mice, a widely used animal model of Alzheimer’s disease, 2-week oral administration of Lyc significantly improved their spatial memory in Morris water maze test. 28 This radioprotective effect of Lyc should be related to the previously suggested effects of Lyc or LBP, including anti-oxidation, anti-inflammation, preventing neuron loss, and promoting neuronal regeneration. 29 The anti-oxidative effect of Lyc or LBP should be due to their activities in scavenging reactive oxygen species (ROS) and inhibiting oxidases.30-32 By using primary cultured neonatal rat hippocampal neurons, LBP significantly decreased the overexpression of inflammatory cytokines and apoptotic biomarkers induced by oxygen–glucose deprivation/reperfusion. 33 These anti-oxidative and anti-inflammatory effects of Lyc should contribute to its neuroprotection and subsequent improvement of spatial memory and depression.
Lyc Improves Radiation-Induced Hippocampal Neuron Loss
Our previous study has demonstrated that the oral administration of epimedium 5 or intraperitoneal injection of amitriptyline, an inhibitor of acid sphingomyelinase (ASMase), 6 significantly prevent radiation-induced loss of Ki67, doublecortin (DCX), and PV immunopositive cells in the subgranular zone (SGZ) of the dentate gyrus, indicating the improvement of neurogenesis in SGZ. In the pilocarpine induced mouse model of status epilepticus, dramatic reduction of NeuN, CB, and PV immunopositive neurons in different hippocampal areas implicated that these biomarkers should help to present the status of neuron loss and neuron regeneration in hippocampus.34,35 A recent study has reported that LBP treatment could increase the number of NeuN positive neurons in the dentate gyrus of APP/PS1 mice. 18 In this study, our behavioral tests have demonstrated obvious cognitive impairment and depression in those irradiated mice. Therefore, we focused on the changes of those hippocampal neurons and interneurons which should related to the cognitive dysfunction and depression.
The number of NeuN-immunopositive neurons in the dentate hilus could be reduced by ionizing irradiation. 36 In the chronic mouse model of temporal lobe epilepsy (TLE), reduced number of the hilar mossy cells was found to be the major reason of spatial memory deficit. 37 As the major excitatory neurons in dentate hilus, mossy cells are responsible for receiving inputs from local interneurons, granule cells and the CA3 input and protruding axons into different layers of the dentate gyrus.38,39 They play essential roles in maintaining cognitive neuronal network and even in the regeneration and/or the reorganization of neuronal network after neuronal damage.34,40,41 Therefore, the decreased number of hilar NeuN immunopositive cells after irradiation in this study may implicate the mossy cell loss and the consequent cognitive deficit and depression. 41
Several previous studies have demonstrated that the loss of hippocampal interneurons should lead to cognitive dysfunction and behavior alteration.42-44 As a calcium binding protein expressed in some hippocampal interneurons, CB plays a critical role in hippocampal learning function. 45 Down-regulated hippocampal CB expression and/or CB positive neurons were observed in various neurological disorders and were related to cognitive deficits,46-48 while the increased number of CB positive hippocampal neurons implicated the generation of new neurons and the establishment of functional neuronal network. 49 The CB positive interneurons located in the strata radiatum, lacunosum moleculare (CA1-SRLM) and oriens (CA1-SO) of CA1 area are mostly GABAergic interneurons which mainly form synapses with CA1 pyramidal cells and are significantly reduced during ageing.50,51 In the present study, we found that the radiation exposure reduced the number of CB positive interneurons in the CA1-SRLM, which are involved in the regulation of the inputs from the entorhinal cortex to hippocampal CA1 apical dendrites of pyramidal neurons or spatial information. 45 And the reduced number of CB expressing interneurons in the CA1-SO may affect the interaction between the CA1 pyramidal cells and CA1-projecting subiculum neurons which should play a role in the spatial memory.52,53
PV positive cells in hippocampal area are inhibitory GABAergic interneurons which function to configure learning related neuronal network. 54 The PV positive interneurons in CA1 area provide inhibitory synapses onto CA1 pyramidal neurons to regulate their activities in cognitive network.55,56 Promotion of PV immunopositive interneurons in CA1 area could rescue cognitive dysfunction of adult mouse model of schizophrenia. 57 In addition, the activity of PV positive interneurons in dentate gyrus should have an essential role in cognitive function.58,59 The irradiation-induced PV immunopositive neuron loss in the stratum pyramidum of CA1 (CA1-SP) and in the stratum granulosum of the dentate gyrus (DG-SG) was observed in current study, which should be responsible for the cognitive deficits of those irradiated mice.
The Lyc has been reported as an anti-depressive drug previously. 60 Our behavioral tests indicated the spatial memory impairment and depression of those radiation exposure mice. Given these, we proposed that the radiation exposure at the dosage of 5.5 Gy induced the depressive behavior and the spatial memory impairment of mice by promoting the hippocampal NeuN-positive neuron loss in the hilus, CB-positive interneuron loss in the CA1-SO and CA1-SRLM, and the PV-positive interneuron loss in the CA1-SP and DG-SG areas. Oral administration of Lyc significantly prevented the neuron loss in these areas and then maintained the hippocampal neuronal network which consequently improved the radiation induced cognitive deficit and depression.
Conclusion
Ionizing radiation at the dosage of 5.5 Gy induced the depressive behavior and spatial memory impairment in BALB/c mice 4 weeks after irradiation. Significant NeuN-positive neuron loss in the hilus of the dentate gyrus, CB-positive interneuron loss in the CA1-SO and CA1-SRLM, and PV-positive interneuron loss in the CA1-SP and DG-SG areas 6 weeks after irradiation were demonstrated by immunohistochemistry study. Oral administration of Lyc prevented the loss of those hippocampal neurons and then improved the radiation-induced cognitive dysfunction and depression. While further study is still needed to elucidate the molecular mechanism of Lyc, current study strongly suggests that this “superfood” may be a promising radio-neuro-protective supplement to prevent radiotherapy or other ionizing radiation induced cognitive impairment.
Footnotes
Author Contributions
LG, Q-QD, and P-QC: Investigation. T-TY, C-QX, and X-ZL: Formal analysis. X-CP: Resources, Methodology. FQ: Conceptualization, Data Curation, Writing—Original Draft. J-RH: Project administration, F-RT: Conceptualization, Methodology, Writing—Review and Editing.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by grants from the College Students Innovative Entrepreneurial Training Program of Hubei province (S20191049071) to GL, the Scientific Research Foundation of Hubei Health Commission (WJ2016-Y-01), the Jingzhou Science and Technology Department (2019EC61-17) and the Yangtze Fund for Youth Teams of Science and Technology Innovation (2016cqt04) to QF, the nature science foundation of Hubei province (2017CFB786) and the Jingzhou Science and Technology Department (2017-93) to PXC, the Research Project of Traditional Chinese Medicine (WJ2017-15) and the Scientific Research Foundation of Hubei Health Commission (WJ2019-02) to HJR, and National Research Foundation of Singapore to Singapore Nuclear Research and Safety Initiative (TFR).