
Abstract
Acetaminophen (APAP) overdose accounts for the highest incidence of acute liver failure, despite the availability of an antidote i.e. N-acetylcysteine. This calls for alternative strategies to manage APAP-induced liver injury (AILI). Therapeutic hypothermia has been explored in past studies for hepatoprotection, but these phenomenal reports lack clarification of its optimal window for application, and mechanistic effects in specific AILI. Hence, we conducted an in vitro study with transforming growth factor-α transgenic mouse hepatocytes cell line, TAMH, and human liver hepatocytes cell line, L-02, where cells were conditioned with deep (25°C) or moderate (32°C) hypothermia before, during or after APAP toxicity. Cell viability was evaluated as a hallmark of cytoprotection, along with cell death. Simultaneously, cold shock proteins (CSPs) and heat shock proteins expressions were monitored; key liver functions including drug-metabolizing ability and hepatic clearance were also investigated. Herein, we demonstrated significant hepatoprotection with 24-hour moderate hypothermic conditioning during AILI and this effect sustained for at least 24 hours of rewarming. Such liver preservation was associated with a CSP—RNA-binding motif protein 3 (RBM3) as its knockdown promptly abolished the cytoprotective effects of hypothermia. With mild and reversible liver perturbations, hypothermic therapy appears promising and its RBM3 involvement deserves future exploration.
Introduction
Drug-induced liver injury (DILI) has been a recalcitrant problem in the medical industry. In 2018, the United States Food and Drug Administration (FDA) have identified more than 700 drugs responsible for triggering DILI of different severities. 1 This may range from mild asymptomatic biochemical derangements to severe acute liver failure (ALF) where liver transplantation remains the only resort for survival. 2,3 Notably, acetaminophen-induced liver injury (AILI) transcends all other DILI by several folds in causing ALF-related deaths. 4 Its high prevalence, despite the presence of an approved antidote, N-acetylcysteine (NAC), 5 highlights an irony underlying the current regimens for tackling hepatotoxicity. Hitherto, the management of liver injuries mainly advocates 2 key principles—the discontinuation of drug agents likely to cause DILI and the provision of supportive therapy for symptomatic relief. 6 -8 While antidotes such as NAC are available for attenuating liver injury, early administration of NAC would be critical to achieve better outcomes of lower incidence of AILI and in-hospital deaths. 9 Even so, there are repeated reports of variable efficacy, and AILI or ALF may still ensue. 10,11 This, therefore, illustrates the limitations of the current regimens and calls for an urgent need to expand the treatment options for DILI, and more specifically, for the highly pervasive AILI today.
Herein, we aim to address AILI from an alternative perspective, away from a pharmaceutical solution. By casting the safety net for hepatotoxicity management beyond pharmaceuticals, we aim to address the inadequacies with the current pharmacological management. This includes the variable efficacy of antidotes, the dilemma of antidote dose to be administered when medical supervision is not readily available or even in complex cases of combined drugs intoxication or in clinically unstable patients. To do so, we revisited a century-old concept of therapeutic hypothermia (TH). 12 as a potential drug-free approach to ameliorate AILI. By definition, TH refers to a downshift of core body temperature from its physiological range (36.5-37.5°C) to below 35°C. 13 With a gentle cooling to 32°C, moderate hypothermia 14 has been shown to promote positive outcomes in various neurological conditions including cardiac arrest, 15,16 neonatal hypoxic ischemic encephalopathy 17,18 and traumatic brain injury. 19 Most notably, TH has also been recommended by the American Heart Association and the European Resuscitation Council as a standard of care in patients who achieved a return to spontaneous circulation following cardiac arrest. 20,21 These neuroprotective effects have, in fact, been driven by an innate adaptive response to cold stress, involving a class of glycine-rich RNA-binding proteins (RBP), known as the cold shock proteins (CSPs), albeit a potential slowdown of metabolism. Of which, 2 CSPs in the mammalian cells have been most well-characterized—the cold-inducible RNA-binding protein (CIRP) and the RNA-binding motif protein 3 (RBM3). 22,23 These CSPs are strongly induced with mild to moderate hypothermia, between 32-34°C, 24 and regulate post-transcription and translation to mitigate the undesirable biochemical effects following an injury. 25 -27 Since a well-known key response to toxic injury involves RBP-mediated epitranscriptomics regulation, 28 the increased expression of CSPs via TH deserves a thoughtful investigation for resolving AILI.
Past studies have adopted the concept of TH on various types of DILI and hepatoprotection have been demonstrated. 29,30 In particular, Vaquero et al. has also reported on desirable liver recovery following acetaminophen (APAP) overdose in mice. 30 Yet, these studies performed with TH on DILI were mainly to investigate for in vivo protective response at single time and condition of hypothermia while the mechanistic association with the underlying cold-inducible adaptive mechanism involving CSPs and the finding of an optimal protective response of TH have not been clearly established. Hence, to anchor the concept of hepatoprotection with TH beyond phenomenology, we conducted an in vitro investigation to deepen the fundamental understanding of hypothermic effects in the field of liver toxicology. Specifically, this study began with an exploration on the optimal magnitude and timeliness of hypothermic conditioning on AILI, followed by an inquiry of cold-inducible mechanisms governing the hepatoprotection and finally, a safety evaluation of hypothermic effects on important hepatocyte functions was carried out. Ultimately, all these would be crucial to build the foundation for an informed clinical use of TH in AILI and, if any, in DILI in future.
Materials and Methods
Cell Culture and Reagents
Transforming growth factor-α transgenic mouse hepatocytes cell line, TAMH, (a kind gift from the lab of Professor Nelson Fausto, University of Washington) were cultured in Dulbecco’s modified eagle medium: nutrient mixture F-12 (Life Technologies, Carlsbad, CA) supplemented with 10 mM nicotinamide (Sigma-Aldrich, St. Louis, MO), 100 nM dexamethasone (Sigma-Aldrich, St. Louis, MO), 2 µg/mL of gentamicin (Sigma-Aldrich, St. Louis, MO) and 0.1% (v/v) of insulin-selenium-transferrin supplement (Bio Laboratories, Singapore, SG) at 37°C in a humidified atmosphere containing 5% CO2, based on Wu et al. 31 while human liver hepatocytes cell line, L-02, (a kind gift from the lab of Associate Professor Yu Chun Kong Victor, National University of Singapore) were cultured in Dulbecco’s modified eagle medium supplemented with 10% (v/v) fetal bovine serum (GE Healthcare, Pittsburgh, PA) at 37°C in a humidified atmosphere containing 5% CO2. For both TAMH and L-02, passages 22-30 were used for all experiments. APAP (Sigma-Aldrich, St. Louis, MO) stock solution was prepared in phosphate-buffered saline (PBS) at 40 mM and diluted with fresh media into 5 and 10 mM for all experiments. Anti-RBM3, anti-CIRP and anti-HSP70 antibodies were purchased from Abcam (Cambridge, UK) while anti-β-actin was obtained from Santa Cruz Biotechnology (Dallas, TX). The horseradish peroxidase (HRP)-conjugated anti-rabbit and anti-mouse secondary antibodies were purchased from Cell Signaling (Danvers, MA). All other reagents were obtained from Sigma-Aldrich (St. Louis, MO) or Thermo Scientific (Waltham, MA).
Hypothermic Conditioning and Rewarming
Three experimental set-ups were carried out (Figure 1)—first, TAMH were incubated at 25°C or 32°C for 24 h followed by 5 mM APAP treatment for 24 h at 37°C to investigate for any prophylactic effect of hypothermia; second, TAMH were treated with 5 mM APAP for 24 h at 37°C followed by an incubation at 25°C or 32°C for 24 h in APAP-free culture media to investigate if hypothermia could escalate the recovery of a liver injury; lastly, TAMH were concurrently treated with 5 mM APAP for 24 h at 25°C or 32°C to investigate for any liver preservative effects of hypothermia during a DILI. All hypothermic incubation was conducted in a humidified atmosphere containing 5% CO2. Following hypothermic conditioning in each set-up, cells may be subjected to rewarming back to 37°C for 24 h. In the setups with observed cytoprotection in TAMH, the same experimental set-ups were performed on L-02 cells, with 5 mM or 10 mM APAP, to validate the positive outcomes in a human in vitro hepatocyte system.
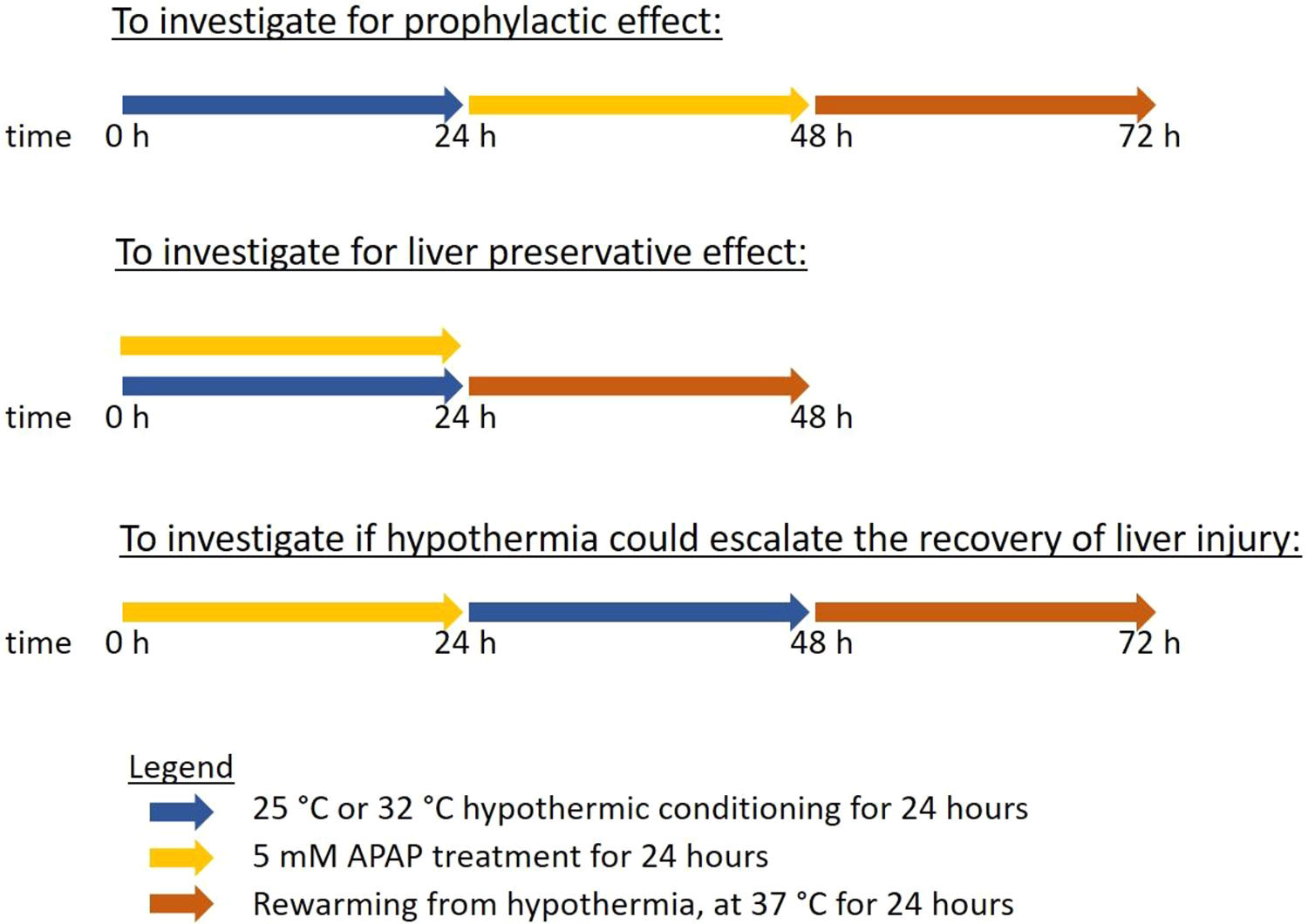
A summary of the 3 hypothermic conditioning procedures relative to the time of APAP administration to evaluate for temporal relationships.
Cell Viability Assay
Cell viability was assessed via 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) (Duchefa Biochemie, Haarlem, NL) assay and CellTiter-Glo® luminescent cell viability assay (Promega, Madison, WI). For the former assay, 1 × 104 TAMH cells per well and 5 × 103 L-02 cells per well were plated in a 96-well plate for 24 h prior to treatment with APAP, hypothermia or N-acetylcysteine (NAC). For NAC treatment, 500 μM and 5 mM of NAC was added 2 hours before APAP treatment and continued for 24 h throughout APAP treatment in TAMH and L-02 cells respectively. Thereafter, 0.4 mg/mL of MTT was added and incubated at 37°C for 4 hours. Following on, the MTT solution was replaced with 50 µL of dimethyl sulfoxide (DMSO) and the absorbance was measured at 570 nm with a microplate reader (Hidex, Turku, FI). Fold changes in absorbance were determined by normalizing against the negative control i.e. APAP-free treatment with continuous 37°C incubation. Each treatment is run in quadruplicates and the entire experiment is triplicated.
For the latter assay, the same cell seeding density was employed on 96-well white plates for both TAMH and L-02. After specific hypothermic conditioning and APAP treatment, CellTiter-Glo® reagent was added in the same volume as the culture medium in each well. Following on, the plates were placed on an orbital shaker at room temperature for 10 minutes before reading the luminescence intensity with a microplate reader. The relative percentage changes in luminescence reading were determined by normalizing against the negative control i.e. APAP-free treatment with continuous 37°C incubation. Each treatment is run in triplicates and the entire experiment is triplicated.
Apoptotic/Necrotic Cell Death Analysis
4 × 105 cells were plated on each 6 cm dish for 24 h prior to APAP treatment at 32°C for 24 h. Following hypothermia or after rewarming for 24 h at 37°C, both live and dead cells were harvested by trypsinization and cell pellets were washed twice with PBS prior to subsequent assay. To determine the percentage of cell death in each sample, cells were stained with 10 µg/mL propidium iodide (PI), diluted in PBS containing 0.05% (w/v) sodium azide and 0.5% (w/v) bovine serum albumin (BSA). On the other hand, to further analyse the extent of necrotic and apoptotic cell death, cells were stained using Annexin-V FLOUS staining kit (Sigma-Aldrich, St. Louis, MO) according to manufacturer’s instructions. Briefly, cells were stained with Annexin V and PI diluted in HEPES buffer and incubated at room temperature for 10 minutes prior to analysis. All cell death analysis was conducted with flow cytometry using CyAn ADP (Beckman Coulter, Brea, CA) and the percentage of PI-stained and/or Annexin V-stained cells were determined based on 20,000 events. A total of 3 biological replicates was performed.
Western Blotting
4 × 105 cells were plated on each 6 cm dish for 24 h prior to APAP treatment at 32°C. Following hypothermia or after rewarming for 24h at 37°C, cell samples were lysed in cell lysis buffer comprising 10 mM sodium fluoride, 1 mM phenylmethylsulfonyl fluoride, 2 M sodium orthovanadate, 2 μg/mL aprotinin, 1% (v/v) octylphenoxypolyethoxyethanol, 0.5% (w/v) sodium deoxycholate and 0.1% (w/v) sodium dodecyl sulfate (SDS) diluted in PBS. 20 µg protein samples were separated via 8% and 15% (v/v) SDS-polyacrylamide gel electrophoresis and transferred onto polyvinylidene fluoride (PVDF) membranes (Bio-Rad, Hercules, CA) for all proteins, with the only exception of CIRP using nitrocellulose membrane (Thermo Scientific, Waltham, MA). All PVDF membranes were blocked with 5% (w/v) BSA in tris-buffered saline containing 0.1% (v/v) Tween-20 (TBS-T) while nitrocellulose membrane was blocked with 5% skim milk in TBS-T. Thereafter, all membranes were incubated overnight at 4°C in primary antibodies diluted in 1:1000, except anti-β-actin antibody which was diluted in 1:10,000. Next, all membranes were washed with TBS-T before incubating with HRP-conjugated secondary antibodies, at 1:10,000 dilution, for an hour at room temperature. The protein bands were then visualized with chemiluminescence image analyzer (G: BOX Chemi XX6, Syngene, Cambridge, UK) using western lightning plus-ECL reagent (PerkinElmer, Waltham, MA). β-actin was used as the housekeeping protein. A total of 3 biological replicates was performed and densitometric analysis was performed with ImageJ (National Institutes of Health, USA) for quantification of western blots.
Real-Time PCR Analysis
4 × 105 cells were plated on each 6 cm dish for 24 h prior to APAP treatment at 32°C for 24 h. After hypothermia or following rewarming for 24 h at 37°C, total RNA was extracted using RNeasy mini kit (Qiagen, Venlo, NL) and quantified with NanoDrop 1000 UV/Vis spectrophotometer (Thermo Scientific, Waltham, MA). cDNA was then synthesized from 1 µg of total RNA using qScript cDNA SuperMix (Quantabio, Beverly, MA) based on manufacturer’s recommendations. The list of primers (Integrated DNA Technologies Coralville, IA) used were shown in Table 1 and β-actin was used as the housekeeping gene. Real-time PCR analysis was performed using QuantiFast SYBR Green PCR Kit (Qiagen, Beverly, MA) on CFX96 touch real-time PCR detection system (Bio-Rad, Hercules, CA) and the cycling conditions were as follow—the samples were heated at 95°C for 5 minutes, followed by 40 cycles of 95°C for 10 s and 60°C for 30 s. The relative mRNA expressions were determined based on fold changes calculated using 2-ΔΔCt, where normalization was performed against the negative control i.e. APAP-free treatment with continuous 37°C incubation. To investigate the inducibility of CYP3A4 during hypothermia and rewarming, 20 µM of rifampicin was added to APAP treatment at 32°C for 24 h or during rewarming at 37°C for 24 h before real-time PCR analysis. All samples were run in triplicates and a total of 3 biological replicates was performed.
Primer Sequences Used in Real-Time PCR Analysis.

Relative Quantification of APAP With LC-MS/MS
4 × 104 cells per well were plated on a 24-well plate for 48 h prior to 5 mM APAP treatment at 32°C for 24 h. At 0, 2, 4, 8, 12 and 24 h of APAP treatment, the media was collected, and cells were trypsinized and collected in a separate tube. For the former, acetonitrile (HPLC grade, Thermo Scientific, Waltham, MA) containing 2 µM of cyclophosphamide (Sigma-Aldrich, St. Louis, MO) as the internal standard (IS), was added to the media in 3:1 for protein precipitation. The mixture was then vortexed for 2 minutes prior to centrifugation at 14,000 rpm for 15 minutes at 4°C. Thereafter, the supernatant was collected and diluted 20X in water before measuring with liquid chromatography tandem mass spectrometry (LC-MS/MS). For the latter, trypsinized cells were lysed with 0.2 N sodium hydroxide, containing 2 µM IS, and subsequently neutralized with equal volumes of 0.2 N hydrochloric acid, containing acetonitrile in 1:4 for protein precipitation. Following on, the lysed cell mixture was centrifuged at 16,000 g for 10 minutes at 4°C and the supernatant was collected for LC-MS/MS run.
The LC-MS/MS setup comprised of a coupled binary ultra-performance liquid chromatography (UPLC) system (Acquity UPLC®, Waters, Wilford, MA) and a triple quadrupole mass spectrometer with electrospray ionization (QTRAP® 3200, Ab Sciex, Framingham, MA). Chromatographic separation of APAP and IS, were carried out using AQUITY UPLC BEH C18 column (2.1 mm × 50 mm, dp = 1.7 µm, Waters, Wilford, MA, USA). The temperature of column and auto sampler rack was set at 45°C and 6°C respectively. The injection volume used was 4 µL and the chromatographic run time was 4.0 min. Gradient elution (Table 2) was employed at a uniform flow rate of 0.5 mL/min, in which the aqueous phase (A) was 5 mM ammonium acetate in water (pH 5) and the organic phase (B) was 5 mM ammonium acetate in acetonitrile (pH 5). The eluent was transferred to the electrospray probe, with a positive ionization mode, after 0.6 min of run.
Gradient Condition for LC-MS/MS Run.

For the mass spectrometer, the ion spray voltage was 4500 V and temperature was 500°C. Curtain gas, collision gas and the ion source gas pressures were 30, 9 and 25 psi respectively. The analyte specific parameters are specified in Table 3.
Analyte Specific Parameters for LC-MS/MS Run.

A total of 3 biological replicates was performed and relative quantification of peak area ratios was performed with MultiQuant software, version 3.0.1 (Ab Sciex, Framingham, MA).
CYP3A4 and GST Activity Assay
4 × 105 cells were plated on each 6 cm dish for 48 h prior to APAP treatment at 32°C for 24 h. Next, after hypothermia or following rewarming for 24 h at 37°C, cells were lysed and the supernatants were collected to measure the activity of CYP3A4 (Abcam, Cambridge, UK) and glutathione-S-transferase (GST) (Abcam, Cambridge, UK) according to manufacturer’s instructions. For CYP3A4 activity, the fluorescence intensity was measured at an excitation wavelength of 535 nm and at an emission wavelength of 587 nm in a kinetic mode for 45 mins while for GST activity, the absorbance was measured at 340 nm in a kinetic mode for 20 mins on a microplate reader. Fold changes in fluorescence intensity, representative of CYP3A4 activity, were determined by normalizing against the negative control i.e. APAP-free treatment with continuous 37°C incubation. A total of 3 biological replicates was performed.
ICG Uptake Assay
8 × 104 cells per well were plated on a 12-well plate for 24 h prior to APAP treatment at 32°C for 24 h. Thereafter, 1 mg/mL of indocyanine green (ICG) was added to each well and incubated at 37°C for an hour. Next, cells were washed with PBS before visualizing the extent of ICG uptake into cells with Olympus microscope (CKX41, Tokyo, JP). Cells were then incubated for another 6 hours at 37°C in fresh media and the extent of ICG release from cells were determined based on the absorbance measured at 780 nm with a microplate reader. Fold changes in absorbance were determined by normalizing against the negative control i.e. APAP-free treatment with continuous 37°C incubation. A total of 3 biological replicates was performed.
RBM3 Silencing With siRNA
To knockdown RBM3, small interfering RNA (siRNA) with RBM3-specific sequences were transfected into L-02 cells. The RBM3-specific siRNA sequences include 5′-GUAUGGAUAUGGAUAUGGAtt-3′ and 5′-UCCAUAUCCAUAUCCAUACcc-3′ while the scrambled siRNA comprises of proprietory sequences, with no match to any genes in the human species. Both siRNA sequences were purchased from Life Technologies (Carlsbad, CA). L-02 cells were plated to reach ∼70% confluence before initiating transfection. To do so, 40 nM of siRNA was transfected with 2% (v/v) lipofectamine 2000 (Invitrogen, Carlsbad, CA) in opti-MEM I media (Thermo Scientific, Waltham, MA) based on manufacturer’s instructions. At 24 hours after transfection, cells were washed once with PBS and treated with hypothermia and/or 10 mM APAP for 24 hours prior to harvesting for further experiments.
Statistical Analysis
All data were expressed as mean ± SD of 3 biological replicates. Statistical analysis was carried out using GraphPad Prism for Windows (version 7.00) (GraphPad Software, La Jolla, CA). Unpaired student t test was performed for all statistical analysis while one-way ANOVA was used to compare the differences in cell viability following NAC and/or hypothermia on AILI. Post-hoc tests were subsequently carried out with correction for multiple comparisons using the Sidak’s method and differences between groups were considered statistically significant for p < 0.05.
Results
Moderate Hypothermic (32°C) Conditioning Promotes Cell Viability of Injured Hepatocytes During AILI
To investigate the temporal relationship between different extents of hypothermic conditioning, i.e. deep (25°C) or moderate (32°C) hypothermia, with the onset of AILI, cell viability was used as the hallmark for cytoprotection. In deep hypothermic conditioning, TAMH alone, without liver injury, displayed lower cell viability across all temporal events while there were unremarkable changes in cell viability following APAP toxicity. As a result, no prophylactic and preservative effect was observed, and it does not escalate recovery after a liver injury too (Figure 2A-C). In contrast, with moderate hypothermia, the cell viability of TAMH in most temporal events were comparable with the respective controls, treated with the same APAP dose. The only exception was observed when moderate hypothermic conditioning (32°C) was introduced during hepatotoxicity—there was a significant improvement in cell viability by ∼1.41 fold (Figure 2C) which denoted the presence of liver preservative effect. In fact, such hepatoprotection was comparable, to a significant extent, as the effect of NAC (∼1.23 fold), the classic rescue therapy administered clinically for APAP overdose (Supplemental Figure 1). Yet, the concomitant administration of NAC and moderate hypothermia does not drive synergistic protective outcomes (Supplemental Figure 1). Upon rewarming, the liver preservative effect of moderate hypothermia persisted with a ∼2.03 fold increase in cell viability significantly (Figure 2C).
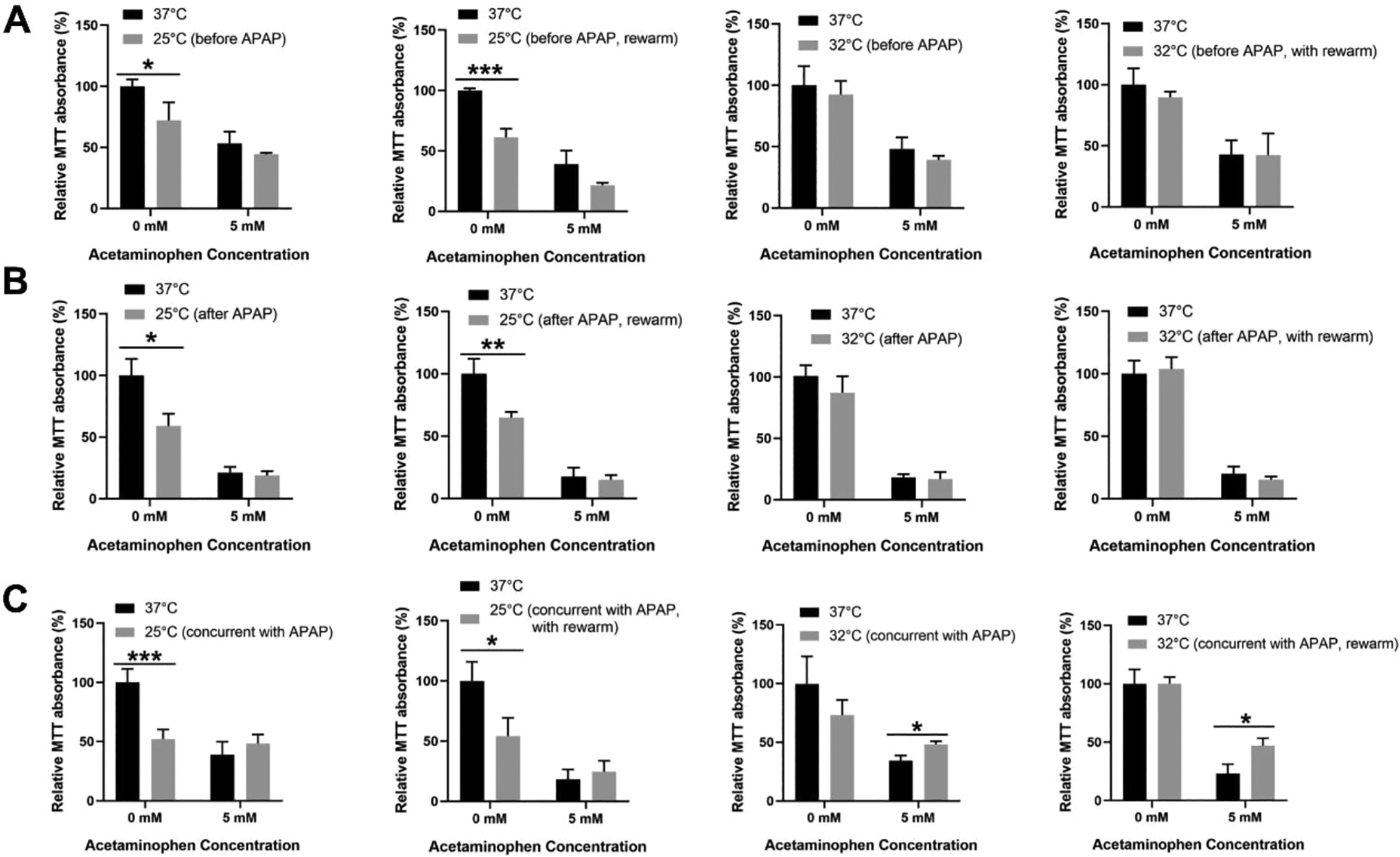
Effect of hypothermia and subsequent rewarming on the cell viability of TAMH. The effects of hypothermia, at 25°C (deep) or 32°C (moderate) for 24 h on TAMH were determined using the MTT assay (A) before, (B) after and (C) during 5 mM AILI followed by a 24 h rewarming back to the physiological temperature at 37°C for each of the above treatment conditions. Data are presented as mean ± SD (n = 3), where unpaired t-test is used to compare the effect of hypothermia or subsequent rewarming on TAMH against their respective controls treated with the same APAP dose. *p < 0.05, **p < 0.01, ***p < 0.001.
Beyond mouse hepatocytes, the cytoprotective effects of hypothermia observed upon a concomitant injury was also validated in the human hepatocytes, L-02, in vitro. During severe 10 mM AILI, cell viability was enhanced with moderate hypothermia by more than 2-fold (Supplemental Figure 2A and B) and a subsequent restoration to physiological temperature continued to sustain the cytoprotection by ∼2.60-2.75 fold (Supplemental Figure 2A and B). Similar to TAMH, hypothermia does not appear to work synergistically with NAC in L-02 cells even though it was shown to alleviate liver injury more prominently than NAC (Supplemental Figure 2C).
Taken together, moderate hypothermia displayed liver preservative effects in the presence of a toxic stimulus and the positive outcomes continued beyond hypothermia and sustained for at least 24 h of rewarming. Furthermore, the gentle downregulation of temperature from 37°C to 32°C could potentially evade unnecessary perturbations to healthy hepatocytes and allow a more targeted response to the site of injury, though this is seemingly unachievable with lower temperatures i.e. 25°C.
Hypothermia Induces RBM3 During Cooling in Hepatocytes
To associate the molecular effects of moderate hypothermia (32°C) with its cytoprotective behavior observed during AILI (Figure 2), the transcriptional and protein expressions of well-known CSPs i.e. RBM3 and CIRP were investigated. With a relative temperature upshift during rewarming, from 32°C back to 37°C, the potential confounding effect of heat shock was also evaluated with HSP70, an inducible HSP expressed in the liver. Indeed, with an initiation of moderate hypothermia at 32°C, both RBM3 transcripts and proteins were strongly induced in comparison with cells kept at physiological temperature (37°C) (Figure 3A and C). On the contrary, CIRP was only induced on a transcriptional level while its protein expressions remained unaltered, and HSP70 transcripts were downregulated albeit unchanged protein levels (Figure 3A and C). Thereafter, with rewarming, dynamic transcriptional regulation was apparent as the transcript expressions of both CSPs promptly declined to a comparable level as their respective control cells treated with the same APAP dose at 37°C (Figure 3B). HSP70 transcript levels was, however, restored back to similar levels as the control cells, without a significant induction during rewarming (Figure 3B). Of note, all these were accompanied with comparable protein expressions of CSPs and HSP from the cooling phase (Figure 3C). Seemingly, the prolonged half-life of these proteins may have cushioned the potential disruptions following temperature transitions. Beyond TAMH, similar trends in protein expressions were also observed in human hepatocytes, L-02 (Supplemental Figure 3).

Effect of hypothermia and its subsequent rewarming on the transcriptional and protein expressions of RBM3, CIRP and HSP70. Quantitative analysis with real time RT-PCR was performed on TAMH subjected to (A) moderate hypothermia, at 32°C, for 24 h during 5 mM AILI and (B) after 24 h rewarming back to the physiological temperature at 37°C. Western blotting was also carried out to determine the corresponding protein expressions of RBM3, CIRP and HSP70 during (C) moderate hypothermia and (D) following rewarming. β-actin is used as the housekeeping gene for both RT-PCR and western blotting. Data are presented as mean ± SD (n = 3), where unpaired t-test is used to compare the effect of hypothermia or subsequent rewarming on TAMH against their respective controls treated with the same APAP dose. *p < 0.05, **p < 0.01, ***p < 0.001.
Ensuing these molecular changes in hypothermia and rewarming, the cytoprotective effects may primarily be influenced by the more temperature-responsive RBM3 CSP, rather than CIRP. Furthermore, heat shock does not appear to play a role in conferring hepatoprotection via HSP70 in both cooling and rewarming phase.
Next, to strengthen the association between active RBM3 induction and the cytoprotective phenomenon observed in AILI, the impact of temperature downregulation on the in vitro thermodynamics was examined. This includes an evaluation of how a reduction in temperature would affect the rate of diffusion of APAP from the media into the cells, as well as, the rate of APAP metabolism in the hepatocytes since APAP toxicity is a result of an accumulation of its reactive metabolite, N-acetyl-p-benzoquinone imine (NAPQI) in the cells. 32 Accordingly, across different timepoints within 24 h of APAP treatment, there is unremarkable, though marginal, difference in the level of APAP remaining in the media and in the cells (Figure 4). In other words, the rate of diffusion of APAP into the cells and the rate of APAP metabolism are seemingly comparable between moderate hypothermia and physiological temperature in a hepatocyte cell system. Therefore, this largely dismisses the notion in which moderate hypothermia may have driven protection by a slowdown in the general thermodynamics. Rather, the liver preservative effects may have closer association with biochemical changes, possibly brought about by CSPs, such as RBM3.

Extent of APAP depletion following moderate hypothermic conditioning. Relative quantification of APAP levels remaining (A) in the media and (B) in the cells at 0, 2, 4, 8, 12 and 24 h after TAMH was treated with 5 mM APAP, was performed with LC-MS/MS. 2 µM cyclophosphamide was used as the internal standard. Data are presented as mean ± SD (n = 3), where unpaired t-test is used to compare the effect of hypothermia on TAMH against their respective controls at each timepoints.
With the recurrent correlation observed between RBM3 upregulation and cytoprotection in AILI, the study went further to uncover its direct relationship through knockdown of RBM3 in the human hepatocytes, L-02 (Figure 5). By suppressing the upregulation of RBM3 following hypothermia in AILI, it promptly eliminates the hepatoprotective effects, with no significant improvement in cell viability observed (Figure 5B). This clearly demonstrated an explicit cytoprotective role of RBM3 in APAP toxicity following moderate hypothermia.
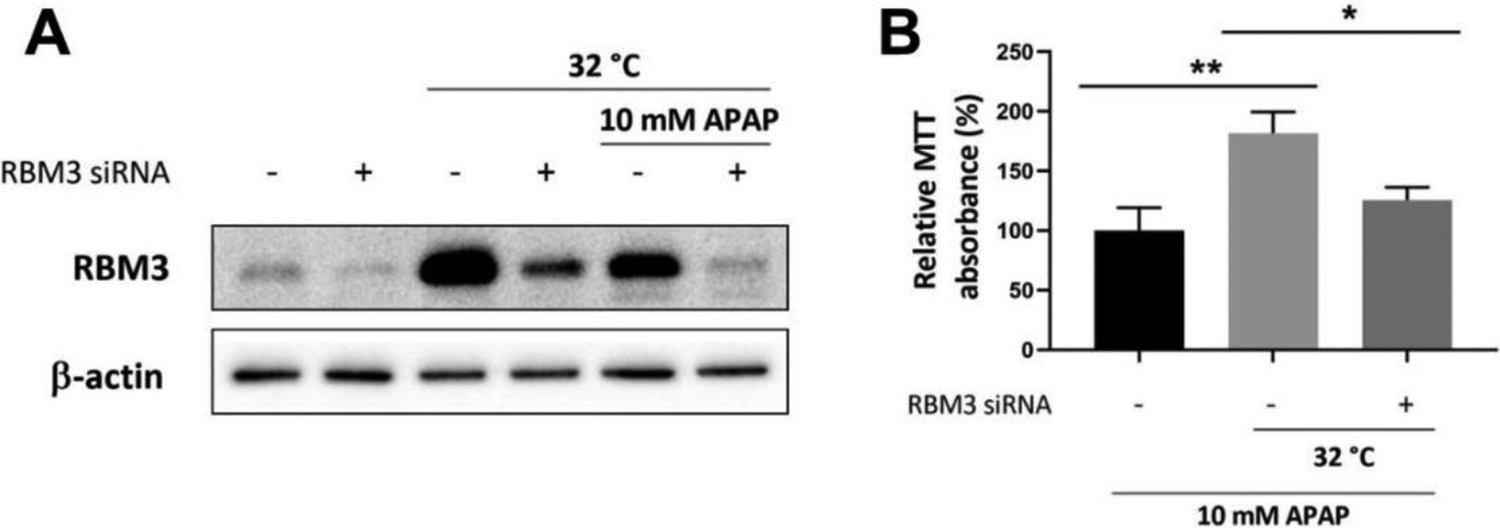
Effect of RBM3 silencing on cell viability upon APAP toxicity in L-02. A, Western blotting was performed to confirm the silencing effect of RBM3 siRNA at 37°C control set-up, in the presence of moderate hypothermia alone at 32°C and with moderate hypothermic conditioning during 10 mM AILI. B, Changes in cell viability following RBM3 silencing, was assessed with MTT assay. Data are presented as mean ± SD (n = 3), where unpaired t-test is used to compare the effect of RBM3 silencing on L-02 against the respective control treated with the same APAP dose and hypothermia. *p < 0.05, **p < 0.01.
Moderate Hypothermia Attenuates Cell Death Through Distinct Mechanisms During Cooling and Upon Rewarming
Beyond elucidating cytoprotective effects, cell death analysis was also performed to examine the extent and mode of cell death following AILI and/or hypothermia. While necrosis has been commonly recognized as the main mode of cell death in APAP toxicity, 33 little has been known regarding the specific effects of hypothermia and/or rewarming on evading cell death. Here, we demonstrated a significant ∼2.82-fold reduction in total cell death during hypothermia (Figure 6A) and this predominantly involved an attenuation of necrotic cell death, as illustrated with a reduction in cells stained with both Annexin V and PI dye in the top right quadrant of the dots plot (Figure 6C). Following temperature restoration from hypothermia, with the removal of APAP toxic insult after 24 hours, total cell death was also abated by ∼1.93-fold (Figure 6B). Interestingly, this was accompanied with a prominent attenuation of early apoptotic cell death instead, as denoted by a reduction in cells stained only with Annexin V in the bottom right quadrant of the dots plot (Figure 6D). Together, these underscored the stark contrast in the mechanism alleviating APAP toxicity following the cooling and rewarming phase. On a similar note in L-02 human hepatocytes, a remarkable fall in cell death by ∼2.36 and ∼1.76-fold during moderate hypothermia and rewarming in 10 mM AILI were also observed respectively (Supplemental Figure 4).

Effect of hypothermia and its subsequent rewarming on the cell death profile of TAMH. Hypothermia, at 32°C (moderate), was performed for 24 h on TAMH during 5 mM AILI followed by a 24 h rewarming back to the physiological temperature at 37°C. At both (A) hypothermic and (B) rewarming state, percentage of total cell death was determined with PI staining. Further analysis on the extent of necrotic and apoptotic cell death was also conducted with Annexin V and PI staining during (C) hypothermia and (D) rewarming. The top right quadrant, in blue, represents the extent of necrotic cell death while the bottom right quadrant, in green, represents the extent of apoptotic cell death. The relative extent of cell death, in percentage, has been indicated in each of the quadrants. Data are presented as mean ± SD (n = 3), where unpaired t-test is used to compare the effect of hypothermia or subsequent rewarming on TAMH against their respective controls treated with the same APAP dose. **p < 0.01.
Moderate Hypothermia Displays Varying Effects on Hepatic Metabolism and Clearance Capacity, Which Are Mostly Reversible Upon Rewarming
To holistically examine the moderate hypothermic effects in AILI, its safety profile in hepatocytes was characterized alongside its efficacy. To do so, principal hepatocellular functions of metabolic and clearance capacity, were investigated. For the former, transcriptional changes and activities of key phase I (CYP3A4) and phase II (GSTP1) drug-metabolizing enzymes were measured. Interestingly, hypothermia alone, in the absence of APAP toxicity, were observed to mediate prominent reductions in CYP3A4 and GSTP1 transcript expressions even though their activities remained well preserved (Figure 7). Following restoration to physiological temperature, these transcriptional effects were, however, rapidly attenuated (Figure 7A and B). In the presence of 20 μM rifampicin, a CYP3A4 inducer, CYP3A4 remained inducible across hypothermia and rewarming, albeit a weaker induction (Figure 7A). In contrast, with APAP toxicity, prominent reduction in GSTP1 transcript levels were observed together with a higher activity upon cooling; there were, however, comparable inducibility of CYP3A4 regardless of hypothermia (Figure 7). Collectively, the effect of moderate hypothermia may vary vastly across different classes of drug-metabolizing enzymes and could be dependent on the health status of cells. Nonetheless, most of these effects on drug metabolism may dwindle following an inevitable rewarming phase from transient hypothermia.

Effect of hypothermia and its subsequent rewarming on the drug-metabolizing function and hepatic clearance capacity of TAMH. Hypothermia, at 32°C (moderate), was performed for 24 h on TAMH during 5 mM AILI followed by a 24 h rewarming back to the physiological temperature at 37°C. Transcriptional expressions of (A) CYP3A4, in the absence and presence of 20 μM rifampicin, and (B) GSTP1 were determined with real time RT-PCR in TAMH subjected to hypothermia or upon rewarming. β-actin is used as the housekeeping gene. Drug-metabolizing activity of (C) CYP3A4 and (D) GST were examined in both hypothermic and rewarming state while (E) the extent of ICG uptake and release in TAMH was investigated in hypothermia. For CYP3A4 activity assay, TAMH treated with CYP3A4 enzymes was used as the positive control while TAMH treated with 150 µM ketoconazole, a CYP3A4 inhibitor, was used as the negative control. For GST activity assay, TAMH treated with GST enzymes was used as the positive control. Scale bar = 50 μm. Data are presented as mean ± SD (n = 3), where unpaired t-test is used to compare the effect of hypothermia or subsequent rewarming on TAMH against their respective controls treated with the same APAP dose. *p < 0.05, **p < 0.01, ***p < 0.001.
Next, the hepatic clearance capacity was evaluated with the uptake and release of ICG, a chemical which is actively cleared through the hepatocytes. By visual assessment of ICG uptake with microscopy, moderate hypothermia appeared to promote cellular uptake of ICG (Figure 7E). While ICG release from hepatocytes was insignificant between cells incubated at moderate hypothermia and at physiological temperature, the modest impairment on hepatic clearance during hypothermia could not be discounted, considering an apparent higher cellular uptake of ICG.
Discussion
The potential severity and fatality of DILI, alongside its variability in occurrence, have rendered DILI a protracted concern. Of which, APAP overdose as a leading cause of DILI, and being inadequately managed with the current strategies, warrants the need for a sustainable solution. To do so, we build on past in vivo evidences of attenuating DILI with TH 29,30 and attempt to uncover its optimal protective response for AILI. By refining the concept of hypothermia for hepatoprotection, we seek to variegate the possibilities of handling AILI and other acute liver injuries, where non-pharmacological therapy may empower early interventions in a non-invasive manner and avert potential concerns for drug-drug interactions.
In this study, 2 cell lines were employed to cross-validate the hypothermic effects in AILI—mouse hepatocytes, TAMH and human hepatocytes, L-02. While TAMH is of mouse species origin, it is chosen for its comparable cytochrome P450 enzyme expressions with wild-type liver or primary mouse hepatocytes, which renders it desirable for the study of AILI where toxicity arises from the accumulation of its active metabolite, NAPQI. 34,35 In fact, it has been frequently reported as a cell model for studying AILI and it displayed distinct features of APAP toxicity. 36,37 Furthermore, to ensure translatability of results beyond mouse species, hypothermic effects were once again examined in human hepatocyte model, L-02. As a non-cancerous hepatocyte cell line, the changes observed in L-02 following hypothermia and/or AILI would more realistically represent an in vivo human system. Its typical hepatocyte morphology and phenotypes have been closely monitored and maintained across passages to ensure validity of results.
Two hypothermic conditions were under investigation—moderate (32°C) and deep (25°C) hypothermia. While past studies exploring TH in DILI were mostly conducted in the moderate hypothermic range (above 32°C) and proven to be beneficial, a deeper cooling (up to 25°C) has also demonstrated efficacy in other organ toxicity models besides the liver. 38 -40 Hence, to discern the optimal therapeutic window of hypothermia for AILI, a widened scope of investigation was performed, together with a concomitant evaluation of its temporal effects relative to the time of liver injury. Specifically, we conditioned TAMH and L-02 hepatocytes to hypothermia before, during and after AILI (Figure 1) and we have demonstrated, for the first time, a time-dependent liver preservative effect of moderate hypothermia (32°C) during AILI. Although cell line variations may result in differential susceptibility to toxic doses of APAP and hypothermia, with L-02 displaying cytoprotection at 10 mM APAP dose instead of 5 mM (Supplemental Figure 2), the observed hepatoprotection during AILI remains consistent across our study. Moreover, such rescue therapy was comparable, or potentially more superior, to that of NAC, the well-known antidote for APAP overdose (Supplemental Figure 1 and 2C). We note the minor, yet significant, improvement in cell viability following NAC administration in vitro was similarly demonstrated by Tobwala et al. in APAP toxicity. 41 Unlike commonly perceived efficacy of NAC as a clinically useful antidote, its vacillating effect, despite early administration, further rationalize the need to restructure and implement other strategies to better manage AILI. Therefore, the cytoprotective effects of moderate hypothermia presents a non-pharmacological alternative worthy of further exploration. The demonstrated improvement in cell viability through 2 distinct assays i.e. MTT and CellTiter-Glo® assay in TAMH (data not included for CellTiter-Glo® assay) and L-02 further corroborates the observed cytoprotective phenomenon, eliminating potential confounders in cell lines or specific assays. While moderate hypothermia in AILI may behave paradoxically from its expected thermodynamic behavior at a reduced temperature—where cells should experience a subdued metabolism and hence, retard proliferation, 42 there are studies suggesting otherwise. For instance, both Wellmann et al. and Venugopal et al. have reported a strong positive association between RBM3 expressions and cell viability 43,44 ; CIRP has also been shown to improve cell proliferation via the regulation of cyclin E1 and through the phosphorylation of extracellular signal-regulated kinase 1 and 2 (ERK1/2). 45,46 The increased expressions of RBM3 and CIRP may, hence, be crucial factors to promote cell viability despite an expected modest reduction in thermodynamic rate following moderate hypothermia. On this note, our study has displayed a prominent induction of both RBM3 and CIRP on a transcriptional level following moderate hypothermia (Figure 3). The protein levels of CIRP, however, remained relatively constant after 24 hours of cooling while RBM3 protein was upregulated with moderate hypothermia (Figure 2 and Supplemental Figure 3). This could be attributed to a distinct 2-stage transcriptional and translational regulation specific for CIRP, unlike RBM3. Reportedly, the presence of multiple transcription start sites on CIRP have enabled a mixed expression of CIRP transcripts with long and short lengths of 5′-untranslated region (5′-UTR). 47 For prolonged hypothermia beyond 6 hours, there is a documented upregulation of long 5′-UTR CIRP transcripts owing to its mRNA stability in TH. However, it has an internal ribosome entry site (IRES)-like activity unresponsive to temperature. 47 This greatly limits its translational efficiency despite a transcriptional induction. In contrast, RBM3 could exhibit an almost 5-fold increase in IRES-like activity during TH to facilitate an increased protein expression through a cap-independent translation. 48 Henceforth, we postulate that a stark difference in the translational efficiency between different CSPs, rendering RBM3 as the more dominant CSP to mediate the cytoprotection observed in AILI. Albeit its indiscriminate upregulation regardless of toxic response, its distinct induction amid a global suppression of protein synthesis during temperature fall, may confer RBM3 a unique role in hypothermia. 49 Therefore, following its cytoprotective effects exerted in the presence of AILI, we postulate the possibility of other epitranscriptomic drivers, working alongside RBM3, to precisely influence its targeted response on injured cells in liver toxicity. For example, Pilotte et al. has reported on the role of RBM3 in regulating miRNA biogenesis through a direct interaction with ∼70 nt pre-miRNAs. 26 Similarly, Dresios et al. has also demonstrated the binding potential of RBM3 with 60 S ribosomal subunit to influence miRNA levels. 27 In the case of AILI, the exact machinery of epitranscriptomic regulation by RBM3 has, however, remained unknown till today. Nonetheless, its active role in AILI may worth future investigation, especially when the occurrence of cytoprotection, in this study, has precluded the passive effect of a slowdown thermodynamics following a cooling phase (Figure 4), and the significance of RBM3 upregulation has been justified through RBM3 knockdown experiments (Figure 5). Of note, while the thermokinetics behavior in a complete human system may still experience a global slowdown of metabolic rate i.e. reduced volume of distribution and reduced clearance, 50 the controlled in vitro setup here can dispel confounding effects and allow a direct evaluation of the underlying driving forces of protection.
Next, ensuing the transient hypothermic conditioning is the inadvertent restoration to physiological temperature. In line with Neutelings et al. who reported worsening cellular outcomes following rewarming from 25°C, 51 a global suppression of cell viability was also observed in this study when TAMH was rewarmed from deep hypothermia, at 25°C (Figure 2). More importantly, this was less pronounced in cells rewarmed from moderate hypothermia, at 32°C (Figure 2) In fact, there was a continued improvement in cell proliferation following a temperature upshift from moderate hypothermia after AILI i.e. 32°C to 37°C (Figure 2C, and Supplemental Figure 2). These contrasts may have stemmed from a difference in the temperature gradient employed for rewarming. Of which, a steeper gradient of 25-37°C may result in greater undesirable perturbations to the cellular system. Likewise, Bhalala et al. has also suggested the potential of a gentle rewarming gradient to incur less rewarming-associated neurologic injury following a cardiopulmonary bypass. 52 Intuitively, the choice of hypothermic conditions may, therefore, influence the direct effects during hypothermia and more so, the downstream outcomes upon rewarming. As demonstrated, moderate hypothermia (32°C) displayed prominent hepatoprotection across both cooling and rewarming phase unlike deep hypothermia (25°C). This demarcates a therapeutic window where extreme temperature reductions of 25°C and lower would be undesirable while modest temperature downregulations are preferred for attenuation of liver injuries.
Another area of scrutiny in the rewarming phase involves an evaluation of HSP70, an inducible HSP in the liver upon heat stress. 53 Although rewarming only involves a relative temperature upshift to normothermia, the induction of HSP70, like in heat stress, was previously reported. 51 Herein, we showed unchanged HSP70 protein levels both during hypothermia and rewarming (Figure 3C), though there is a significant downregulation in its transcript levels upon cooling (Figure 3A). Notably, this discrepancy was documented by Lewis et al. too, as he proposed a differential temperature preference for the induction of HSP70 mRNA and proteins. 54 Notwithstanding this variable HSP70 levels, the apparent absence of its upregulation during rewarming would exclude its cytoprotective role during a temperature upshift. Instead, the persistently high RBM3 protein levels, which continued from cooling to rewarming (Figure 3C), may better correlate with the extended hepatoprotection observed beyond hypothermia. Its prolonged stability, regardless of temperature transitions, has further accentuated the potential of moderate hypothermia as a drug-free solution for AILI.
The other key aspect of hepatoprotection being inquired in moderate hypothermia, besides cell viability, was the extent and mode of cell death following AILI. Being one of the main yardsticks for measuring the severity of APAP toxicity, we seek to examine the role of hypothermia in allaying APAP toxicity for liver preservation. For the former, there was a significant reduction in cell death across both hypothermia and rewarming phase (Figure 6 and Supplemental Figure 4); for the latter, this suppression of cell death was driven by vastly different mechanism—with a predominant suppression of necrotic- and apoptotic-mediated cell death during cooling and rewarming respectively (Figure 6). Since AILI is known for driving necrotic cell death, 33 the hypothermic effects become directly relevant and targeted for APAP toxicity. In contrast, even though apoptosis does not largely evoke cell death in AILI, the stark difference in the mode of hepatoprotection during the rewarming phase may encourage the versatile use of TH for different causes of liver injuries, other than APAP. 55
Lastly, we went further to explore its impact on the key functions of hepatocytes, namely its drug-metabolizing ability and its clearance capacity. With a subtle temperature downshift in moderate hypothermia, signature phase I (CYP3A4) and phase II (GSTP1) drug-metabolizing enzymes deviated largely in their behavior, with greater perturbation seen with GSTP1 in injured hepatocytes (Figure 7). Without AILI, hypothermia alone suppressed the expressions of CYP3A4 and GSTP1, even though there were unremarkable changes in their enzymatic activities and CYP3A4 remained inducible (Figure 7). As reported by Legendre et al., various drug-metabolizing enzymes could display temperature-dependent expressions, 56 hence a reduced transcript level of CYP3A4 and GSTP1 was expected. Yet, their comparable drug-metabolizing activity could potentially be fueled by the following—First, CYP3A4 has a long documented half-life of 26 to 79 hours in vitro 57 and this may compensate for the transcript changes following 24 hours of hypothermia in this study. Next, while GSTP1 has a much shorter half-life of 19 to 51 minutes, 58 the measurement of a combined activity of all GST might have masked the potential changes in specific GSTP1 activity, among a total of 22 enzymes within the GST family. On a similar note, with AILI, a higher GST activity following hypothermia (Figure 7D), despite a repressed mRNA expression of GSTP1, may suggest a compensatory increase in activity of other GSTs instead, rather than GSTP1. Nonetheless, this was, in coherence, with Hackenhaaret al. who also reported an increased GST activity following 33°C hypothermia in post cardiac arrest patients for up to 72 hours. 59 Since an increased GST activity has been commonly associated with reduced oxidative damage 60 and AILI is largely mediated by towering oxidative stress, 61 this further substantiates the potential molecular effects of hypothermia desirable for alleviating AILI, rather than a subdued thermodynamic rate. While the functional impact of moderate hypothermia on liver metabolism appears variable, most perturbations which occurred during cooling are rapidly reversible with a 24-hour rewarming (Figure 7). Beyond drug-metabolizing function, the hepatic clearance capacity was not largely impaired after hypothermic conditioning too (Figure 7E). Hence, although moderate hypothermia may adversely affect the key liver functions, the impact appeared, mostly, mild and transient. Consequently, this gives moderate hypothermia the edge for clinical use as a drug-free therapy for AILI.
Conclusion
Taken together, in accordance with the in vitro data from both mouse and human hepatocytes, we have uncovered an optimal window for liver protection with TH—where moderate hypothermia, conditioned concomitantly with APAP toxicity, could alleviate AILI with an extended liver protection for at least 24 hours of rewarming. Its liver preservative role for rescuing liver injury, rather than being prophylactic or rehabilitative, may confer practicality for handling hepatotoxicity which are more often than not, acute and inadvertent in nature. Furthermore, these hepatoprotections are strongly associated with the upregulation of RBM3 during cooling and rewarming phase and this is worth future investigations to uncover its mechanistic role in attenuating liver injury. Finally, with the demonstrated efficacy and reversible adverse effects associated with moderate hypothermia, we have made a leap to unveil the possibility of fostering TH as a drug-free clinical therapy for AILI.
Supplemental Material
Supplemental Material, Supplemental_material_DOS - Moderate Hypothermia Effectively Alleviates Acetaminophen-Induced Liver Injury With Prolonged Action Beyond Cooling
Supplemental Material, Supplemental_material_DOS for Moderate Hypothermia Effectively Alleviates Acetaminophen-Induced Liver Injury With Prolonged Action Beyond Cooling by Yeong Lan Tan, Siew Min Tey and Han Kiat Ho in Dose-Response
Footnotes
Acknowledgments
This research was supported by the NUS Department of Pharmacy and NUS Graduate School for Integrative Sciences and Engineering (NGS).
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the Singapore Ministry of Education Academic Research Grants [R148-000-272-114] and [R148-000-295-114].
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.