
Abstract
Radiation biodosimeters are required urgently for fast and accurate evaluation of absorbed dose for irradiated individuals. Lipidomics has appeared as a credible technique for identification and quantification of lipid for researching biomarker of diseases. We performed a lipidomic profile on mice serum at time points of 6, 24, and 72 hours after 0, 2, 5.5, 7, and 8 Gy irradiation to select radiation-responsive lipids and conducted Kyoto Encyclopedia of Genes and Genome pathway enrichment analysis to recognize the pathways and network changes. Then, Pearson correlation analysis was performed to evaluate the feasibility of radiation-responsive lipids to estimate radiation dose. Seven radiation-responsive lipids including PC (18:2/18:2), PC (18:0/18:2), Lyso PC 18:1, PC (18:0/20:4), SM (D18:0/24:1), PC (16:0/18:1), and Lyso PC 18:2 were identified in which glycerophospholipid metabolism presented as the most significant pathway, and they all presented good linear correlation with the irradiated dose. This study identified 7 radiation-responsive lipids in mice serum and certificate their feasibility of dose estimation as biodosimeters.
Keywords
Introduction
In potential widespread radiation accidents such as nuclear and radiological events, biodosimeters of high throughput and high accuracy are needed to fast evaluate the absorbed dose of the wounded. 1 There are some approaches of biodosimeter including measuring the direct radiation-induced changes such as stable-free radicals and measuring biological responses to the radiation damage such as dysfunctional genes, proteins, and metabolites. 1 -5 Nowadays, more and more efforts have been put into metabolomics (analysis of molecules <1 kDa) as an approach to find biomarkers of diseases and body dysfunction including radiation damage by detecting biofluids such as urine, serum, and saliva. 6 And lipidomics is considered as a branch of metabolomics, lipid-targeted metabolomics, which has been used to recognize biological changes in lipid level. 7
Lipids are an essential component of biological membranes and play crucial roles in biological systems including making the cell comparatively independent of the exterior environment by lipid bilayer structures, providing hydrophobic medium for the functional performance and interactions of membrane proteins, and producing second messengers by enzyme reactions. 8 On the basis of the diversity of chemical structure and the hydrophobic and hydrophilic elements, there are 8 categories of lipids including glycerolipids, saccharolipids, sphingolipids, glycerophospholipids (GPs), sterols, polyketides, fatty acyls, and prenols. 9,10 Furthermore, exploring lipid biochemistry by lipidomics not only inquire into the unique functions of lipid molecular species but also investigate potential biomarkers of diseases. The phospholipidome of human serum ferritin (SF) acted as a potential biomarker for the diagnosis of Parkinson disease, and the dysregulated ethanolamine plasmalogens, particularly those with polyunsaturated fatty acids in the circulatory system, was considered to be connected with neurodegeneration. 11 Eicosanoid oxygenation by lipoxygenase, CYP-450, and cyclooxygenase pathways resulted in inflammation and are considerable biomarkers of tissue damage. 12 The deficient plasmalogen was the marker of increase oxidative stress and peroxisomal disorders. 13
Several studies have investigated the relationship between lipid level in serum/plasma and radiation. A phospholipids profiling analysis of rat plasma after γ-irradiation exposure indicated that ionizing radiation could disorder phospholipid metabolism, as phosphatidylethanolamine (PE) and phosphatidylserine (PS) increased remarkably. 14 With the nonhuman primate model, a significant increase was observed in the level of polyunsaturated fatty acids at 7 days after 10 Gy irradiation, including 20:4 (arachidonic acid) and 22:6 (docosahexaenoic acid) acyl moieties. 4 The subsequent study with the same pattern found increases of PC (38:6), ePC (40:3), and (40:5). 15 After 6.5 Gy irradiation, the rise levels of LysoPCs and reduced levels of SMs would be considerable markers between 2 and 3 days postirradiation. 16
In the current study, we performed a lipidomic profile on mice serum at time points of 6, 24, and 72 hours after 0, 2, 5.5, 7, and 8 Gy irradiation to select potential lipid biomarkers for radiation biodosimeters. Perturbations of SMs, PEs, PCs, LPEs, and LyPs were detected overall. Partial least squares-discriminant analysis (PLS-DA) clearly separated subject in 8 Gy groups from 0 Gy groups, which was an essential dose for selection of radiation-responsive lipids. Seven radiation-responsive lipids including PC (18:2/18:2), PC (18:0/18:2), Lyso PC 18:1, PC(18:0/20:4), SM (D18:0/24:1), PC (16:0/18:1), and Lyso PC 18:2 were identified, then we performed Kyoto Encyclopedia of Genes and Genome (KEGG) pathway analysis to recognize the pathways and network changes. Ultimately, Pearson correlation analysis was conducted to evaluate the ability of the 7 lipids to estimate radiation doses as radiation biodosimeters.
Materials and Methods
Chemicals
Standards for lipidomics included 19:0 Lyso PC, 19:0 PC, 17:0 PE, and 12:0 SM (Avanti Polar Lipids, Inc, Alabaster, Alabama). All reagents were Optima LC/MS grade (Fisher Scientific, Pittsburg, Pennsylvania) and all standards were of the highest purity available.
Mouse Model, Radiation, and Serum Collection
Male C57BL/6J mice (6-8 weeks old) were purchased from and raised in the Academy of Military Medical Sciences (Beijing, China). Mice were irradiated in 0, 2, 5.5, 7, and 8 Gy by 60Co source γ-ray at a dose rate of 101.90 cGy/min. Blood was collected from the orbital plexus at different time points (6, 24, and 72 hours) after irradiation. Then serum was separated by centrifugation at 3000 rpm/min for 5 minutes at 4°C and stored at −80°C. Five mice were consisted in each group. Animal care and handling were performed in accordance with the “Guide for the Care and Use of Laboratory Animal of AMMS in China” and all animal experiments were approved by the Animal Care and Use Committee of the Beijing Institute of Radiation Medicine (Beijing, China).
Sample Preparation and Analysis
Serum samples (10 μL) were extracted with cold chloroform:methanol (100 μL, 2:1) containing internal standards, incubated for 30 minutes at 4°C, vortexed for 20 seconds, and centrifuged for 3 minutes (7800g, 4°C). The lower organic phase was removed with a glass pipette, evaporated under N2, stored at −20°C and reconstituted in 20 µL isopropanol:acetonitrile (1:1). Samples were injected (2 μL) into SCIEX Triple TOF 5600 System (SCIEX, Framingham, Massachusetts) equipped with a Waters XBridge Peptide BEH C18 (Milford, Massachusetts) (2.1 mm × 100 mm; 3.5 μm) column with the temperature set at 40°C and a flow rate of 0.40 mL/min. And both positive and negative electrospray ionized lipid species were performed.
Data Processing, Statistics, and Marker Validation
PeakView workstation (version 1.2; SCIEX) was used to check lipid mass spectrometry (MS/MS) information, and MultiQuant software (version 2.1; SCIEX) was used to obtain lipid peak area. In this study, subsequent data analysis was achieved based on positive electrospray ionized lipid species. Principal component analysis (PCA) was used to observe the relationships between different radiation dose groups. A 2-sample Student t test (2 tailed) and PLS-DA were used to select radiation-responsive lipids (P value < .05, variable importance in projection [VIP] values >1, representing a 1.5-fold change). Kyoto Encyclopedia of Genes and Genome enrichment analysis was performed through MetaboAnalyst software (http://www.metaboanalyst.ca/). Pearson’s correlation analysis was applied to assess the correlation between lipids expression and irradiated doses and values of P < .05 were considered statistically significant. Student t tests, hierarchical clustering analysis, PCA, PLS-DA plots, and Pearson correlation analysis were performed using OmicShare (http://www.omicshare.com/tools).
Results
Selection of Radiation-Responsive Lipids
To obtain a full overview of lipids trend between different irradiation dose and postirradiation time, a hierarchical clustering analysis with the normalized abundance for a total of 42 lipids detected was performed (Supplementary Table 1). As observed in Figure 1, all PEs and 4 of 6 SMs were clustered together and the remaining lipids were assembled in another clustering. In order to observe the trend of lipidomic changes, PCA was used to cluster serum samples in each dose group at each time point (Supplementary Figure 1). For the determination of lipidomic signature, PLS-DA, a supervised method, was performed for all irradiation doses at 6, 24, and 72 hours postirradiation, respectively. Figure 2 shows 0 and 8 Gy were separated well no matter at 6 hours (R2Y = 0.952, Q2Y = 0.828) or at 24 hours (R2Y = 0.928, Q2Y = 0.479) and 72 hours (R2Y = 0.99, Q2Y = 0.963) postirradiation, while 2, 5.5, and 7 Gy were not scattered well from 0 Gy (Supplementary Figure 2). Therefore, we selected radiation-responsive lipids from the results of PLS-DA between 0 and 8 Gy, and the selection criteria of radiation-responsive lipids were based on a P value <.05, fold change greater than 1.5, and VIP values more than 1 (Supplementary Table 2). Consequently, 2 lipids, PC (18:2/18:2) and PC (18:0/18:2), were selected at 6 hours, 3 lipids, LysoPC 18:1, PC (18:0/20:4), and SM (D18:0/24:1), were chosen at 24 hours, and 5 lipids, LysoPC 18:1, PC (16:0/18:1), PC (18:0/18:2), PC (18:0/20:4), and LysoPC 18:2, were elected as radiation-responsive lipids. Coincidently, PC (18:0/18:2) represented responsive at 6 and 72 hours postirradiation, LysoPC 18:1 and PC (18:0/20:4) were all reactive at 24 and 72 hours postirradiation. And Figure 3 shows the dose–response relationship of the 7 radiation response lipids at 6, 24, and 72 hours after radiation.

Heatmap generated by hierarchical clustering analysis comparing 42 lipids in mouse serum after exposure to 0, 2, 5.5, 7, and 8 Gy at 6, 24, and 72 hours postradiation.
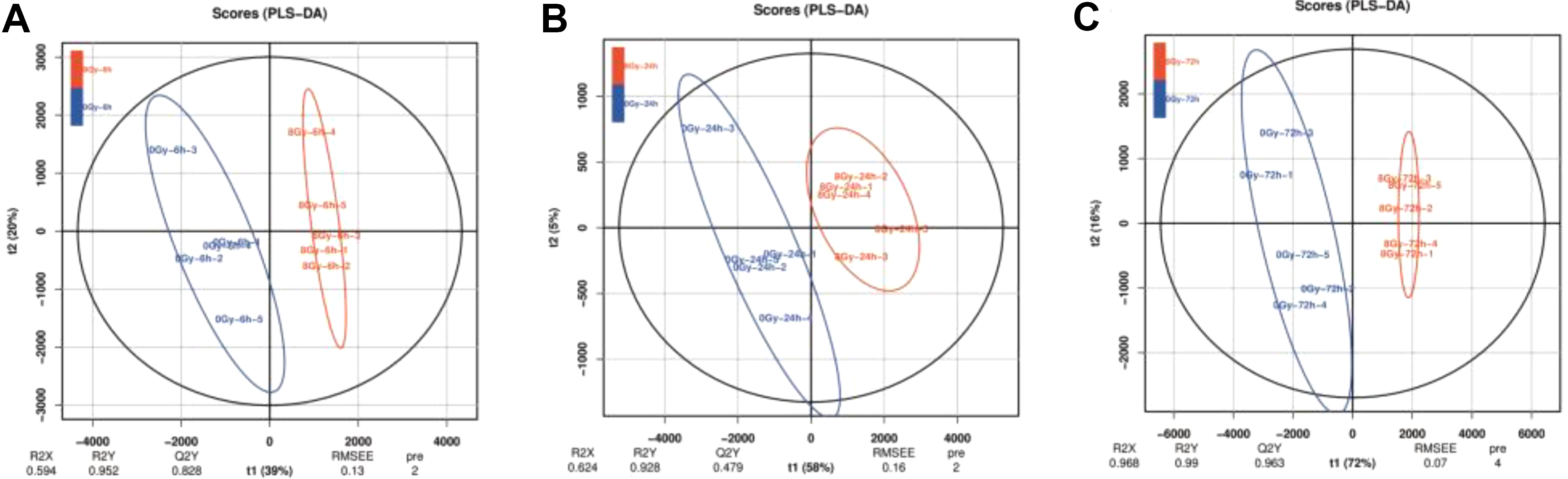
PLS-DA score plots of serum samples from 0 and 8 Gy groups represented by the blue line and red line, respectively, at (A) 6, (B) 24, and (C) 72 hours postradiation. One data point stands for 1 mouse. n = 5 per group. The corresponding R2X, R2Y, and Q2Y values are shown in (A, B, and C), respectively. PLS-DA indicates partial least squares discriminant analysis.

Dose–response relationship of (A) PC (18:2/18:2) and PC (18:0/18:2) at 6 hours after radiation; (B) SM (D18:0/24:1), PC (18:0/20:4), and LysoPC 18:1 at 24 hours after radiation; (C) LysoPC 18:1, PC (16:0/18:1), PC (18:0/18:2), LysoPC 18:2, and PC (18:0/20:4) at 72 hours after radiation. Error bars indicate ± 1 SD for each radiation exposure group. n = 5 per group. *P < .05 in the irradiated mice compared with mice in the 0 Gy group.
Dose–Response Relationship of Radiation-Responsive Lipids
Two lipids significantly changed in the 8 Gy group at 6 hours postirradiation. Their normalized abundance in the 2, 5.5, and 7 Gy group is shown in Figure 3A, and a 2-sample Student t test (2 tailed) was performed with 0 Gy group. PC (18:2/18:2) and PC (18:0/18:2) decreased as the dose of radiation increased. PC (18:2/18:2) began to decrease in the 2 Gy group and significantly decreased in the 5.5, 7, and 8 Gy group. PC (18:0/18:2) slightly decreased in the 2 and 7 Gy group and resulted in a significant reduction at 5.5 and 8 Gy.
Three lipids significantly changed in the 8 Gy group at 24 hours postirradiation. As shown in Figure 3B, SM (D18:0/24:1) and PC (18:0/20:4) increased while LysoPC 18:1 decreased as the dose of radiation increased. SM (D18:0/24:1) showed an increasing trend from 5.5 Gy and significantly increased in the 8 Gy group. Similar trend was observed in PC (18:0/20:4), which increased after 5.5 Gy irradiation and significantly increased in the 7 and 8 Gy group. LysoPC 18:1 start to decrease in the 2 Gy group and significantly decreased in the 7 and 8 Gy group.
Five lipids significantly changed in the 8 Gy group at 72 hours postirradiation. As shown in Figure 3C, only PC (18:0/20:4) resulted in an increased trend, while LysoPC 18:1, PC (16:0/18:1), PC (18:0/18:2), and LysoPC 18:2 decreased with the increase in the dose. Firstly, LysoPC 18:1 and LysoPC 18:2 smoothly and significantly decreased in all dose groups. Then, PC (16:0/18:1) and PC (18:0/18:2) showed a statistically significant response from 5 to 8 Gy. Finally, PC (18:0/20:4) significantly increased only in the 7 and 8 Gy group.
Kyoto Encyclopedia of Genes and Genome Enrichment Analysis of the Radiation-Responsive Lipids
To recognize the pathway and network changes in which the 7 radiation-responsive lipids are involved, KEGG enrichment analysis was performed by the MetaAnalyst software. A total of 5 pathways were enriched, including “Glycerophospholipid metabolism,” “Linoleic acid metabolism,” “alpha-Linolenic acid metabolism,” “Sphingolipid metabolism,” and “Arachidonic acid metabolism,” and the top 4 were significant (Figure 4, Supplementary Table 3). In addition, “Glycerophospholipid metabolism” was the most remarkable pathway, as a total of 30 compounds were in the pathway and 2 hits matched with. In the other 3 significant pathways, only 1 hit corresponded.

KEGG pathway enrichment analysis of the 7 radiation-responsive lipids. Count: number of compounds related to the enriched KEGG pathway. The color of the dot indicates the P value. KEGG indicates Kyoto Encyclopedia of Genes and Genome.
Selection of Potential Lipid Biomarkers as Radiation Biodosimeters
To select potential lipid biomarkers as radiation biodosimeters, the correlation between lipid expression and irradiated dose at all time points was assessed by Pearson correlation analysis. Table 1 shows all correlation coefficients and P values of the 7 radiation-responsive lipids. The expression of PC (18:2/18:2) showed a good linear correlation with the irradiated dose at 6 hours after exposure. PC (18:0/18:2) was correlated with the irradiated dose both at 6 and 72 hours postirradiation. Then, LysoPC 18:1 showed a good linear correlation with the irradiated dose both at 24 and 72 hours after exposure. PC (18:0/20:4) was responsive to the irradiation after 24 and 72 hours, but its increase was correlated with the dose only at 24 hours postirradiation. Finally, SM (D18:0/24:1) was correlated with the irradiated dose at 24 hours, while PC (16:0/18:1) and LysoPC 18:2 presented a good linear correlation only at 72 hours postirradiation.
Correlation Between Lipid Expression and Irradiated Doses at Different Time Points Postradiation.

Discussion
“An event that has led to significant consequences to people, the environment or the facility” is the definition of a nuclear and radiation accident by the International Atomic Energy Agency. Since the first nuclear reactors were created in 1954, more than 100 serious nuclear accidents have occurred since 2014. 17 Thus, the estimation of the radiation dose has always been placed as first in the medical management of these accidents. Biomarkers of radiation biodosimetry including lymphocyte depletion and analysis of chromosomal aberrations, especially the latter one, are widely used in the assessment of the radiation-induced changes. Nowadays, gene expression and protein level are promising biomarkers. 18 Furthermore, lipid perturbations after radiation exposure have been reported, although specific lipids considered as biomarkers of radiation have not been investigated much enough. 4,14 -16 Therefore, a lipidomic profile to evaluate biomarkers for radiation biodosimetry was performed in the current study. A change was detected in a total of 42 lipids in the mouse serum at 6, 24, and 72 hours after 0, 2, 5.5, 7, and 8 Gy radiation and were included in 5 lipid classes, such as SMs, PEs, PCs, LPEs, and LyPs. Then, PLS-DA, a linear classification model usually used to select discriminative features in the data and to classify the samples, 19 was performed between the 0 Gy group and each radiation dose group (2, 5.5, 7, and 8 Gy). Unsurprisingly, 8 Gy groups were the ones best separated from 0 Gy groups at 3 time points because 8 Gy was considered as the appropriate dose for the occurrence of radiation metabolism effect. 4 Moreover, 7 Gy groups were also separated well from 0 Gy groups at 24 and 72 hours postirradiation (Supplementary Figure 1), as 7 Gy was also high enough to be lethal and a reaction time of more than 24 hours was sufficient to obtain metabolic changes after exposure. Therefore, the selection criteria were defined with a P value <.05, fold change greater than 1.5, and VIP value more than 1 in 8 Gy groups. Consequently, 7 lipids, such as PC (18:2/18:2), PC (18:0/18:2), Lyso PC 18:1, PC (18:0/20:4), SM (D18:0/24:1), PC (16:0/18:1), and Lyso PC 18:2, were selected as radiation-responsive lipids for further exploration as biomarker for radiation biodosimetry.
PC is a component of biological membranes and its biosynthesis and degradation is considered necessary for cell cycle progression and its missing synthesis is a hallmark of cell apoptosis. 20 After 2, 4, 6, 7, or 10 Gy total body irradiation, PCs in serum of non-human primates (NHPs) generally declined in a dose-dependent trend. 4 A significant or slight increase in PCs is observed in the serum of NHPs exposed to 6.5 Gy γ-radiation. 16 In our study, PC (18:2/18:2), PC (18:0/18:2), and PC (16:0/18:1) decreased and PC (18:0/20:4) increased in a dose-dependent trend. It is well known that radiation can induce cell apoptosis. The destruction of phosphatidylcholines metabolism has been identified during apoptosis. 21 On the other hand, p53 is an important biological regulator of DNA damage-induced G1 arrest in cells after irradiation, and a p53 pathway would be stimulated by aberrant biological situation of deficient PC synthesis and degradation. 22 Therefore, this suggests that PC perturbation may be due to apoptosis caused by radiation and simultaneously playing a role in G1 arrest.
LyPs are membrane-derived signaling molecules that play various roles in a wide range of biological activities and diseases. In addition, serum LyPs level can be considered as biomarker for numerous disorders including myeloma, ovarian, and colorectal cancer. 23 -25 Moreover, after exposure to 6.5 Gy γ-radiation, Lyso PC 18:2 and Lyso PC 18:3 slightly decrease at 6 and 24 hours postirradiation in serum of NHPs, 16 and serum Lyso PCs level is significantly higher in NHPs exposed to 10 Gy than 0, 2, 4, 6, and 7 Gy. 4 G-protein-coupled receptors–interacting proteins are considered as playing crucial roles in repair mechanisms of DNA damages caused by radiation. 26 Furthermore, LyPs activities are mediated by G-protein–coupled receptors. 27 This aspect could be the reason of the declined trend of Lyso PC 18:1 and Lyso PC 18:2 by the irradiation dose in our study. SMs, converted from ceramide response to sphingomyelin synthase in the Golgi apparatus, can be stimulated by pro-inflammatory cytokines and oxidative stress, 28 and their change can be also the result of the radiation damage to the organism. This could be the explanation of the increase of SM (D18: 0/24:1) after 8 Gy exposure in our work.
Glycerophospholipid metabolism—a pathway is related to acute lymphoblastic leukemia, 28 and it is considered as the most significant pathway among the 7 radiation-responsive lipids. The biosynthesis of GPs depends on CDP-DAG pathway and Kennedy pathway, and the degradation is regulated by different phospholipases including phospholipase A1, phospholipase A2, and phospholipase B. 29 Because of its complexity and hidden aspects of the GP metabolism, the mechanism after the occurrence of radiation should be explored more in detail by further studies.
Finally, in our purpose of finding the potential biomarkers of radiation biodosimetry, Pearson correlation analysis was performed to evaluate the ability of the 7 lipids to estimate radiation doses. This is the first report attempting to apply lipids to estimate the radiation dose rather than only find out the biomarkers. Except for PC (18:0/20:4) that was not precise enough at 72 hours, the other lipids resulted in a satisfactory correlation coefficient, suggesting that these 7 lipids not only were radiation-responsive but also suitable as biomarkers to establish radiation doses within 3 days after exposure. In our subsequent study, longer time points beyond 7 days, such as 30 days will be considered in order to determine different biomarkers in different radiation stages. Secondly, cytokines regulated by radiation including interleukin (IL)-1β, IL-6, tumor necrosis factor-α, granulocyte-macrophage colony stimulating factor (GM-CSF), and granulocyte colony-stimulating factor (G-CSF) will be detected to further to explore the mechanism of lipid regulation after exposure.
Conclusions
In this study, a lipidomic profile was performed to select potential lipid biomarkers for radiation biodosimetry in mice serum at 6, 24, and 72 hours postradiation with 0, 2, 5.5, 7, and 8 Gy. Seven lipids including PC (18:2/18:2), PC (18:0/18:2), Lyso PC 18:1, PC (18:0/20:4), SM (D18:0/24:1), PC (16:0/18:1), and Lyso PC 18:2 were detected as modified and they also resulted appropriately for the estimation of the radiation dose. Further studies on nonhuman primates as a radiation model should be performed to better understand the application value of the results; then the combination with longer time points and more reactive cytokines could play a significant role in biodosimetry.
Supplemental Material
Supplementary_Figure_1 - Lipidomic Profiling for Serum Biomarkers in Mice Exposed to Ionizing Radiation
Supplementary_Figure_1 for Lipidomic Profiling for Serum Biomarkers in Mice Exposed to Ionizing Radiation by Jinfeng Huang, Qi Wang, Zhenhua Qi, Shixiang Zhou, Meijuan Zhou and Zhidong Wang in Dose-Response
Supplemental Material
Supplementary_Figure_2 - Lipidomic Profiling for Serum Biomarkers in Mice Exposed to Ionizing Radiation
Supplementary_Figure_2 for Lipidomic Profiling for Serum Biomarkers in Mice Exposed to Ionizing Radiation by Jinfeng Huang, Qi Wang, Zhenhua Qi, Shixiang Zhou, Meijuan Zhou and Zhidong Wang in Dose-Response
Supplemental Material
supplementary_Table_1 - Lipidomic Profiling for Serum Biomarkers in Mice Exposed to Ionizing Radiation
supplementary_Table_1 for Lipidomic Profiling for Serum Biomarkers in Mice Exposed to Ionizing Radiation by Jinfeng Huang, Qi Wang, Zhenhua Qi, Shixiang Zhou, Meijuan Zhou and Zhidong Wang in Dose-Response
Supplemental Material
supplementary_Table_2 - Lipidomic Profiling for Serum Biomarkers in Mice Exposed to Ionizing Radiation
supplementary_Table_2 for Lipidomic Profiling for Serum Biomarkers in Mice Exposed to Ionizing Radiation by Jinfeng Huang, Qi Wang, Zhenhua Qi, Shixiang Zhou, Meijuan Zhou and Zhidong Wang in Dose-Response
Supplemental Material
supplementary_Table_3 - Lipidomic Profiling for Serum Biomarkers in Mice Exposed to Ionizing Radiation
supplementary_Table_3 for Lipidomic Profiling for Serum Biomarkers in Mice Exposed to Ionizing Radiation by Jinfeng Huang, Qi Wang, Zhenhua Qi, Shixiang Zhou, Meijuan Zhou and Zhidong Wang in Dose-Response
Footnotes
Authors’ Note
J.H. and Q.W. contributed equally to this study.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by Logistics research key projects: BWS18J008.
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.