
Abstract
The phenomenon by which irradiated organisms including cells in vitro communicate with unirradiated neighbors is well established in biology as the radiation-induced bystander effect (RIBE). Generally, the purpose of this communication is thought to be protective and adaptive, reflecting a highly conserved evolutionary mechanism enabling rapid adjustment to stressors in the environment. Stressors known to induce the effect were recently shown to include chemicals and even pathological agents. The mechanism is unknown but our group has evidence that physical signals such as biophotons acting on cellular photoreceptors may be implicated. This raises the question of whether quantum biological processes may occur as have been demonstrated in plant photosynthesis. To test this hypothesis, we decided to see whether any form of entanglement was operational in the system. Fish from 2 completely separate locations were allowed to meet for 2 hours either before or after which fish from 1 location only (group A fish) were irradiated. The results confirm RIBE signal production in both skin and gill of fish, meeting both before and after irradiation of group A fish. The proteomic analysis revealed that direct irradiation resulted in pro-tumorigenic proteomic responses in rainbow trout. However, communication from these irradiated fish, both before and after they had been exposed to a 0.5 Gy X-ray dose, resulted in largely beneficial proteomic responses in completely nonirradiated trout. The results suggest that some form of anticipation of a stressor may occur leading to a preconditioning effect or temporally displaced awareness after the fish become entangled.
Introduction
Background to Bystander Effect
The bystander effect (BE) in its broadest sense refers to an effect detected in a cell, tissue, or organism, which was not itself exposed to a stressor but which was in receipt of signals from the exposed entity. Bystander effect has been seen across plant and animal kingdoms and in response to radiation, chemical, and pathological stressors. 1 They are thought, therefore, to result from a very ancient mechanism of communication that has been highly conserved through evolution and time. 2 –4 Most BE research has been conducted using electromagnetic radiation because it is nonresidual and simplifies the methods needed to demonstrate true communication and induction of responses that cannot be attributed to the residue of the original stressor. However, there are convincing reports that in fish at least, heavy metals and pathogens can induce BE. 5 –8
Background to Physical Signal
Although the existence of BE is widely accepted, the mechanisms are still not well worked out. It is known that the physical conditions of the experiment can influence outcome as can the dose of inducing agent, the underlying genetics, and the time at which effects are measured. 9 –12 The signal/response mechanisms are unclear with many candidate signals and response pathways. Most researchers agree that reactive oxygen and/or nitrogen species are involved and that p53, cytokines, and stress response pathways such as Mitogen activated protein kinase (MAPK) are important. 13 –15 Ion channels in both outer cell membranes and mitochondrial membranes appear to be involved. 16 –19 More recently, a role of UV resulting from both excitation decay and exosomes has been identified, 20 –23 and these are thought to contain miRNAs, although exosome profiling is at a very early stage. 24,25
Although most of BE research has been conducted using medium transfer, microbeam-targeted irradiations, or coculture techniques, 26 –29 there have been reports mainly from the authors’ or close collaborators’ laboratories of a physical component to the bystander mechanism. This was suspected early on by Mosse et al, 30 who showed that melanin could prevent BE. Experiments using multiwells suggested that if cells were added to wells of a dish after irradiation of cells in other wells, the newly added cells showed BE even though there was no medium transfer. 31 Other in vitro evidence comes from experiments using acupuncture or Reiki techniques to alter radiobiological outcomes 32 —both techniques are thought to work by balancing electromagnetic fields. Experiments using a Faraday cage to house cells during irradiation also support the role of EM fields in determining BEs. 31,33 Convincing evidence was also produced in vivo when irradiated fish were put in a closed aquarium within another aquarium containing unirradiated fish. 34,35 The bystander fish received signals from the irradiated fish even when the inner aquarium was wrapped in aluminum foil to prevent light signal transfer. In those experiments, we speculated that sound or vibrational signals might be involved and the attenuation calculations did not exclude this possibility. More recently, we have become aware of vibration-gated ion channels in hearing organs, which further supports the idea of a physical signal. 36,37 Most recent data from our laboratory using cell cultures has provided strong evidence that one of the physical signals may be Ultraviolet light A (UVA). 38 –40 Photons in the UVA range are emitted from cells treated with tritium and these photons can induce BEs in unirradiated cells. The fact that physical signals that are components of the electromagnetic spectrum can act as communicators of information between biological systems led us to consider whether quantum mechanical processes might mediate some of the signaling pathways involved in BEs.
Quantum Processes in Biology
In the last few years, there have been many studies of possible quantum effects in biology. In particular, the phenomenon of quantum entanglement is seen as a process by which communication in biology and the performance of complex molecular tasks could be speeded up and efficiently achieved. 41 –43 Although some have presented alternative explanations not involving entanglement while accepting coherence (eg, in the study by Duan et al 44 ), the idea remains attractive and many theoretical and experimental articles are now published in the area. An early article in the field suggested that quantum processes were operating in chloroplasts and were enhancing the efficiency of photosynthesis. 45 More recently, the process has been suggested as a mechanism for highly efficient DNA repair synchronisation 46 for explaining magnetic aspects of migration in insects, birds, and fish among other species 47 and for some emergent properties in the neural system in humans and other species. 48 Entanglement between experimenter and experiment has even been suggested as an explanation for the “memory of water emergent signal.” 49 The idea that quantum entanglement might play a role in bystander signal communication is attractive on many levels. Communication via photons resulting presumably from excitation decay in organic molecules and the involvement of voltage and vibration-gated ion channels in the mechanism together with the reported involvement of entanglement in photosynthesis and in DNA repair make it at least worth testing whether it could be involved in this form of communication.
The experiments reported here proposed to test this hypothesis built on our previous experiments where irradiated fish were let to swim with unirradiated fish for a few hours before all groups and sham controls were examined for production of bystander signals. For the entanglement experiments, fish from 2 distinct populations that had never met before, swam together for 2 hours. They were then separated, 1 group was irradiated and the others were not. They did not meet again and were subsequently examined along with controls (positive and negative) for bystander signal production.
Methods
The experimental design is presented in Figure 1. Two separately sourced fish populations were designated as group A, which were actually irradiated, or group B which were bystanders. Essentially, there were 9 experimental groups—fish irradiated before meeting and their bystanders (positive controls), fish irradiated after meeting and their bystanders, fish which met but neither population were irradiated, sham-irradiated population A fish (handling control), and absolute controls from each population which were never handled. Within each experiment, there were 2 biological repeats of 4 fish per group (total 8 fish per treatment) and each experiment was repeated twice with trout and twice with zebra fish.

Scheme of the experiments. A and B refer to the 2 fish populations.
Fish Husbandry
Zebra fish
Zebra fish (550-650 mg), of mixed sex, were purchased from 2 different local fish retailers—a local pet store (PetSmart, Hamilton, Ontario, Canada) and Aqua-Life International, Winnipeg, Manitoba, that is, certified commercial outlets. All fish were monitored for loss of appetite, lethargy, lesions signs of physical damage, pigment or scale loss as well as distension of the abdomen, and exophthalmia (as is the standard practice for all fish held in the facility). However, no antibiotics or other specific treatments were used since (1) all groups of fish (zebra fish and rainbow trout) had to be treated identically and (2) the nature of this experiment demanded that additional factors, apart from the X-ray exposure, were (as much as possible) eliminated.
Rainbow Trout
The rainbow trout were obtained from Humber Springs Hatchery, Orangeville, Ontario, Canada, and Rainbow Springs Hatchery, Thamesford, Ontario, Canada, that is, certified producers. The fish were monitored for the conditions/symptoms listed above.
Both species of fish were acclimated in separate rooms over 2 weeks in 40 L of dechlorinated Hamilton city water in a glass aquarium. Water temperature was set to 28°C (Top Fin 50 W aquarium heater) and was mechanically, chemically, and biologically filtered through an external power filter (AquaClear), before being returned to the aquarium via a “waterfall” inflow. This, together with aeration from a diaphragm type air pump (Elite model 800) and air-stone diffuser (Top Fin), provided adequate aeration. Feeding was twice daily with a commercially available tropical fish flake diet (Nutrafin). All fish husbandry supplies were obtained from PetSmart, Burlington, Ontario, Canada. Full details on husbandry can be found in the study by Mothersill et al 50 All experiments were covered by an Animal Utilisation Protocol in place at McMaster University.
Irradiation
Fish meeting after irradiation of the “A” group
A single 0.5 Gy X-ray dose was administered using a portable X-ray machine (Cabinet X-ray system; Faxitron X-ray Corporation, Wheeling, Illinois), delivering 100 mGy per minute, exactly as previously described. 50 This acute exposure took 5 minutes, that is, the acute X-ray exposure dose rate was 0.1 Gy min−1. Once X-rayed, the zebra fish were placed in one side of a container divided by a mesh screen. After a 2-hour interval (the time span was chosen based on previous investigations to allow bystander signals from the fish to build up in water), 4 nonirradiated (bystander) zebra fish were placed on the other side of the mesh-divided container. Two hours later, both the irradiated and nonirradiated fish were sacrificed by concussion followed by spinal transection, using protocols approved by McMaster University’s Animal Care Committee. Under aseptic conditions, the caudal fins were immediately removed (at the peduncle) and placed in ice cold Roswell Park Memorial Institute (RPMI)-1640 culture medium (Gibco Biocult; VWR, Burlington, Ontario, Canada).
Processing of these tissue samples, primary explant culture, assessment of the direct effects of radiation and the BE on colony-forming ability, and apoptosis of the human papillomavirus strain G (HPV-G)–transfected human keratinocytes reporter cells have all been previously described and validated, 50,51 and brief descriptions only are included below.
To evaluate the handling and confinement stress associated with the X-ray procedure and transfer of fish to the holding and bystander containers, a full series of sham controls were also included. Sham X-rayed fish were placed in the X-ray cabinet for 5 minutes (the time required to deliver a 0.5 Gy dose) but without the X-ray being switched on. These sham X-rayed zebra fish were then used to generate sham bystander fish (as described above). Sham tissue samples were collected and processed in the same way as were tissue samples from completely untreated fish, that is, taken directly from the holding stock.
Fish meeting before irradiation of the “A” group
The procedure for this group was identical except that the 2-hour period of swimming on either side of the mesh grid took place immediately before irradiation of the “A’ group. After irradiation, the “A” and “B” groups were placed in separate containers in separate rooms and did not meet again. They were sacrificed 2 hours after irradiation.
Clonogenic Assay procedure
Explant culture
Explants of zebra fish caudal fins were established as described previously. 50 Briefly, tissues were dissected aseptically, and 3 equal-sized pieces (approximately 1-2 mm2) were plated as single explants in the center of 25 cm2 growth area, 50 mL volume flasks (Falcon; VWR) in 2 mL growth medium. Flasks were left undisturbed for 48 hours at 19°C in a refrigerated incubator. All tissue was handled according to biosafety guidelines at McMaster University.
Reporter cell culture
The HPV-G cell line was originally given to us by Dr J DiPaolo, National Institutes of Health, Bethesda, Maryland. It was obtained at an early passage and expanded and frozen in our laboratory. The cell line has a well-characterized and stable bystander response, showing a reduction in cloning efficiency of ∼40% over a wide range of radiation doses and well-characterized calcium fluxes and mitochondrial effects.
52,53
This makes it ideal as a reporter system. All cell culture procedures were performed in a class II biosafety cabinet. The cells were grown in RPMI medium containing 60 mL prescreened fetal bovine serum, 5 mL penicillin–streptomycin, 5 ml
Clonogenic assay technique and bystander protocol
Cell cultures that were 85% to 90% confluent and that had received a medium change the previous day were selected. Cells were removed from the flasks using 0.25% wt/vol trypsin/1 mM EDTA solution (1:1) obtained from VWR. When the cells had detached, they were resuspended in medium, and an aliquot was counted using a Z2 Coulter particle count and size analyzer (Beckman Coulter Electronics, Mississauga, Ontario, Canada). Appropriate cell numbers (∼500) were plated for the recipient or bystander flasks to optimize the ratio of signal molecules to cell number. Forty-eight hours after set up of the explants, medium was poured off from the flasks containing the explants. The medium was filtered through a 0.22-mm filter to remove any debris from the explant culture medium and then added to the cells in the reporter flasks from which the original medium had been removed. Ten days later, colonies of reporter cells were stained with Carbol Fuchsin (Ziehl-Neelsen, Sigma-Aldrich, St. Louis, Missouri), and colonies were counted to determine reporter cell survival.
Calcium Assay
Fura-2 measurements to determine intracellular free calcium in HPV-G cells
The HPV-G cells were seeded in 30-mm glass-bottomed culture vessels (MatTek, obtained from VWR Burlington Ontario) at a density of approximately 500 000 cells and incubated at 37°C and 5% CO2 for 18 to 24 hours prior to measurement to achieve 50% confluence. Cells were washed 3 times with buffer (130 mM NaCl, 5 mM KCl, 1 mM Na2HPO4, 1 mM CaCl2, 1 mM MgCl2, and 25 mM Hepes [pH 7.4]) followed by incubation with 1 mL of 4.17 μM fura-2-acetoxymethyl (Fura-2-AM) ester (all chemicals obtained from Sigma-Aldrich) at 37°C for 30 minutes. Cells were washed 3 times with buffer to remove residual Fura-2 with 300 μL of fresh buffer remaining in the dish for imaging. An Olympus 1X81 microscope was used with a ×40 oil objective and Fura filter cube with 510 nm emission. Five cells were selected from the field of view and Fura-2 was excited at 380 and 340 nm. The ratio images were recorded every 4 seconds for 5 minutes with addition of 100 μL of irradiated cell conditioned medium or control media after a stable baseline was reached approaching 30 seconds. All measurements were conducted in the dark at room temperature.
Proteomics
Proteomics was only attempted on the rainbow trout due to the costs involved and the difficulty of harvesting and processing zebra fish tissue.
Two-dimensional gel electrophoresis, proteome analysis, in-gel digestion mass spectrometry, and protein identification
A weighed sample of gill filament (approximately 15-20 mg) from each fish was homogenized in 300 µL of ice-cold 2-dimensional electrophoresis (2-DE) lysis buffer (for composition refer to Smith et al 54 ). The lysates were clarified by centrifugation (10 000g for 5 minutes at 4°C), de-salted using a commercially available de-salting kit (Thermo Fisher Scientific, Ontario, Canada) and the total protein content measured, using the Bradford protein assay (BioRad Laboratories Inc, Mississauga, Ontario, Canada). A 40-µg protein were then subjected to 2-DE using the Protean 2-DE system apparatus, gels, buffers, and stain (BioRad). Following rehydration, pH 4 to 7 immobilized pH gradient strips were loaded with the protein lysate. Isoelectric focusing used a 3-step ramped voltage 55 and second dimension electrophoresis used Laemmli chemistry gels and tris/glycine SDS buffer. 55 The gels were then fixed with water, methanol, and acetic acid (Calendon Laboratories Ltd, Ontario, Canada) and stained with SYPRO-ruby stain. After de-staining, the gels were washed in 10% ethanol and the images captured using a Fluor-S Max gel imager; wavelength = 660 nm.
Gel image analysis was carried out using the Phoretix 2-D software (Progenesis PG200, Phoretix International, United Kingdom) with protein expression being quantitatively expressed as “normalized spot volume,” a parameter which combines the spot size and intensity. Protein spots were selected for excision and protein identification based on the consistency of protein expression and a statistical comparison of normalized spot volumes. Specifically, (1) the spot had to be present on all gels and (2) show a statistical difference to the same spot from the untreated control fish. 5,54 –59
Selected protein spots were excised from the gel and prepared and digested for analysis by liquid chromatography mass spectrometry (LCMS), as previously described. 55 Analysis was completed using an Orbitrap Velos Pro (Thermo Fisher Scientific, Essen, Germany) mass spectrometer coupled to a nanoelectrospray ionisation source (Thermo Fisher Scientific), with 0.1% formic acid as effluent A and HPLC grade acetonitrile as effluent B, as previously described. 5
It is important to stress this analysis was conducted as a “blind study.” Gill samples from untreated trout, bystander trout, which swam with irradiated fish after they had been irradiated, and trout which swam with irradiated fish prior to irradiation, were all given random number sample codes. All proteomic analysis was then performed without knowing the origin of tissue samples. Only when the gel images had all been fully analyzed and the protein spots had been selected for identification were the fish treatments and the proteomic results combined.
Calculations and Statistical Analysis
All data are expressed as mean values (standard deviation [SD]). The effects of direct irradiation and BE HPV-G clonogenic data were analyzed by analysis of variance followed by least squares difference. The figures are therefore annotated to show which data are similar and which are statistically significant, that is, annotation with the same letter indicates similar data, whereas annotation with different letters indicates a statistically significant difference. All statistical analysis was carried out using Statistix analytical software (version 10) and in all cases a probability of <.05 was considered statistically significant.
For the proteomics section, LCMS peak area was used compare protein abundance. Peak area for each protein from the irradiated or bystander fish samples was divided by the mean peak from the untreated control fish to give a “fold change” 60 index of protein expression. All fold change data were then expressed graphically 61 as mean ± standard error of the mean. The effects of early life stage irradiation were then compared using Student t test (2-tailed). We did not restrict the selection of differentially expressed proteins to those which showed a minimal change (eg, >1.5-fold or <1.5-fold), 62 but we only report proteomic changes, which showed a statistically significant change (P < .05). However, given that the proteome is widely accepted as a highly fluid, dynamic entity and that proteomics can equally refer to the protein complement of a cell at a certain time or the complete set of proteins expressed during a cell life cycle, 63 the decision was made to adopt previous criteria of restricting our selection of proteins to those which were quantitatively detected by the analytical software (see above) in at least 50% of the samples. 64 The gills from a total of 11 control fish, and 5 irradiated and 5 bystander fish, from each irradiation group, were analyzed. Thus, the analysis described here was based on a minimum of n = 6 control and n = 3 irradiated or bystander fish.
Results
Clonogenic Assay Data
Figure 2A and B presents the clonogenic reporter assay data for rainbow trout fin (Figure 2A) and gill (Figure 2B) tissue explants from directly irradiated trout and their bystanders, which communicated either before or after the X-ray dose. The assay measures strength of the bystander signal as a reduction in the survival of the reporter cells, which respond to the signal by initiating a death response when exposed to bystander medium. Thus, an elevated readout compared to the controls means the signal is weaker than the controls, whereas a reduced readout means the signal is stronger than the controls. The strength or weakness of the signal in this assay does not imply any function of the signal in a system other than the reporter cells, it merely scores the signal as strong or weak relative to the control signal. The results show that a strong bystander signal is produced whether the group B fish meet before the radiation exposure to the group A fish or after that exposure. The data for the gill tissue are slightly more significant than the data for the fin, but in both tissues, it is clear that strong bystander signals are being produced under both experimental conditions. The data were normalized to the absolute controls. The sham X-ray group is not significantly different to the controls for either tissue. For the fish which met but were not X-rayed, the group A fish is not significantly different from the group A sham X-rayed group for either tissue, but the group B fish has a significantly elevated readout from the gill but a significantly reduced readout from the fin, although it is still significantly elevated compared with the group which communicated before irradiation.

Clonogenic survival of HPV-G cells receiving media from rainbow trout fin (A) and gill (B) explants from fish treated as shown. Letters above the bar indicate the degree of similarity or differences between the bars. Errors are SD for n = 8. HPV-G denotes human papillomavirus strain G; SD, standard deviation.
The data for zebra fish fin are shown in Figure 3. In a tropical species of considerably smaller size, these data again confirm that the bystander signaling mechanism is triggered whether the fish swim together only before irradiation of group A fish or only after irradiation of group A fish. Again the sham group A fish and group A fish which communicate with group B fish but are not irradiated are not significantly different from the controls. However, the group B fish have a significantly elevated readout, meaning the strength of the signal is weaker than in the absolute control group.

Clonogenic survival of HPV-G cells receiving media from zebra fish tail fin explants from fish treated as shown. Letters above the bar indicate the degree of similarity or differences between the bars. Errors are SD for n = 8. HPV-G denotes human papillomavirus strain G; SD, standard deviation.
Calcium Flux Data
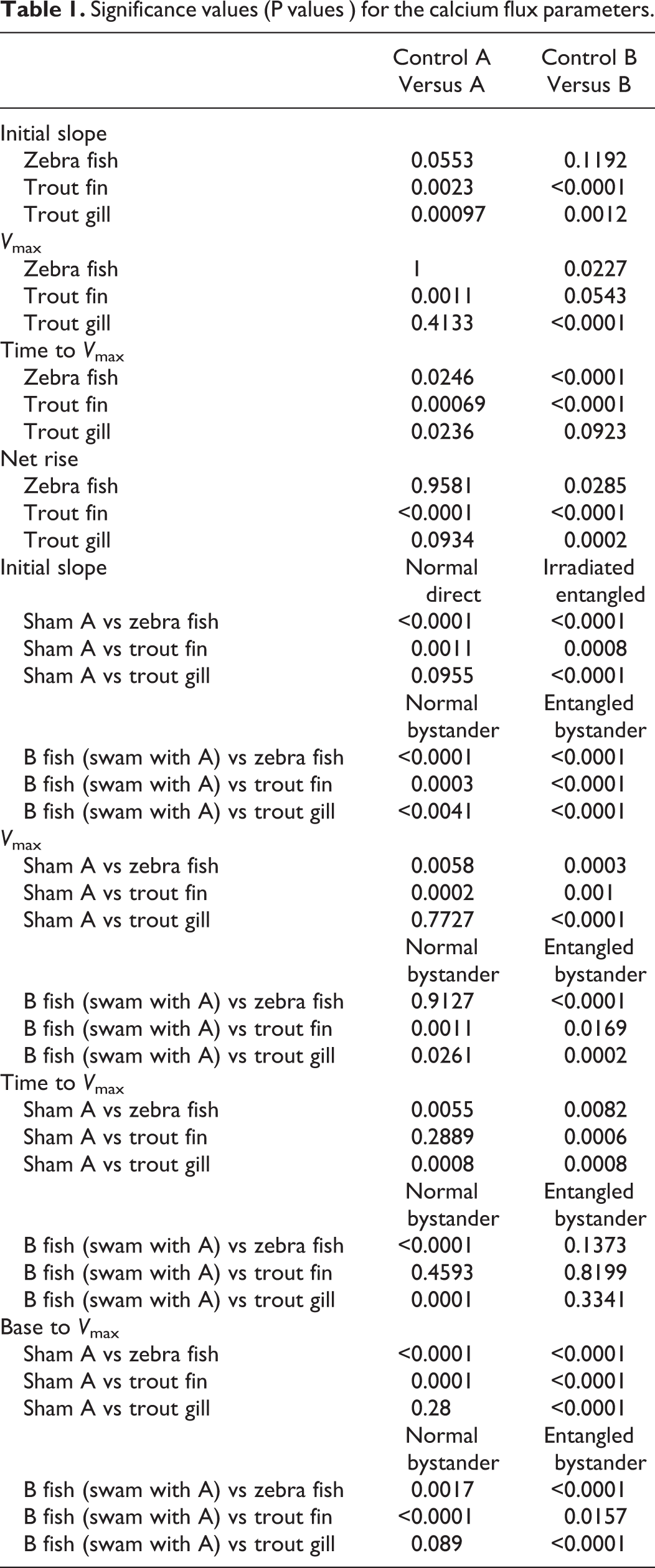
The first response of cells receiving irradiated bystander medium is to show a big influx of calcium through ion-gated calcium channels in the cell membrane 53,65 ; therefore, the calcium flux assay was done as a further check that bystander signals were being produced. The data for zebra fish tail fin, trout fin, and gill are shown on Figure 4A-D presented as percentage difference from the control for initial slope of the influx curve, the V max, maximum calcium influx (the difference between the baseline value and the maximum value and the time taken to reach this). The data were normalized to the appropriate control because of the significant differences in the control groups. Group A fish were all normalized to the sham irradiated A fish which were treated exactly the same as the A fish which swam with B fish but were also placed in X-ray machine although it was not turned on. The B group were normalized to the B fish swam with A fish where neither group was irradiated. The figures show expected influx results for the directly irradiated group A fish irrespective of when they met the group B fish. The positive control group B bystander fish, which met the irradiated fish after irradiation, also showed the expected normal calcium flux. However, in all experiments with both trout fin and gill and zebra fish tail fin, the group B fish which met before the group A fish were irradiated had calcium influxes that were significantly below those of the control fish. The P values for the calcium analysis are presented in Table 1.
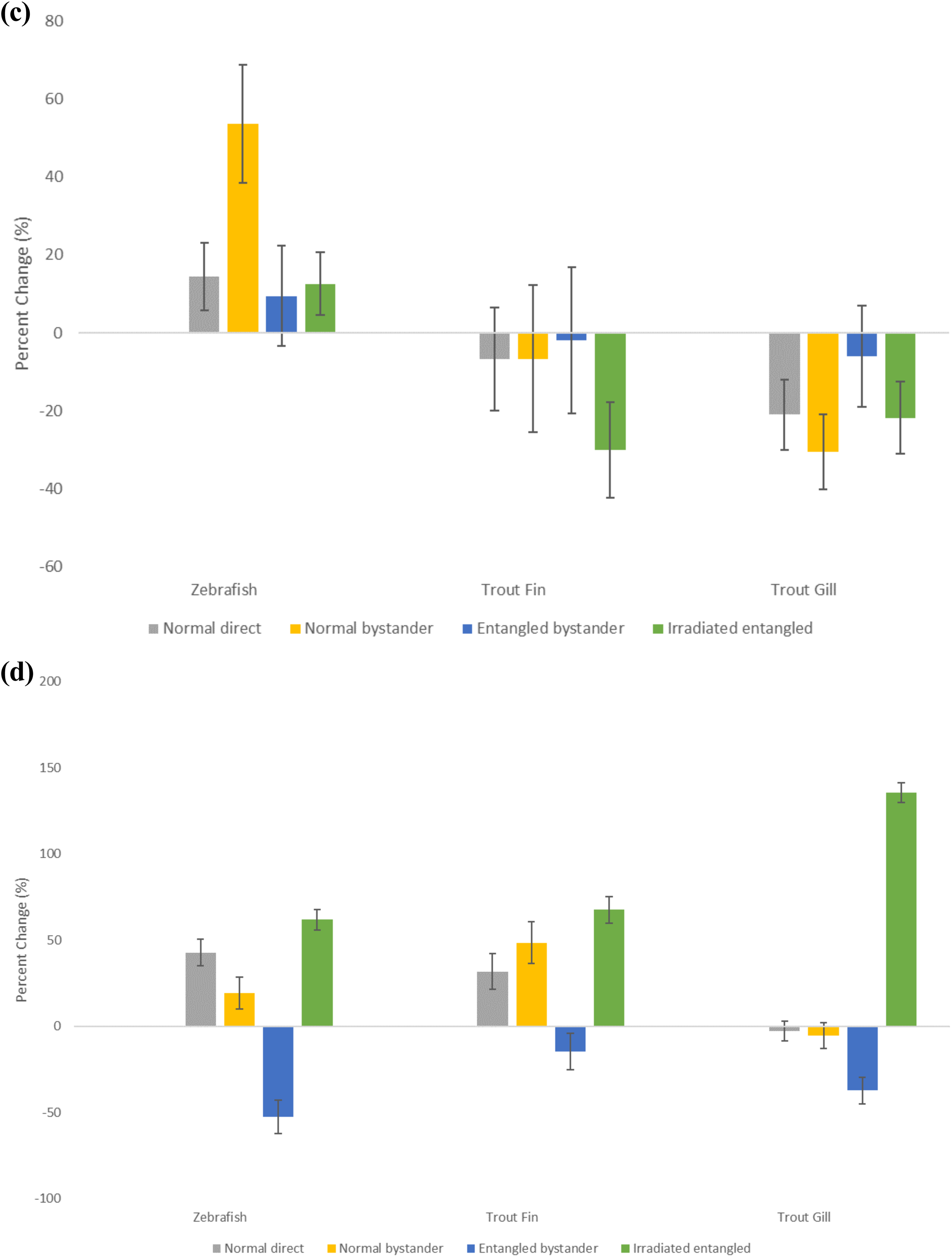
Calcium influx kinetics for zebra fish tail, rainbow trout fin, and rainbow trout gill expressed as percentage change from the relevant control. Kinetic parameters examined are the initial slope (A), V max (B), time to V max (C), and baseline to max (D). Errors are SD for n = 8. SD denotes standard deviation.
Significance values (P values ) for the calcium flux parameters.
Proteomics
Figure 5 illustrates a typical 2-D gel and Table 2 summarizes the changes in expression of the selected proteins. Two proteins, α-enolase and fructose-bisphosphate aldolase, were upregulated by radiation exposure only. Two proteins, 14-3-3 protein β subunit and ABC transporter G family member, were upregulated by the radiation-induced bystander effect (RIBE) only. Six protein spots showed an increase in normalized volume only in fish which swam with the irradiated fish prior to radiation exposure. However, 2 of these spots were identified as canalicular multispecific organic ion transporter/multidrug resistance associated protein 2 (cMOAT/MRP2). Therefore, 5 proteins were upregulated by quantum entanglement only; cMOAT/MRP2, bifunctional P450/NADPH-P450 reductase (hOR), glycine-tRNA ligase β subunit, gasdermin, and tubulin β-1 chain. Three protein spots showed a decrease in normalized volume only in fish which swam with the irradiated fish prior to radiation exposure; ubiquitin-40S ribosomal protein, tropomyosin α-1 chain/β chain, and tropomyosin α-3 chain.

Representative 2-D gel showing protein spots selected for identification. Nine canalicular multispecific organic ion transporter/multidrug resistance–associated protein 2, 14 intermediate filament protein, 18 α-enolase, 21 bifunctional P-450/NADPH-P450 reductase, 22 canalicular multispecific organic ion transporter/multidrug resistance–associated protein 2, 30 ubiquitin-40S ribosomal protein, 36 intermediate filament protein, 45 glucose-6-phosphate 1-dehydrogenase, 64 glycine-tRNA ligase β subunit, 69 14-3-3 protein β subunit, 74 gasdermin, 80 fructose bisphosphate aldolase, 82 Tubulin β-1 chain, 89 vacuolar membrane–associated protein IML1, 95 tropomyosin α-1 chain/β chain, 101 tropomyosin α-3 chain, 111 glucose-6-phosphate 1-dehydrogenase, 123 ABC transporter G family member. 2-D, 2-dimensional
Fold Changes in the 18 Selected Proteins Indicated in Figure 5.a
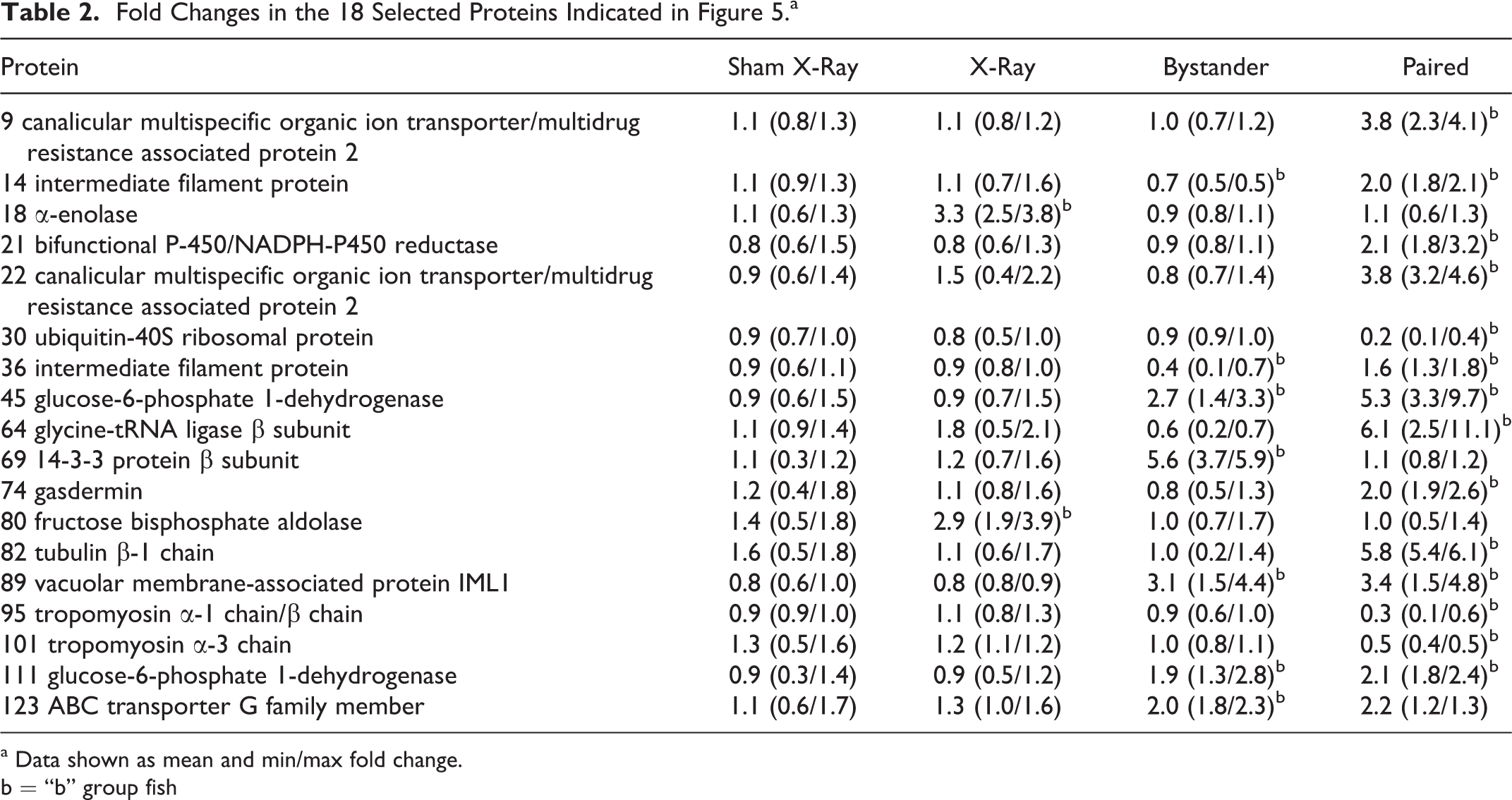
a Data shown as mean and min/max fold change.
b = “b” group fish
In addition, there were examples of proteins which showed a change in expression as a result of the RIBE and in fish which had swam with trout prior to irradiation, that is, in nonirradiated fish which had swam with irradiated trout irrespective of whether this was before or after irradiation. Three protein spots showed an increase in normalized spot volume in both the RIBE fish, which had swam with the irradiated fish after irradiation, and in the fish which swam with the irradiated fish prior to irradiation. However, 2 of these spots were identified as glucose-6-phosphate 1-dehydrogenase (G6PDH). Therefore, 2 proteins, G6PDH and vacuolar membrane-associated protein IML1, were upregulated by both the RIBE and in nonirradiated trout which swam with irradiated trout prior to irradiation. Two protein spots showed a decrease in normalized volume in RIBE fish but an increase in normalized volume in the fish, which swam with the irradiated fish prior to irradiation. However, both spots were identified as intermediate filament protein (IFP). Therefore, IFP showed the opposite responses to the RIBE and to swimming prior to irradiation.
Discussion
The results presented in this article involve 2 very different fish species separated by millions of years of evolution, 2 organs in 1 of the species and 3 different assay end points. All suggest that allowing 2 populations of fish to meet for the first time, then separate never to meet again, before 1 group in each experiment is irradiated, leads to effects in the nonirradiated bystander fish. We have previously published extensively that postirradiation meeting causes effects in nonirradiated fish, rats, and mice, 50,51,66,67 and other groups also have shown these organism level effects in frogs, mice, and water fleas, 68 –70 but this is the first report of preirradiation meeting, influencing outcome. At first sight, the results may seem beyond strange but the robust and well-tested assays require some consideration and explanation.
Clonogenic Data
The results for clonogenic reporter assay used in our traditional bystander experiments show that it does not matter whether the fish met before or after 1 group was X-rayed; the assay “reports” the presence of bystander signals in both cases. It should be noted that while the qlonogenic assay is based on reduced (or increased) clonogenic survival relative to the control, this does not mean the BE has adverse or protective effects, merely that the cell line used responds to the signal, if present, by showing a reduced or increased survival in the colony-forming assay.
Calcium Flux Assay
The calcium influx assay has been extensively used by our group to detect the presence of bystander signals in samples. 53,65,71 A rapid and large increase in intracellular calcium correlates with a strong bystander signal. Controls show a much smaller or absent calcium influx. The data presented in this article are interesting—the positive controls, that is, directly irradiated and bystander fish which swam together after the X-ray show as expected, high calcium influx in both groups. However, the groups which swam together and were separated before 1 group were X-rayed show a significantly reduced calcium influx measured by each parameter, which was below control levels in the group that were not X-rayed, whereas the group that were X-rayed show an increased influx compared with the X-rayed positive control fish. This suggests that the act of swimming with a previously unencountered cohort of fish changed the calcium response. This finding led us to examine the control data sets very carefully.
Analysis of Control Data
The fish groups sourced from different suppliers were initially designated as “group A” fish and “group B” fish. Group “A” were introduced into the tank after group B, although there were only minutes between the introductions. Group A were always irradiated, whereas group B were bystanders. Among the controls were a pairing of group A and group B fish where neither group were irradiated. For the zebra fish clonogenic assay experiments, the mean (SD) for group A is 115.3 (27.1) and for group B is 109.6 (8.4). This is convincingly nonsignificant (P = .58). For the trout experiments, the fin data for group A is 150.9 (16.7) and for group B is 166.8 (14.4). This is just nonsignificant (P = .06). For the trout experiments, the gill data for group A are 162.3 (17.1) and for group B are 154.6 (15.2). This is convincingly nonsignificant (P = .36). Based on the clonogenic data, therefore, the conclusion is that whatever explanation there is for the results where 1 group were actually irradiated, they cannot be explained as solely due to the introduction of a new fish cohort into a tank already containing fish.
Looking at the calcium assay, however, the data are not convincingly nonsignificant. “A” and “B” handling controls which never met produced similar calcium flux results, but the groups which met but where neither group were irradiated had small but statistically significant differences in calcium flux, and the groups which met where 1 group was irradiated after parting the fish showed a significant effect size difference in most of the parameters used to characterize the calcium influx, suggesting altered kinetics in both the “A” and “B” fish (a table of P values is presented (Table 1)). This suggests that the act of introducing new fish into the tank produced a response involving calcium signaling in both groups and this was significantly greater if 1 group was irradiated. The effect is small but does support the idea that introducing a group of fish into a tank containing fish they do not recognize can change biological responses. It is assumed, therefore, that the results presented are partly but not solely due to the control fish meeting, but the effect is greatly increased by the act of irradiating the group A fish either before or after the fish met.
Proteomic responses to direct irradiation
This is the second time a proteomic analysis has been made on the gills of rainbow trout subjected to the radiation exposure and RIBE-induction protocol described here. Previously, the use of the Investigator 2-DE system (Genomic Solutions, Ontario Canada) identified the upregulation of annexin II, a protein strongly associated with growth in irradiated fish, and the upregulation of potentially beneficial hemopexin-like protein, ρ-GDP dissociation inhibitor, and pyruvate dehydrogenase in bystander fish. 56
Differences in gel format, particularly isoelectric running conditions, are known to result in variations in protein spot pattern and, therefore, resolve different aspects of the proteome (eg, in the study by Duncan and Hershey 72 ). Despite this, the use of the Protean 2-DE system (BioRad), in the present investigation, provides additional evidence for alterations in energy metabolism toward growth stimulation or carcinogenesis, by direct exposure to 0.5Gy X-ray dose. 56 Specifically, the upregulation of both α-enolase and fructose bisphosphate aldolase, in the irradiated trout, has been associated with cancer or cancer progression in mammalian systems. α-Enolase is overexpressed in colorectal cancer 73 and is indicative of a poor prognosis of cholangiocarcinoma. 74 Additionally, antibodies against α-enolase have been found following lung small cell carcinoma. 75 Similarly, the downregulation of fructose bisphosphate aldolase (ie, the opposite response of the present investigation) has been associated with tumor suppression 76 and reduced proliferation of the mouse squamous carcinoma cell line, KLN-295. 77
Proteomic responses to the RIBE
The present investigation also provided additional evidence to support the potentially beneficial nature of the RIBE, 56 particularly with respect to cancer suppression. The 14-3-3 expression is an abundant protein involved in metabolism, protein sequestration and trafficking, cell cycle regulation and apoptosis, and also cancer suppression. 78 Thus, altering 14-3-3 expression may offer options for cancer treatment. 78,79 Most importantly, 14-3-3 expression is downregulated in tumors, 80 that is, the opposite of the upregulation seen in the present investigation. The RIBE downregulation of IFP was also the opposite response to that seen in cancers. For example, in small cell lung cancer, nestin (a class IV IFP) assists malignancy. 81 Downregulation would, therefore, suggest an antitumorigenic proteomic response and a reduction in IFP does reduce cancer invasion 81 and reduced cellular motility in astrocytomas. 82 Consequently, targeting IFP is a potential therapeutic strategy for malignancies. 83
The association between ABC transporter G family protein (ABCG) and cancer is less defined and, in fact, sometimes contradictory. For example, ABCG2 exhibits a range of expression in cancers and even in cell lines derived from the same tissue 84 although osteosarcoma cell lines do show higher levels of ABCG2 messenger RNA than osteoblasts. 85 Also, although ABCG2 plays no role in multidrug resistance of ovarian cancer, 86 excretion of tyrosine kinase, by ABCG2, may result in cancer resistance. 87 However, ABC transporters excrete toxic metabolites 87 –89 and protect against external pollutants. 90,91 In addition ABC10 protects against oxidative stress. 92 Radiation is known to cause oxidative stress, 93 and reactive oxygen species (ROS) production is thought to induce the BE (eg, in the study by Riganti 94 ). In terms of ROS protection, the upregulation of ABCG, therefore, augments the RIBE upregulation of hemopexin-like protein, previously seen in rainbow trout, 56 and mirrors the upregulation warm temperature acclimation-related 65-kDa protein (Wap65) in medaka (Oryzias latipes). 57
The relationship between G6PDH and cancer is similarly contradictory. Glucose-6-phosphate 1-dehydrogenase can be considered a putative oncogene but, while cancer cell proliferation is associated with higher G6PDH activity, which would suggest the RIBE-induced increase as being pro-tumorigenic, G6PDH overexpression is associated with a reduction in DNA double-stranded break repair. 94 This equally suggests upregulation of G6PDH has a restorative role for DNA damage. Glucose-6-phosphate 1-dehydrogenase activity is also directly related to oxidative stress. 95 The RIBE-induced upregulation, therefore, provides additional evidence for the involvement of ROS in the BE. Vacuolar membrane-associated protein IML1 is a protein which controls cellular growth and proliferation, 96 in particular by the regulation of autophagy, that is, organelle recycling or degradation. 97 Thus, it is easy to appreciate how, in terms of increasing repair capability, IML1 upregulation (such as G6PDH) could be beneficial to cells subjected to or at risk of oxidative damage.
Proteomic responses in fish which swam with the irradiated fish prior to irradiation
The fact that G6PDH and vacuolar membrane-associated protein IML1 were also induced, in nonirradiated fish, by the trout which were designated to be irradiated prior to irradiation as well as after irradiation, by the RIBE, suggests some degree of commonality in pre- and postirradiation meeting of the fish. However, the majority of responses in the trout which swam with the irradiated fish prior to irradiation were different to the responses resulting from the RIBE.
The upregulation of IFP in trout which swam with fish prior to irradiation was the opposite of the downregulation caused by the RIBE. The potentially beneficial nature of IFP downregulation, with respect to antitumorigenesis have been discussed. However IFP is a vital controller of cellular caspases during apoptosis 98 and a loss of IFP can alter cellular migration. 99 It is therefore quite possible that IFP plays a different role in preirradiation meeting than it does in the RIBE. We therefore propose this question requires additional investigation.
Aside from the above, the actual proteins which were upregulated or downregulated in trout, which had swum with fish prior to irradiation, were different to those which were upregulated or downregulated in bystander fish. Despite this, there were in fact some similarities in the physiological function. Bifunctional P450/NADPH-P450 reductase is a source of ROS, although other sources constitute a greater contribution to oxidative damage. 100 However the overexpression of hOR, the increased production of ROS and the resultant oxidative stress are both damaging to breast adenocarcinoma cells and enhance the treatment of anticancer treatment with 5-fluorouracil. 101 Thus, as was the case with, 14-3-3 expression, hOR upregulation suggests an antitumorigenic response. The downregulation of ubiquitin-40S ribosomal protein is another potentially antitumorigenic response. Ubiquitins are essential for protein homeostasis and apoptosis regulation. 102 Overexpression suppresses apoptosis and, consequently, has been observed in a number or tumors. 103 Similarly the removal (cleavage) of ubiquitin from other proteins enhances tumor necrosis factor–mediated apoptosis. 104 Apart from an antitumorigenic response, communication from the irradiated trout prior to irradiation also elicited proteome changes which were potentially protective to the integrity of the gill epithelium. Like ABCG2, cMOAT/MRP2 is important in xenobiotic detoxification and transports a number of substrates across epithelia. 105 Gasdermin is another protein which plays a vital role in epithelial function, particularly in relation to maintaining a barrier. 106 Gasdermin expression is also linked to cellular proliferation but without promoting tumorigenesis or maliganacy. 107 The freshwater fish gill forms a highly impermeable barrier against a hypoosmotic environment, 108,109 a feature of which is the high cellular turnover rate 110 that occurs throughout the entire length of the gill. 111 Therefore, the upregulation of gasdermin would appear to highly beneficial in maintaining the integrity of the trout gill.
One unique feature of the communication between the irradiated trout prior to irradiation and the completely nonirradiated fish was the upregulation of proteins involved in neuroprotection. This is of particular importance given that fish gills are innervated by cranial nerves IX and X. 112,113 Fish gills are also known to contain neuroepithelial cells. 114 Tubulin controls the growth of axons 115,116 and is involved in spinal cord recovery. 117 A related protein, tubulin α, specifically, regulates neurotubule formation in fish. 118 Since hOR upregulation, in the gills of trout which swam with irradiated fish prior to irradiation, could result in some degree of oxidative stress (see above) which, in turn, can result in a reduction in β-tubulin, 119 the upregulation of tubulin β-1 chain could be interpreted as a compensatory response. Glycine-tRNA ligase β subunit/glycyl-tRNA synthetase is another neuroregulatory protein. tRNA charging enzymes are essential in maintaining peripheral neurons. Decreased glycyl-tRNA activity (ie, the opposite of the upregulation seen in the present study) is characteristic of peripheral nervous system diseases 120 and mutations in glycyl-tRNA result in peripheral nervous system disorders, for example, Charcot-Marie-Tooth neuropathy. 121,122
Malignant central nervous system tumors show no tropomyosin expression. 123 This would suggest the downregulation of tropomyosin α-1 chain/β chain and tropomysosin α-3 chain, in the gills of trout which swam with irradiated fish prior to irradiation, is a detrimental, potentially pro-tumorigenic response. However, conversely, tropomyosin is upregulated in neoplasic astrocytes, 123 tropomyosin-related kinase downregulation inhibits cancer cell growth. 124 Tropomyosin is also involved in stress fiber formation. 123 Thus, as was the case with IFP, we propose that the role of tropomyosin as a response to communication between nonirradiated fish and fish which were to be irradiated warrants additional experimental attention.
We have divided the remainder of this discussion into 2 sections based on our 2 major hypotheses to explain the data.
Explanations Involving Social Interactions With Inducible Stress Responses
Fish are social organisms and a shoal is often described as a superorganism due to the level of coordination, which is thought to involve inter-fish communication by physical, chemical, or visual signals. 125 –127 One explanation which has some merit is the idea that exposure to a small dose of a stressor after rather than before a large dose of the stressor can induce protection. This phenomenon known as postconditioning has been documented 128,129 and is thought to be a form of temporal adaptive response possibly involving hormetic mechanisms where the time sequence of the 2 doses is less important than the fact that a small dose is administered. This implies that the mechanisms induced by the high dose are different to the mechanisms induced by the low dose—an idea which is becoming popular in the radiation field. 130,131 Neurobiological responses to stressors could be involved since it is now apparent that the initial trigger for the bystander signal can be physical involving excitation decay and light production. 132,133 In the particular case of the fish in this study, hierarchical dominance behaviors due to the sudden meeting of stranger populations of fish might explain the data. Effects stimulated by population mixing could also be a factor since there is some evidence that immune responses can be altered, resulting in immune and inflammatory system level effects when populations suddenly meet. 134 The key point in all above hypotheses is that the controls which swam together without any irradiation should show at least some effect of meeting a strange population of fish which they do.
Quantum Entanglement Explanations
The second and potentially more exciting idea involves consideration of a hypothesis, involving biological quantum entanglement. Quantum effects are well established in modern physics and increasingly are being investigated in biology where they are thought to fast track processes in photosynthesis, respiration, neurobiology, and signalling. 135,136 They are especially relevant where excitation, energy transduction, and excitation decay are involved in the mechanism as they are in the case of bystander-based communication. However, although these processes could well be involved in the bystander mechanism at the molecular level, it is hard to conceptualize a quantum-based mechanism operating at the level of the organism.
To conclude, the results using 3 different assay systems and 2 species of fish reveal an unexpected effect in entangled fish which met before or after 1 group was irradiated. The signaling between irradiated and unirradiated fish has been well-documented but effects in bystander fish which met before irradiation of part of the group is unexpected. Two suggestions to explain the data are put forward—first that it is a form of postconditioning effect and second that quantum biology is involved in the BE in these fish.
Footnotes
Acknowledgments
The authors acknowledge support from the Canada Research Chairs program, the National Science and Engineering Research Council (NSERC) Collaborative Research and Development program and NSERC Discovery Grants program, the National CFIDS (Chronic Fatigue and Immune Deficiency Syndrome) Foundation Inc, Bruce Power and the CANDU Owner’s Group (COG).
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.