
Abstract
Duchenne muscular dystrophy (DMD) is a recessive X-linked fatal disorder caused by a mutation in the dystrophin gene. Although several therapeutic approaches have been studied, none has led to substantial long-term effects in patients. The aim of this study was to test a serotonin and histamine (S&H) combination on human skeletal myoblasts and Dmdmdx mice for its effects on muscle strength and injury. Normal human bioartificial muscles (BAMs) were treated, and muscle tetanic forces and muscle injury tests were performed using the MyoForce Analysis System. Dmdmdx mice, the murine model of DMD, were administered serotonin, histamine, or S&H combination twice daily for 6 weeks, and functional performance tests were conducted once a week. The S&H combination treatment caused significant increases in tetanic forces at all time points and concentrations tested as compared to the saline controls. Dose response of the BAMs to the treatment demonstrated a significant increase in force generation at all concentrations compared to the controls after 3 to 4 days of drug treatment. The highest 3 concentrations had a significant effect on lowering contractile-induced injury as measured by a reduction in the release of adenylate kinase. Histamine-only and S&H treatments improved grip strength of Dmdmdx mice, whereas serotonin-only treatment resulted in no significant improvement in muscle strength. The results of this study indicate that S&H therapy might be a promising new strategy for muscular dystrophies and that the mechanism should be further investigated.
Keywords
Introduction
Duchenne muscular dystrophy (DMD) is an X-linked recessive disorder characterized by progressive and severe wasting and weakness of nearly all muscles leading to premature death by cardiorespiratory failure. 1 Duchenne muscular dystrophy has a prevalence of 1 in 5,000 newborn males worldwide and is caused by the truncation or absence of dystrophin protein that anchors the muscle fiber to the extracellular matrix (ECM). 2 -4 Since the mechanical link between the contractile apparatus and the plasma membrane is disrupted, dystrophic myofibers are more susceptible to contraction-induced injury and undergo continuous necrosis and repair until the muscle mass is replaced by fat and fibrous tissue. 5,6 The only clinically validated treatments for DMD are glucocorticoids that prolong muscle strength for short terms but have severe adverse effects. 7 Although research continues to investigate downstream mechanisms, such as muscle necrosis, inflammation, fibrosis, or regeneration, most recent therapeutic approaches focus on individual gene therapies that promote stop codon read-through or correct frameshift mutations. 8 -11 Drugs have also been developed to upregulate surrogate proteins, such as utrophin, which can substitute for dystrophin at the muscle cell membrane. 12
Several mammalian and nonmammalian animal models have been developed and used extensively to assess the therapeutic efficacy of candidate drugs. 13 -15 These animal models made vital contributions to understanding the molecular processes underlying the disease pathology, and the Dmdmdx mouse is the most commonly used murine model in preclinical studies. 16 -18 Tissue engineering of contracting muscle fibers is also a unique tool for in vitro drug screening and is instrumental for basic research to identify new therapies. 19,20 Understanding and modifying the pathological background of DMD and the complex interactions of all muscle cells with their surrounding environment are important for the management of the disease. Changes in this environment can speed up muscle repair, enhance muscle strength, and reduce inflammation and fibrosis, leading to the amelioration of symptoms. 21,22 Therefore, molecules that can modify these complex interactions between muscle cells and their environment can be powerful therapeutics for the treatment of DMD. Histamine is best known as a mediator of allergic reactions, but it is also recognized to participate in numerous physiological processes such as regulation of the immune response, smooth muscle contraction, secretion of gastric acids, and excitatory glutamate neurotransmission in the spinal cord. 23 -27 The neurotransmitter serotonin mediates a wide variety of central and peripheral responses via multiple serotonin receptors. Depending on the secondary messengers with which the receptors are coupled, these responses can regulate various physiological processes such as cognition, cardiovascular functioning, vasoconstriction, learning and memory, and skeletal muscle repair. 28 -30
Based on the positive indications of serotonin and histamine (S&H) on muscle physiology, we set out to study whether these molecules could serve as potential strategies in the treatment of DMD. Therefore, the current investigation was designed to determine the effect of S&H on muscle strength and muscle recovery from contractile-induced injury. Specifically, we measured the active force generation and recovery from the injury induced by electrical stimulation of contracting muscles in a 3-dimensional (3D) cell culture model. Additionally, Dmdmdx mice forelimb performance tests were used to determine whether serotonin, histamine, or S&H combination has any effect on muscle strength in vivo.
Materials and Methods
Cell Culture and Tissue Engineering
Human skeletal muscle cells previously isolated from a disease-free 48-year-old female were used in this study (Myomics Inc, Providence, Rhode Island). The cells were expanded in cell culture in a growth medium optimized for human skeletal muscle, SkGM Bullet kit (Lonza, Allendale, New Jersey), supplemented with 15% fetal bovine serum (Gibco, Grand Island, New York). The cells were cultured in this medium for 5 days before being engineered into bioartificial muscles (BAMs). Histamine was purchased from HollisterStier (Spokane, Washington), serotonin from Sigma-Aldrich (St Louis, Missouri), and phosphate-buffered saline from Hospira (Lake Forest, Illinois). Two micrograms of serotonin is designated as a 1× dose, and 1.1 μg of histamine is designed as a 1× dose; therefore, the 1× S&H combination treatment contained 2 μg serotonin and 1.1 μg histamine.
The detailed description of the muscle engineering was published previously. 19 Briefly, on the day of engineering, BAM cells were trypsinized and counted with a hemocytometer. A sterile 96-well MyoForce plate (Myomics Inc) was used for tissue engineering the BAMs. Myomics’ Robotic Tissue Engineering Platform was used to engineer 72 BAMs in approximately 10 minutes by mixing cells with an ECM solution and casting in the plate wells. The plate containing the BAMs was maintained for 2 days at 37°C in SkGM/15 and then incubated in an optimized differentiation medium consisting of Dulbecco modified Eagle medium (Life Technologies, Grand Island, New York), 0.1% insulin (Sigma, St Louis, Missouri), 1% bovine serum albumin (Sigma), 0.1% human epidermal growth factor (Life Technologies), and 0.5% gentamicin (Sigma). The cells were kept in the optimized differentiation medium for the remainder of the experiment.
Active Force Measurements
On day 9 after plating the cells, the BAM plates were transferred to the Myomics’ Myoforce Assay Device (MAD). The BAMs were electrically stimulated using parameters set for generating maximal muscle contraction, that is, tetanic (active) force. The MAD took approximately 15 minutes to stimulate all the wells in each 96-well MyoForce plate. A high-speed imaging camera captured the images of the BAMs as they contracted and sent the data to the computer system calculating micronewtons (μN) of force generated. After the initial electrical stimulation, the combination treatment (S&H) was added to each well to final concentrations of 0.01×, 0.1×, 1×, 2.5×, 5×, and 10×. Other treatment groups included in the assay were a positive control 80 nmol/L deflazacort (DFZ; a glucocorticoid used as an anti-inflammatory and immunosuppressant), and a vehicle-only control (saline). Each treatment and control group was assayed with 8 replicates, and the experiments were conducted 3 times.
Injury Assay
To assess the effect of S&H combination on muscle injury, BAMs were subjected to repetitive tetanic electrical stimulations in the MAD after 4 days of drug treatment. Each BAM received a total of 16 consecutive electrical pulses. ToxiLight BioAssay (LT07-217; Lonza, Allendale, NJ) was used to measure the release of adenylate kinase (AK) caused by the leakage from the damaged plasma membrane of the muscle cells. The ToxiLight BioAssay measures the conversion of adenosine diphosphate to adenosine triphosphate (ATP) in the presence of AK. The enzyme luciferase then catalyzes the formation of light from luciferin in the presence of ATP. The emitted light, measured with the luminometer, is linearly related to the AK concentration; therefore, the light intensity is indicative of the extent of damage to the cell membrane. Conditioned medium was collected from the BAMs after injury for assay of released AK; BAMs were then lysed, centrifuged, and the supernatant was collected to determine the intracellular AK. The AK released upon injury was calculated as the percentage of the total AK (released plus intracellular).
Animal Care and Treatments
The animal studies were conducted at the Animal Care Facilities of Brown University in Providence, Rhode Island. Four-week-old female C57BL/10ScSn-Dmdmdx/J dystrophic mice and C57BL/10ScSnJ nondystrophic control mice were purchased from Jackson Laboratories (Bar Harbor, Maine). The animals were maintained under controlled conditions (19°C-23°C, 12-hour light/dark cycles) and allowed water and food ad libitum. Treatment and control groups contained 7 and 5 mice per cage, respectively. All procedures involving animals were approved and performed in accordance with the guidelines of the Institutional Animal Care and Use Committee (IACUC) of Brown University in compliance with the guidelines established by the National Institutes of Health. The animals were randomly assigned to all cages prior to the experiment. Following the randomization, animals were injected subcutaneously in the middorsal region twice daily with 0.1 mL treatment or saline for the duration of the study. The concentrations used in the animal experiments were determined by a preliminary study where a spectrum of S&H doses was screened by muscle function tests in a small group of Dmdmdx mice. Based on the preliminary work, various concentrations of serotonin, histamine, and the S&H combination were chosen to be tested in this study (0.4×, 1×, 4×, and 8×).
Grip Strength Measurements
A grip strength meter (Columbus Instruments, Columbus, Ohio) was used to assess neuromuscular function by sensing the peak amount of force an animal applies in grasping a specially designed pull bar assembly (triangle bar). In practice, the grip strength meter was used by first allowing the animal to grasp the forelimb triangle pull bar assembly. The animal was then drawn linearly, leading away from the sensor. At the time of the release of the bar, the sensor recorded the maximum force attained. Measurements were performed once a week, and readings were obtained in micro newtons; 10 readings were collected per animal.
Clonogenic Assay
Human skeletal muscle cells were plated at 200 cells/dish. Then, cells were allowed to adhere to the bottom of the dishes overnight. Following day, cells were continuously exposed to various concentrations of S&H for 10 days. At the end of 10 days, colonies were fixed with ice cold methanol, stained with crystal violet, and counted.
Statistical Analysis
For the cell culture studies, the mean standard error (SE) of 8 samples per group was calculated, and t-test statistical analyses were performed using SigmaStat software (Systat Software, Inc, Chicago, Illinois), with P < .05 considered statistically significant. Power analyses were performed using GraphPad StatMate 2.00, and half-maximal effective concentration (EC50) values were determined with GraphPad Prism (GraphPad Software, Inc, San Diego, California). One-way analysis of variance was performed to determine the differences between the groups in animal studies. Differences between the groups were considered significant with a P value <.05.
Results
Combination Therapy Increases Tetanic Forces
The BAMs were treated with 6 different concentrations of S&H combinations on day 9 for the next 4 days. Media content of the plates was replaced daily during the treatment, and percent changes in the tetanic forces relative to the vehicle control were recorded. Time-course experiments of treatment with S&H combination showed significant increases in tetanic forces at all time points, and all doses tested were compared to the vehicle controls (Figure 1A and B). Positive control, 80 nmol/L DFZ, showed a significant increase (32%) in tetanic forces during the 4 days of treatment compared to the vehicle control, validating the strength of the assay.

A and B, Percent change in tetanic forces as compared to the untreated controls. Effect of the 3 lowest (A) and the 3 highest concentrations (B) of the serotonin and histamine (S&H) combination on the bioartificial muscles’ (BAM) force-generating ability. Deflazacort (DFZ) was used as a positive control. Data are expressed as the mean ± standard error (SE) of the percent change in active force compared to the vehicle controls (saline only) at each time point (n = 8 per group). Percents represent average of 3 independent experiments.
Dose Response on Day 3 and Day 4
Following 3 and 4 days of drug treatment, percent change in the tetanic forces was recorded. Dose responses of BAMs to the S&H combination showed significant increases in force generation at all concentrations tested compared to the saline control after 3 to 4 days of treatment, with the increases ranging from 20.9% to 40.8%. The highest increases in force generation were seen with 0.1× S&H (40.8% day 3 and 35.7% day 4) and 2.5× S&H (35.0% day 3 and 28.8% day 4; Figure 2).

Dose response to serotonin and histamine (S&H) combination after 3 and 4 days of treatment. Dose response of force-generating ability of bioartificial muscles (BAMs) to S&H after 3 and 4 days of drug treatment. Data are expressed as the mean ± standard error (SE) of the percent change in active force versus treatment concentration compared to the vehicle controls (n = 8 per group). Percents represent average of 3 independent experiments.
Combination Therapy Reduces Muscle Injury
Following 4 days of drug treatment, BAMs were subjected to repetitive tetanic electrical stimulation in the MAD. The S&H combination treatments of 2.5×, 5×, and 10× had a significant effect on reducing injury to the BAMs as measured by the decrease in the release of AK by the ToxiLight assay compared to the untreated controls (Figure 3). In contrast, BAMs treated with 0.01×, 0.1×, and 1× combinations of S&H did not show significant reduction in AK release as compared to the untreated controls (Figure 3).

Percent release of adenylate kinase from injured bioartificial muscles (BAMs) treated with serotonin and histamine (S&H) combination. After 4 days of treatment with S&H, BAMs were subjected to repetitive electrical stimulation resulting in contraction-induced injury. Data are expressed as the mean ± standard error (SE) of the percent of adenylate kinase released upon injury. Controls were injured muscles treated with vehicle control (saline), uninjured vehicle-treated BAMs, and injured muscles treated with deflazacort (DFZ). Percents represent average of 3 independent experiments.
Histamine-Only and Combination Therapy Increase Grip Strength Performance of mdx Mice
Four-week-old mdx mice were treated with various concentrations (0.4×, 1×, 4×, and 8×) of serotonin, histamine, or S&H combination for 6 weeks twice daily. Mdx mice treated with the 4× S&H combination showed transient increase in forelimb strength on week 1 and week 3 as compared to the saline-treated animals. Interestingly, the 8× S&H combination caused significant improvement in forelimb strength only at week 6, suggesting that the S&H combination therapy has positive effects on muscle strength; however, this positive effect was not sustainable throughout the treatment period (Figure 4A). Histamine-only treatment increased muscle strength throughout the treatment with the exception of week 1. Among the 4 concentrations of histamine tested, 8× histamine consistently increased muscle strength of mdx mice, whereas 4× was effective on weeks 4, 5, and 6 (Figure 4B). Serotonin by itself did not increase muscle strength of mdx mice throughout the study period (Figure 4C).
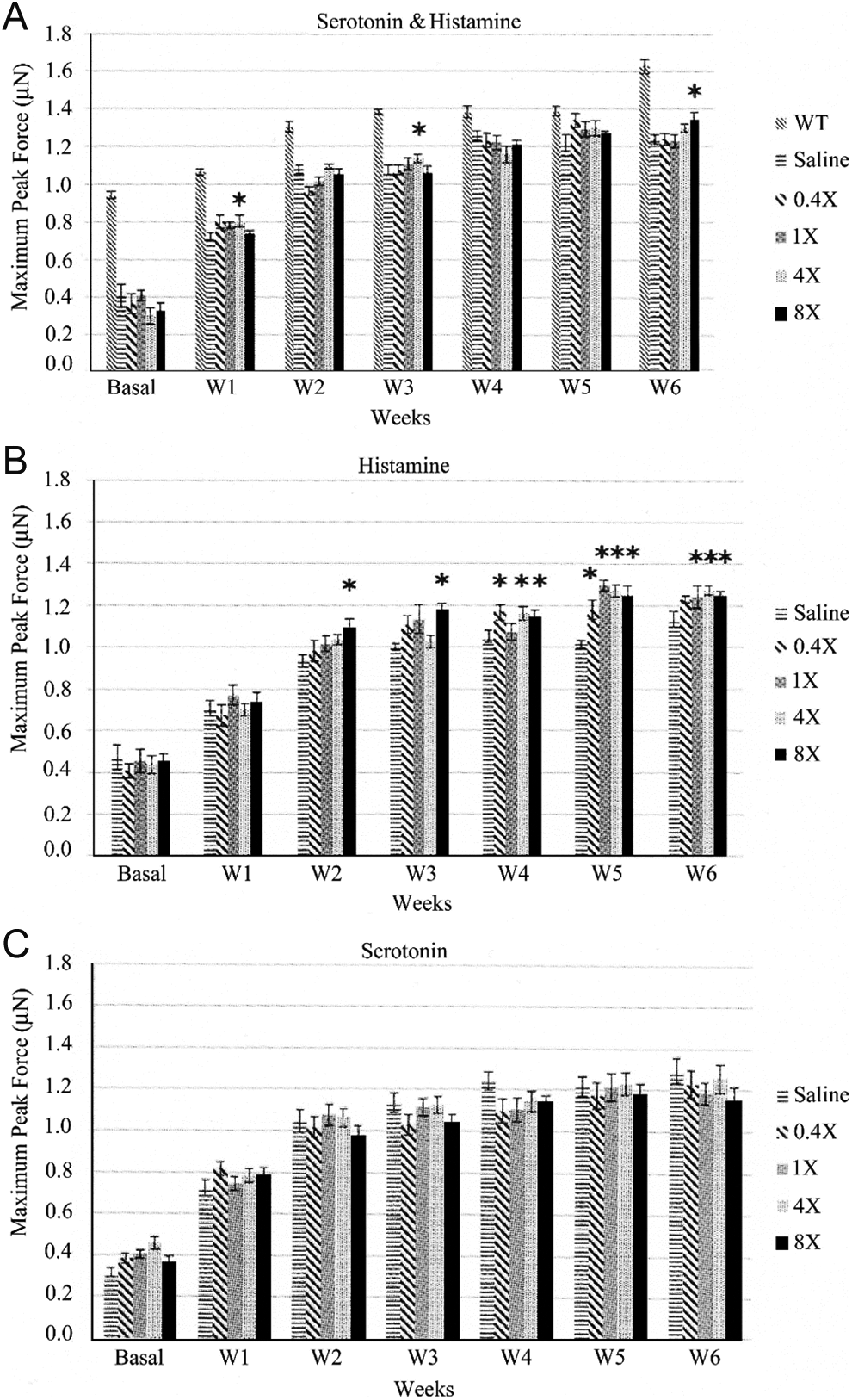
A, B, and C, Weekly forelimb muscle strength tests determined by muscle strength meter for (A) serotonin and histamine (S&H) combination, (B) histamine-only, and (C) serotonin-only. Mice were treated subcutaneously with 0.4×, 1×, 4×, and 8× histamine, serotonin, or S&H twice daily for 6 weeks. WT represents C57BL/10ScSnJ nondystrophic control mice. *Statistical significance at P < .05.
We did not observe any lethal side effects in the treated animals, and the levels of S&H used in this study were lower than the toxicity levels reported in several animal studies in the literature. 31 -33 Average weights of the animals did not show statistically significant differences between the treated and untreated mice (Figure 5A-C). Also, no sign of toxicity was detected with human skeletal muscle cells in 3D cell culture as assessed by Calcein AM viability assays and clonogenic assay (Figure 6).

A, B, and C, Average weights of mice in (A) serotonin and histamine (S&H) combination, (B) histamine-only, and (C) serotonin-only treatment groups. Mice in each treatment group were weighed and averages were recorded for each cage for 6 weeks. The line graphs represent the weight changes (g) for each group for 6 weeks.

Clonogenic assay. Human skeletal muscle cells were exposed to various concentrations of serotonin and histamine (S&H) combination for 10 days, and colonies were counted. Each concentration had 3 replicates, and the values in the final graph represent the average from the 3 replicates.
Discussion
Duchenne muscular dystrophy is the most common, severe childhood form of dystrophy in which the treatment is limited to glucocorticoids that have severe side effects. Multiple treatment approaches have emerged, and the most attractive ones are the molecular-based therapies that can either restore the mutant dystrophin protein or enhance the levels of surrogate proteins. Although the future looks bright for the multiple molecular strategies, there is still need for new therapies with lower side effects targeting downstream mechanisms such as inflammation, fibrosis, and repair. The goal of this study was to determine the effects of S&H on muscle tetanic forces and muscle recovery from contractile injury using engineered BAMs. Dmdmdx, a murine model of DMD, was used to test whether the experimental therapy provides improved functional performance scores in forelimb pull tests. We observed that although all S&H concentrations tested increased tetanic forces of muscle cells (Figure 1A and B), only the highest 3 concentrations provided protection from contractile-induced muscle injury (Figure 3). Dystrophic mice treated with S&H combination and histamine-only performed better in forelimb muscle pull tests, whereas the test scores of serotonin-only-treated mice were not different from that of the saline-treated animals.
Drug screening with engineered tissues based on physiological function culminates all the complex interactions of compounds on multiple intracellular pathways. Therefore, it is a better in vitro model than most high-throughput drug screening technologies in bridging with in vivo animal models to predict the ultimate effects of the compounds. This technology has been used previously to identify or repurpose compounds with established human safety profiles for new potential treatments for muscle weakness in DMD. 34
5-hydroxytryptamine receptor 2A(5-HT2A) is the only functional serotonin receptor that has been identified and localized in skeletal muscle cells. 35,36 Binding of serotonin to 5-HT2A receptor leads to upregulation of genes involved in myogenic differentiation and activation of the JAK/STAT pathway, particularly the transcription activator STAT3 gene. 35 STAT3 activation is known to be the key regulator of a cascade of genes in skeletal muscle ontogenesis and repair pathways. 37 -39 A critical role of serotonin in cell repair has also been demonstrated in Caenorhabditis elegans. 40 In C elegans, serotonin has been shown to be a potent suppressor of striated muscle degeneration, with serotonin being more efficient than prednisone, a palliative treatment against DMD, in blocking the muscle degeneration. 41 These mechanisms could partly explain the outcomes observed with the 3 highest concentrations of S&H combination which reduced the contractile-induced injury of muscle cells by more than 50% compared to the untreated controls (Figure 3). This suggests a potential involvement of S&H in the repair pathway, possibly through Jak/STAT activation. However, this point requires further molecular exploration to confirm the actual activation of the pathway.
Although no evidence exists involving skeletal muscle cells in the literature, in vitro studies conducted with airway smooth muscle cells and cardiomyocytes demonstrated that histamine causes a transient rise in cytosolic Ca2+ levels via the H1 receptor. 42,43 High intracellular Ca2+ levels are crucial for triggering muscle contraction, whereas low levels are critical for initiating muscle relaxation. 44 Additionally, both histamine and peripheral serotonin are important regulators of glucose transport. 45 Histamine has been shown to increase glucose uptake in cultured cardiac microvascular endothelial cells via activation of H1 receptors. 46 Histamine receptors H1 and H2 have been suggested to enhance the availability of glucose to skeletal muscles by facilitating capillary permeability to the interstitium following exercise. 47 -49 Unlike histamine, serotonin can directly increase glucose uptake by coupling with 5-HT2A receptors located on skeletal muscle cells. Taken together, S&H can modulate intracellular Ca2+ and glucose levels, possibly accounting for the increased tetanic forces and resistance to injury we observed in the 3D culture model following S&H treatment. Moreover, serotonin receptor stimulators have been found to be effective in restoring respiratory abnormalities in rats. Thus, the cardiorespiratory symptoms of DMD potentially can be potentially resolved with S&H therapy by the combinatorial effects on the cardiovascular and respiratory systems. 50 However, the approach must be refined considerably to allow for safe translation into clinical development.
Muscle from DMDmdx mice is histologically indistinguishable from that of normal mice early in postnatal development. Between weeks 3 and 6, the muscle undergoes severe degeneration and becomes hypertrophic in the limbs. Subsequently, the majority of the skeletal muscle enters a relatively stable phase due to robust regeneration through adulthood. Yet, despite the muscles’ large size, they are comparatively weaker since their maximum force output per muscle cross-sectional area is usually lower. 51,52 This explains the observed weight differences and the performance results between the wild-type and dystrophic mice in this study (Figure 5A). Interestingly, serotonin-only treatment did not cause an increase in muscle strength throughout the study, suggesting that the role of serotonin in muscle repair is not enough to increase muscle strength. Combination therapy was somewhat effective, and the histamine-only treated mice had the best scores compared to the other 2 treatments. This is possibly because histamine is able to regulate a broader range of cellular activities compared to serotonin, such as cytosolic Ca2+ levels, glucose transport, neurotransmission, and vasodilation.
There is some correlation between mutations in the DMD gene and the resulting disease severity in human patients. The vast majority of DMD mutations result in the complete absence of dystrophin, whereas the presence of low levels of a truncated protein results in the milder form of the disease, Becker muscular dystrophy. In addition to these diseases, mutations in the genes encoding many components of the dystrophin-associated protein complex cause other forms of muscular dystrophy such as the limb-girdle muscular dystrophies and congenital muscular dystrophy. The Mdx mouse lacks full-length dystrophin protein because of a point mutation in exon 23 of the DMD gene, which forms a premature stop codon. 53 However, the mdx mouse still retains expression of some COOH-terminal dystrophin isoforms. Interestingly, mice also lacking this COOH- terminal display phenotypical similarities to the mdx mouse, arguing that the full-length dystrophin is the functionally significant isoform in mouse muscle and partially functioning protein does not result in milder disease phenotype. 54 Therefore, testing the therapeutic effect of S&H in less severe forms of muscular dystrophy in a mouse model remains elusive.
To the best of our knowledge, this is the first report of the use of S&H in both 3D functional muscle tests and an animal model of DMD to assess muscle strength and repair. The present study suggests that there is a foundation for further investigation of S&H or histamine-only therapy which can potentially increase muscle strength and reduce contractile-induced injury. Although there is a need for more extensive in vivo and in vitro work, including investigating the gene expression profiles, muscle histology, and cardiorespiratory assessments, this report provides valuable preliminary insight for future studies.
Footnotes
Acknowledgments
The authors thank Christoph Schorl, PhD, and Herman Vandenburgh, PhD, from Brown University for their critical contribution in conducting the experiments and writing this article.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.