
Abstract
Background. The positive correlation between therapeutic exercise and memory recovery in cases of ischemia has been extensively studied; however, long-term exercise begun after ischemic neuronal death as a chronic neurorestorative strategy has not yet been thoroughly examined. Objective. The purpose of this study is to investigate possible mechanisms by which exercise ameliorates ischemia-induced memory impairment in the aged gerbil hippocampus after transient cerebral ischemia. Methods. Treadmill exercise was begun 5 days after ischemia-reperfusion (I-R) and lasted for 1 or 4 weeks. The animals were sacrificed 31 days after the induction of ischemia. Changes in short-term memory, as well as the hippocampal expression of markers of cell proliferation, neuroblast differentiation, neurogenesis, myelin and microvessel repair, and growth factors were examined by immunohistochemistry and/or western blots. Results. Four weeks of exercise facilitated memory recovery despite neuronal damage in the stratum pyramidale (SP) of the hippocampal CA1 region and in the polymorphic layer (PoL) of the dentate gyrus (DG) after I-R. Long-term exercise enhanced cell proliferation and neuroblast differentiation in a time-dependent manner, and newly generated mature cells were found in the granule cell layer of the DG, but not in the SP of the CA1 region or in the PoL of the DG. In addition, long-term exercise ameliorated ischemia-induced damage of myelin and microvessels, which was correlated with increased BDNF expression in the CA1 region and the DG. Conclusions. These results suggest that long-term treadmill exercise after I-R can restore memory function through replacement of multiple damaged structures in the ischemic aged hippocampus.
Introduction
Vascular cognitive impairment is defined as mild to severe cognitive impairment of cerebrovascular origin such as stroke, multiple cortical infarcts, and small-vessel disease.1,2 Stroke is the second leading cause of death among the elderly, and the number of stroke patients is steadily increasing.3-5 Because of the high prevalence of cerebrovascular disease in the elderly population, stroke is one of the most common problems for the elderly. Ischemic stroke or transient ischemic attacks can occur following transient cerebral ischemia and cause defects in memory.6,7 In particular, neurological deterioration of the hippocampus can lead to loss of memory and other cognitive functions because the hippocampus plays an important role in learning and memory.8-10
Therapeutic exercise is recommended for the rehabilitation of patients with stroke; it has been shown to minimize the severity of functional deficits and reduce the risk of recurrence.11,12 The positive correlation between physical exercise and memory recovery has been extensively studied in animal models of ischemic insults.9,13,14 Physical exercise is associated with an enhancement in neurotrophin and growth factor expressions, synaptogenesis, neurogenesis, and angiogenesis.9,13-19
Most previous studies on therapeutic exercise for stroke recovery have been conducted in young adult animals,20-22 and few studies have been done in aged animals even though there is a high prevalence of cerebrovascular disease in the elderly population. In addition, in many reports, therapeutic exercise was begun before ischemia-induced neuronal death as an acute neuroprotective treatment23,24; however, the effects of long-term exercise begun after ischemic neuronal death as a chronic neurorestorative strategy have not been extensively studied.
Gerbils are used as an animal model of transient cerebral ischemia by the deprivation of blood flow to the brain through bilateral carotid arteries occlusion, which results in neuronal damage/death in several vulnerable regions including the hippocampus.25-27 Therefore, we investigated the effects of long-term treadmill exercise begun after transient ischemic insult on neuronal and memory recovery in the aged gerbil hippocampus. We also investigated cell proliferation, neuroblast differentiation, neurogenesis, myelin and microvessel repair, and the expression of growth factors to elucidate the possible mechanism of restorative exercise in neuronal and memory recovery.
Materials and Methods
Experimental Animals
Aged male Mongolian gerbils (Meriones unguiculatus, 22-24 months) weighing 80 to 90 g were supplied by the Experimental Animal Center, Kangwon National University (Chuncheon, South Korea). Animal handling and care followed the guidelines of current international laws and policies (NIH Guide for the Care and Use of Laboratory Animals, The National Academies Press, 8th ed., 2011), 28 and experimental protocols were approved by Institutional Animal Care and Use Committee of Kangwon National University (Approval No. KW-130424-1).
The gerbils were randomly divided into 5 groups (n = 63): (1) sham-operated group (sham group, n = 14); (2) ischemia-operated group (ischemia group), sacrificed to examine neuronal death 5 days following ischemia (n = 7); (3) SD4 group had 4 weeks sedentary (SD) from 5 days postischemia (n = 14); (4) TR1 group has 1 week treadmill (TR) from 5 days postischemia (n = 14); and (5) TR4 group had 4 weeks TR from 5 days postischemia (n = 14). The animals were sacrificed 31 days after ischemia when the treadmill exercise training was completed in the TR4 group.
Induction of Transient Cerebral Ischemia
As previously described, 29 in brief, the animals were anesthetized and bilateral common carotid arteries were occluded 5 minutes using nontraumatic aneurysm clips (Yasargil FE 723K; Aesculap, Tuttlingen, Germany; see online supplement). Sham-operated animals were subjected to the same surgical procedures except for the bilateral common carotid arteries occlusion.
Treadmill Exercise
The running speed and durations were determined according to Sim’s protocol,20-22 with modification. The treadmill exercise group was familiarized with the treadmill running on a motorized treadmill (Exer3/6, Columbus Instruments, Columbus, OH) for 15 min/day for 3 consecutive days. From 5 days after I-R, treadmill exercise groups were forced to run on a motorized treadmill for 30 min/day and 5 days/week for 1 or 4 consecutive weeks. The exercise workload consisted of running at the speed 5 m/min for the first 5 minutes, 7 m/min for the next 5 minutes, and then 10 m/min for the last 20 minutes with 0° inclination. Animals in the sedentary groups were left on the treadmill for 30 minutes and were not made to run.
Passive Avoidance Test
The short-term memory ability was evaluated by assessing the latency of the passive avoidance test (PAT) with an apparatus that consisted of 2 compartments (light and dark) with a grid floor (GEM 392, San Diego Instruments, San Diego, CA). The test was performed according to a previous study 30 (see online supplement); the PAT (training and test sessions) was performed 1 day before I-R and 5, 10, and 31 days after I-R repeatedly in the sham, SD4, TR1, and TR4 groups (n = 14 in each group).
BrdU Administration
To examine cumulative labeling of newly generated cell in the dentate gyrus (DG) after ischemia, animals (n = 7 at each time in each group) received intraperitoneal BrdU injection (50 mg/kg in saline) 3 consecutive days from 4 days after I-R. They were then sacrificed 31 days after ischemia when the treadmill exercise training was completed in the TR4 group.
Tissue Processing for Histology
The animals (n = 7 each group) were anesthetized with sodium pentobarbital (JW Pharm Co, Ltd, Seoul, Korea, 40 mg/kg, ip) and perfused transcardially with 0.1 M phosphate-buffered saline (PBS; pH 7.4) followed by 4% paraformaldehyde in 0.1 M phosphate buffer (PB; pH 7.4). The brain tissues were serially sectioned on a cryostat (Leica, Wetzlar, Germany) into 30-µm coronal sections.
General Immunohistochemistry
According to the method of our previous study, 31 immunostaining was performed using mouse anti-neuronal nuclei (NeuN; 1:1000, Chemicon International, Temecula, CA), rabbit anti-doublecortin (DCX; 1:50, Santa Cruz Biotechnology, Santa Cruz, CA), mouse anti-BrdU (1:250, Rosche, Basel, Switzerland), rabbit anti-myelin basic protein (MBP; 1:1000, Abcam, Cambridge, UK), rabbit anti-oligodendrocyte specific protein (OSP, 1:500, Abcam, Cambridge), rabbit anti-glucose transporter (GLUT)-1 (1:1000, Chemicon International), rabbit anti-brain derived neurotrophic factor (BDNF; 1:500, Abcam) or rabbit anti-insulin like growth factor (IGF)-1 (1:200, Santa Cruz Biotechnology), biotinylated goat anti-rabbit, or horse anti-mouse IgG (Vector, Burlingame, CA) and streptavidin peroxidase complex (1:200, Vector).
Double Immunofluorescence Staining for BrdU and NeuN
The sections were incubated with the mixture of mouse anti-BrdU (1:150, Rosche) and rabbit anti-NeuN (1:400, Millipore, Billerica, MA) after the denaturation process of DNA for BrdU detection and incubated in a mixture of both FITC-conjugated goat anti-mouse IgG (1:200, Jackson ImmunoResearch, West Grove, PA) and Cy3-conjugated goat anti-rabbit IgG (1:200, Jackson ImmunoResearch). The immunoreactions were observed under a confocal microscope (LSM510 META NLO, Carl Zeiss, Oberkochen, Germany). A series of high magnification images were collected at 1-µm intervals with excitation by 488 and 543 nm lasers to create a stack in the z-axis.
Cell Counting and Immunoreactivity Measurement
In order to quantitatively analyze NeuN immunoreactive neurons, DCX immunoreactive neuroblasts, and BrdU-positive cells, digital images from 6 sections per animal were taken using an AxioM1 light microscope (Carl Zeiss) equipped with a digital camera (Axiocam, Carl Zeiss) connected to a PC monitor. NeuN immunoreactive neurons were counted in a 250 × 250 µm square, applied approximately at the center of the CA1 region or DG, and DCX- and BrdU-positive cells were counted in whole DG using an image analyzing system (Optimas 6.5, CyberMetrics, Scottsdale, AZ). Cell counts were obtained by averaging the counts from each animal.
To quantitatively analyze MBP, OSP, GLUT-1, BDNF, and IGF-1 immunoreactivities, digital images of the corresponding areas were captured with an AxioM1 light microscope (Carl Zeiss) equipped with a digital camera (Axiocam, Carl Zeiss) connected to a PC monitor. The densities of all MBP, OSP, GLUT-1, BDNF, and IGF-1 immunoreactive structures were evaluated on the basis of optical density (OD), which was obtained after the transformation of the mean gray level using the following formula: OD = log(256/mean gray level). After the background was subtracted, a ratio of the OD of image file was calibrated as percentage (relative optical density [ROD]) using Adobe Photoshop version 8.0 and NIH Image J software (National Institutes of Health, Bethesda, MD). The mean value of the OD of the sham group was designated as 100%, and the ROD of each group was calibrated and expressed as percentage of the sham group.
Western Blot Analysis for GLUT-1 and BDNF
As previously described, 32 in brief, after removing the brains, the hippocampus of the animals (n = 7 in each group) was dissected. After the tissues were homogenized and centrifuged, the supernatants were subjected to western blot analysis. Rabbit anti-GLUT-1 (1:1000, Chemicon International) or rabbit anti-BDNF (1:1000, Abcam) were used as primary antibody. The result of western blot analysis was scanned, and densitometric analysis for the quantification of the bands was done using Image J 1.46 (National Institutes of Health), which was used to count ROD: A ratio of the ROD was calibrated as percentage, with sham group designated as 100%.
Statistical Analysis
Data are expressed as the mean ± SEM. All comparisons were tested for normality and variance homogeneity using SPSS 17.0 software. The latency of the short-term memory ability were analyzed with a 2-way repeated analysis of variance (ANOVA), followed by post hoc Bonferroni-Dunn test. Mean numbers of immunoreactive structures and ROD of immunoreactive structures were analyzed with a one-way ANOVA, followed by post hoc Bonferroni-Dunn test. Statistical significance was considered at P < .05.
Results
Passive Avoidance Test
Five days after I-R, the latency in the PAT was significantly decreased (P < .05) in the SD4, TR1, and TR4 groups compared with that in the sham group. Ten days after I-R, 1 week of treadmill exercise significantly (P < .05) increased the latency in the PAT in the TR1 and TR4 groups compared with that in the SD4 group (Figure 1). At 31 days postischemia, 4 weeks of treadmill exercise significantly increased (P < .05) the latency in the PAT in the TR4 group compared with that in the SD4 and TR1 groups; however, the latency in the PAT in the TR1 group was not significantly different from that of the SD4 group (Figure 1).
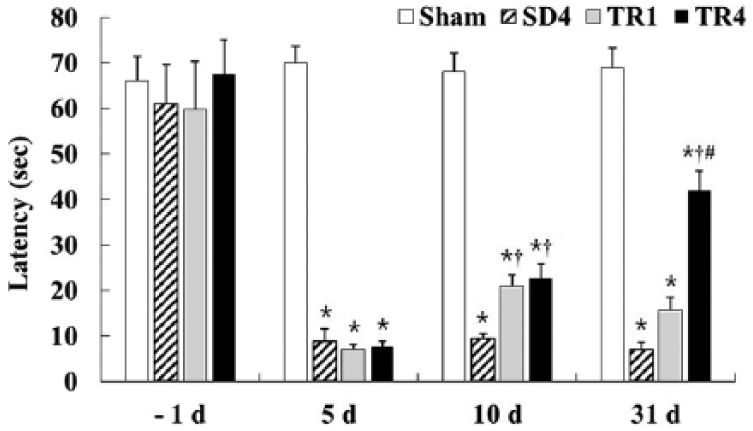
The effect of treadmill exercise on the latency in the passive avoidance test in the sham, SD4, TR1, and TR4 groups (n = 14 per group; *P < .05, significantly different from the sham group;
Neuronal Damage
NeuN Immunoreactive Neurons
Sham group
NeuN immunoreactive neurons were found in specific hippocampal subregions including the pyramidal and granule cell layers of the sham group (Figure 2A1-A4).

NeuN immunohistochemistry in the hippocampus of the sham (A), ischemia (B and C), and TR (D and E) groups. A few NeuN immunoreactive neurons are observed in the stratum pyramidale (SP; asterisks) of the CA1 region in the ischemia and TR groups, and these groups show a decrease in the number of NeuN immunoreactive neurons in the polymorphic layer (PoL; asterisks) of the dentate gyrus (DG). GCL, granule cell layer; MoL, molecular layer; SO, stratum oriens; SR, stratum radiatum. Scale bars = 400 µm (A1-E1), 40 µm (A2-E2), and 100 µm (A3-E4). (F) The mean number of NeuN immunoreactive neurons in the sham, ischemia, and TR groups (n = 7 per group; *P < .05, significantly different from the sham group). The bars indicate the means ± SEMs.
Ischemia group
The number of NeuN immunoreactive neurons was significantly decreased (P < .05) in the stratum pyramidale of the CA1 region and in the polymorphic layer of the DG, but not in the other subregions, 5 days after I-R (Figure 2B1-B4 and F). In the SD4 group, the number of NeuN immunoreactive neurons in the stratum pyramidale of the CA1 region and in the polymorphic layer of the DG was similar to that in the ischemia group 5 days postischemia (Figure 2C1-C4 and F).
TR group
In both the TR1 and TR4 groups, the distribution pattern of NeuN immunoreactive neurons in the hippocampus was very similar to that in the SD4 group. No significant difference was found between the TR1 and TR4 groups (Figure 2D1-D4, E1-E4, and F).
Endogenous Neurogenesis
DCX and BrdU Immunoreactive Cells
Sham group
A few DCX immunoreactive cells (neuroblasts) were seen in the subgranular zone of the granule cell layer of the DG (Figure 3A1 and A2). In addition, a few BrdU immunoreactive cells were found in the granule cell layer of the DG (Figure 3A3).

Immunohistochemistry for DCX, BrdU, and double immunofluorescence staining for BrdU, NeuN, and merged images in the dentate gyrus (DG) of the sham (A), SD4 (B), TR1 (C), and TR4 (D) groups. In the sham group, a few DCX and BrdU immunoreactive cells (black arrows) are found in the DG. In the SD4 group, the number of DCX and BrdU immunoreactive cells are increased compared with the sham group. The number of DCX, BrdU immunoreactive cells and double-immunoreactive (BrdU/NeuN) cells (white arrows) are significantly increased in the TR groups, much more so in the TR4 group. GCL, granule cell layer; MoL, molecular layer; PoL, polymorphic layer. Scale bars = 100 µm (A1-D1 and A3-D3), 40 µm (A2-D2), and 30 µm (A4-D6). (E) Relative number expressed as a percentage of the number of BrdU and DCX immunoreactive cells (E1) and the mean number of BrdU/NeuN immunoreactive cells (E2) in the sham, SD4, TR1, and TR4 groups (n = 7 per group; *P < .05, significantly different from the sham group;
Ischemia group
In the SD4 group, the number of DCX immunoreactive neuroblasts was significantly increased (P < .05) in the subgranular zone compared with the sham group, and most of the DCX immunoreactive neuroblasts had well-stained processes that extended into the molecular layer (Figure 3B1, B2, and E1). We also found that the number of BrdU immunoreactive cells was significantly greater (P < .05) than in the sham group (Figure 3B3 and E1).
TR group
The number of DCX immunoreactive neuroblasts was significantly greater (P < .05) in the TR1 group than in the SD4 group, and the processes of the DCX immunoreactive cells in the TR1 group were much more complex than those in the SD4 group (Figure 3C1, C2, and E1). In the TR4 group, the number of DCX immunoreactive neuroblasts in the DG and their dendritic complexity were markedly increased (P < .05) compared to the SD4 and TR1 groups (Figure 3D1, D2, and E1). In addition, the number of BrdU immunoreactive cells was significantly increased (P < .05) in the TR4 group than in the SD4 and TR1 groups (Figure 3C3, D3, and E1).
BrdU/NeuN Immunoreactive Cells
To elucidate whether newly generated BrdU immunoreactive cells transformed into mature neurons, double immunofluorescence staining was performed at 31 days postischemia. In the sham group, a few BrdU+ cells colocalized with NeuN immunoreactive neurons in the subgranular layer (Figure 3A6 and E2). In the SD4, TR1, and TR4 groups, many BrdU+ cells colocalized with NeuN immunoreactive neurons in the granule cell layer; the highest number of BrdU/NeuN immunoreactive cells was found in the TR4 group (Figure 3B6-D6 and E2).
MBP Immunoreactive Nerve Fibers and OSP Immunoreactivity
Sham Group
MBP immunoreactive myelinated nerve fibers were present in all layers of the hippocampus, especially the strata radiatum and lacunosum-moleculare of the CA1 region and the polymorphic layer of the DG (Figure 4A1-A4). On the other hand, weak OSP immunoreactive dot-like processes were found in all layers of the hippocampus; in particular, OSP immunoreactivity was high in the stratum lacunosum-moleculare of the CA1 region and in the polymorphic layer of the DG (Figure 4A5-A8).

MBP (left column) and OSP (right column) immunohistochemistry in the hippocampus of the sham (A), SD4 (B), TR1 (C), and TR4 (D) groups. The density of MBP immunoreactive fibers (arrows) are reduced in the strata radiatum (SR) and lacunosum-moleculare (SLM) of the CA1 region, and in the granule cell (GCL) and polymorphic (PoL) layers of the dentate gyrus (DG) in the SD4 group, although OSP immunoreactive processes are increased in the SR and SLM of the CA1 region. In the TR groups, MBP immunoreactivity is significantly increased compared with the SD4 group; in the TR4 group, MBP immunoreactivity is significantly increased in the SR (arrows), SLM (asterisk), and GCL (asterisk) compared with the TR1 group. The OSP immunoreactivity is significantly increased in the TR groups compared with the SD4 group. MoL, molecular layer; SO, stratum oriens. Scale bars = 400 µm (A1-D1 and A5-D5) and 40 μm (A2-D4 and A6-D8). (E) Relative optical density expressed as a percentage of MBP (E1) and OSP (E2) immunoreactive fibers in the sham, SD4, TR1, and TR4 groups (n = 7 per group; *P < .05, significantly different from the sham group;
Ischemia Group
In the SD4 group, the density of MBP immunoreactive fibers was significantly decreased (P < .05) in the strata radiatum and lacunosum-moleculare of the CA1 region, and in the granule cell and polymorphic layers of the DG compared to the sham group (Figure 4B1-B4 and E1). On the other hand, the density of OSP immunoreactive processes was significantly increased (P < .05) in the strata radiatum and lacunosum-moleculare of the CA1 region in the SD4 group, whereas OSP immunoreactivity was significantly decreased (P < .05) in the polymorphic layer of the DG (Figure 4B5-B8 and E2).
TR Group
The density of MBP immunoreactive fibers in the TR1 group was significantly increased in the strata radiatum and lacunosum-moleculare of the CA1 region compared with the SD4 group (Figure 4C1-C4 and E1). In the TR4 group, the density of MBP immunoreactive fibers was significantly increased (P < .05) in the strata radiatum and lacunosum-moleculare of the CA1 region and in the granule cell and polymorphic layers of the DG compared with the SD4 and TR1 groups (Figure 4D1 -D4 and E1). On the other hand, the density of OSP immunoreactive processes was increased (P < .05) in the strata radiatum and lacunosum moleculare of the CA1 region and in the polymorphic layer of the DG in the TR1 and TR4 groups, although there were no significant differences between the TR1 and TR4 groups (Figure 4C5-D8 and E2).
GLUT-1 Immunoreactive Microvessels and Protein Levels
Sham Group
GLUT-1 immunoreactive microvessels were observed in all layers of the CA1 region and DG (Figure 5A1 and A2).

GLUT-1 immunohistochemistry in the hippocampus of the sham (A), SD4 (B), TR1 (C), and TR4 (D) groups. The density of GLUT-1 immunoreactive microvessels is markedly decreased in the strata oriens (SO) and radiatum (SR) of the CA1 region, and in the polymorphic layer (PoL) of the dentate gyrus (DG) in the SD4 group. The density is significantly increased in the TR4 group (arrows) compared with the SD4 group. GCL, granule cell layer; MoL, molecular layer; SP, stratum pyramidale. Scale bar = 100 µm. (E) Relative optical density expressed as a percentage of GLUT-1 immunoreactive microvessel in the hippocampus in the sham, SD4, TR1, and TR groups (n = 7 per group; *P < .05, significantly different from the sham group;
Ischemia Group
In the SD4 group, the density of GLUT-1 immunoreactive microvessels was significantly decreased (P < .05) in the strata oriens and radiatum of the CA1 region and in the polymorphic layer of the DG compared with the sham group (Figure 5B1, B2, and E).
TR Group
The density of GLUT-1 immunoreactive microvessels in the strata oriens and radiatum and polymorphic layer was increased in all TR groups compared with the SD4 group; the density in the TR4 group was significantly higher (P < .05) than that in the TR1 group (Figure 5C1, C2, D1, D2, and E).
GLUT-1 Protein Levels
The level of GLUT-1 in the hippocampus of the SD4 group was lower than that in the sham group, and the levels in the TR1 and TR4 groups were higher than that in the SD4 group, although there were no significant differences between any of the groups (Figure 5F).
BDNF Immunoreactivity and Protein Levels
Sham Group
Weak BDNF immunoreactivity was observed in the pyramidal neurons of the CA1 region and in granule and polymorphic cells of the DG (Figure 6A1 and A2).
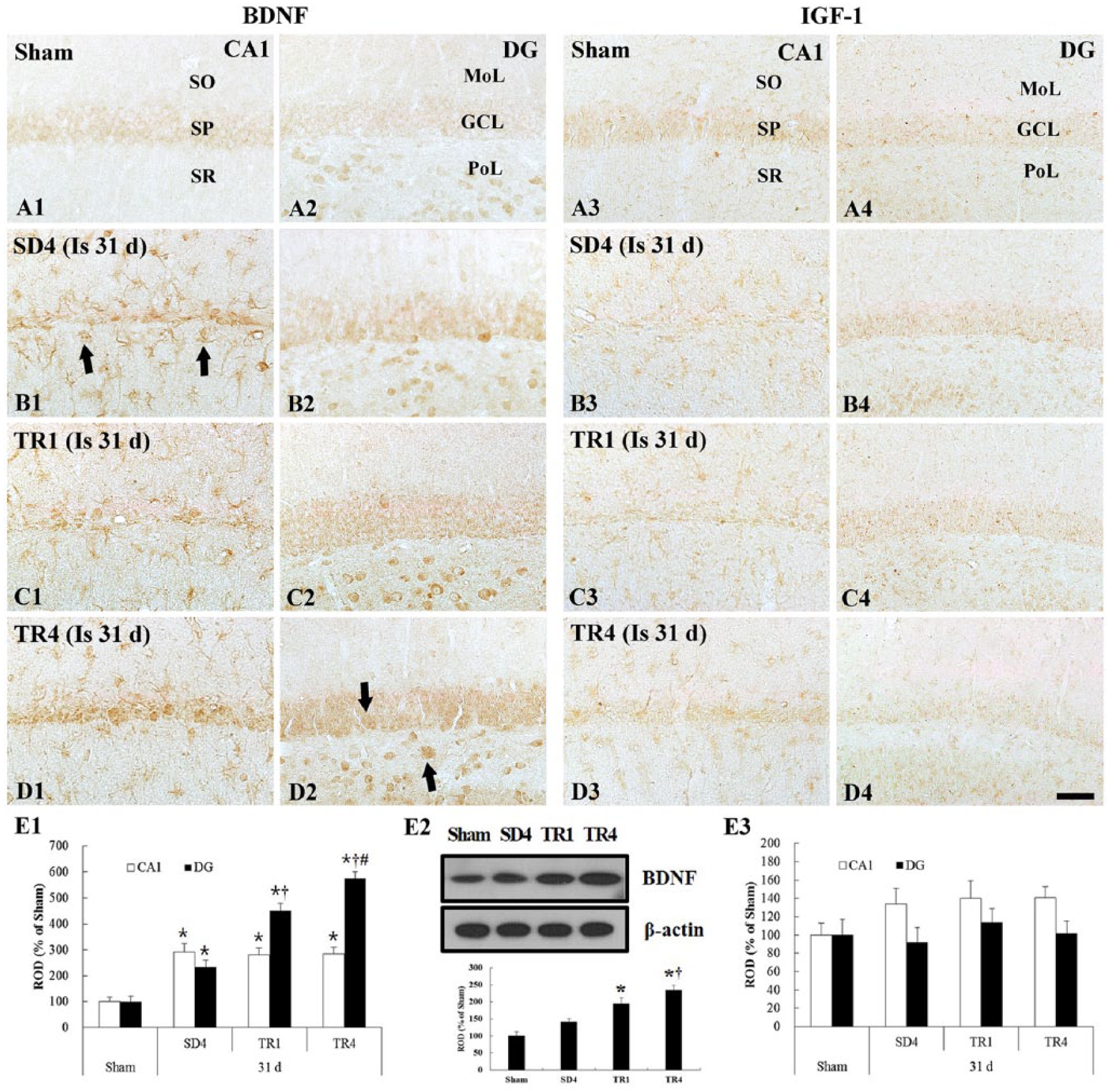
Immunohistochemistry for BDNF (left column) and IGF-1 (right column) in the hippocampus of the sham (A), SD4 (B), TR1 (C), and TR4 (D) groups. In the SD4 group, the BDNF immunoreactivity is strong and newly expressed in nonpyramidal cells (arrows) in the CA1 region. In the TR4 group, the density of BDNF immunoreactive nonpyramidal cells is similar to the SD4 group in the CA1 region and immunoreactivity in the dentate gyrus (DG) (arrows) is significantly increased compared with the SD4 group. The IGF-1 immunoreactivity is not significantly different in the SD4 and TR groups. GCL, granule cell layer; MoL, molecular layer; PoL, polymorphic layer; SO, stratum oriens; SP, stratum pyramidale; SR, stratum radiatum. Scale bar = 40 µm. (E) Relative optical density expressed as a percentage of BDNF (E1), IGF-1 (E3) immunoreactive structures, and western blot analysis of BDNF (E2) in the hippocampus in the sham, SD4, TR1, and TR4 groups (n = 7 per group; *P < .05, significantly different from the sham group;
Ischemia Group
In the SD4 group, BDNF immunoreactivity was strongly increased in the cells in all layers of the CA1 region and in the granule and polymorphic cells of the DG, and it was newly expressed in many nonpyramidal cells that showed strong immunoreactivity in the CA1 region (P < .05; Figure 6B1, B2, and E1).
TR Group
BDNF immunoreactivity was strong in the TR groups; the density of BDNF immunoreactive cells in the CA1 region was similar and the density in the DG was significantly increased (P < .05) compared with the SD4 group (Figure 6C1, C2, D1, D2, and E1).
BDNF Protein Levels
In the SD4 group, BDNF protein levels were slightly increased compared with those in the sham group; however, BDNF protein levels in the TR4 group were significantly increased (P < .05) compared with the SD4 and TR1 groups (Figure 6E2).
IGF-1 Immunoreactivity
Sham Group
IGF-1 immunoreactivity was found in pyramidal neurons of the CA1 region as well as in granule cells of the DG (Figure 6A3 and A4).
Ischemia Group
In the SD4 group, IGF-1 immunoreactivity was similar to that in the sham group, although the density of IGF-1 immunoreactive cells was slightly increased (Figure 6B3, B4, and E3).
TR Group
IGF-1 immunoreactivity and the density of IGF-1 immunoreactive cells in all TR groups were similar to those in the SD4 group (Figure 6C3, C4, D3, D4, and E3).
Discussion
In this study, we investigated the effects of long-term treadmill exercise on neuronal and memory recovery in the aged gerbil hippocampus following transient cerebral ischemia, and we investigated neurogenesis, repair of myelin microvessels, and changes in growth factors in order to understand the link between restorative exercise and neuronal and memory recovery.
In this study, the results of a PAT showed that short-term memory was significantly decreased after ischemia; however, short- and long-term treadmill exercise significantly improved memory function in the TR group. This result is consistent with those of previous studies that showed that wheel running and treadmill exercise restored impaired hippocampus-dependent memory function in adult mice or rats induced by focal cerebral ischemia.33,34 Therefore, it is likely that prolonged exercise facilitates recovery from ischemia-induced memory impairment.
It has been reported that 10-day or 4-week treadmill exercise started before neuronal death improved short-term memory by suppressing the ischemia-induced apoptosis of neurons in the CA1 region of the adult gerbil induced by transient cerebral ischemia.20,21 In addition, Li et al demonstrated that 3 weeks of pre-ischemic treadmill exercise significantly reduced neurologic deficits and infarct volume after focal cerebral ischemia in the adult rat. 35 Also, Bendel et al demonstrated that exercise-induced protection of neurons in the hippocampal CA1 region was closely related to recovery of learning and memory. 36 In the present study, however, we found that both short- and long-term treadmill exercise begun at 5 days postischemia, when neuronal death had occurred in the aged hippocampal CA1 region and DG, did not have any neuroprotective effect in the TR groups; the number of NeuN immunoreactive neurons was not significantly different in the CA1 region or the DG between the SD and TR groups. This finding indicates that treadmill exercise begun at a relatively late time point after transient cerebral ischemia cannot protect neurons in the aged hippocampus from ischemic damage.
On the other hand, we found that newly generated BrdU immunoreactive cells and DCX immunoreactive neuroblasts in the DG were significantly increased in the SD4 group. This finding is consistent with other studies that reported that stroke potently increased neurogenesis in the granule cell layer of the DG, and suggested that ischemia itself triggered neurogenesis in the granule cell layer of the DG.37 -39 In addition, treadmill exercise significantly increased the number of BrdU immunoreactive cells and DCX immunoreactive neuroblasts in a time-dependent manner in the granule cell layer of the DG in the TR group compared with the SD4 group, and furthermore, many BrdU immunoreactive cells were colocalized with NeuN immunoreactive mature neurons. Similarly, Briones et al reported that 2 weeks of treadmill training increased the number of newly proliferated cells that were colabeled with an immature neuronal marker, TuJ1, or a mature neuronal marker, NeuN, in the granule cell layer of the DG following transient global cerebral ischemia in adult rats. 40 Furthermore, some studies have suggested that mature neuroblasts formed from neuronal progenitor cells integrated into the existing DG network and increased plasticity 41 and that newly generated neurons in the DG may play an important role in the acquisition of new hippocampus-dependent memories.14,19 Based on our findings and those of previous studies, we suggest that prolonged exercise is closely related to an improvement in short-term memory via enhancement of cell proliferation, neuroblast differentiation, and neuronal maturation in the DG of the aged gerbil subjected to transient cerebral ischemia.
In the present study, memory function was restored despite neuronal death in the ischemic aged gerbil hippocampus, and we hypothesized that the improvement in memory caused by exercise may be related to adaptive plasticity, which can facilitate reconstruction of damaged neural circuits in the hippocampus after ischemia-reperfusion.
In the present study, the density of MBP immunoreactive myelinated fibers was markedly reduced in the CA1 region and DG in the SD4 group after ischemia, although the density of OSP immunoreactive processes was significantly increased. In addition, long-term exercise significantly increased numbers of myelinated fibers and OSP immunoreactive processes in a time-dependent manner in the TR4 group. This finding is supported by some studies that showed that focal cerebral ischemia induced oligodendrogenesis in peri-infarct gray and white matter, which is necessary to form myelin sheaths for sprouting axons during brain repair processes because existing mature or injured oligodendrocytes do not make new myelin sheaths.42,43 Moreover, Zatorre et al demonstrated that functional activity during learning stimulated the myelination of unmyelinated axons and the modification of myelinated axons during repair after myelin damage. 44 These results, along with our present findings, suggest that exercise may recover ischemia-induced damage of myelinated hippocampal nerve fibers.
In this study, hippocampal microvessels were examined using GLUT-1 immunohistochemistry, 45 because GLUT-1 is present in the endothelial cells of microvessels, and an increased number of GLUT-1 immunoreactive microvessels reflects an enhancement in functional angiogenesis. 46 We found that the density of GLUT-1 immunoreactive microvessels was significantly decreased in the SD4 group but significantly increased in the TR1 and TR4 groups. It is well known that ischemia itself stimulates angiogenesis in an effort to restore oxygen and nutrient supply to impaired tissues. 47 Many researchers have demonstrated that exercise enhances angiogenesis by improving cerebral vascular integrity and increases the levels of angiogenic growth factors, such as vascular endothelial growth factor and angiopoietin 1 and 2.14,46,48-50 Therefore, our data suggest that prolonged treadmill exercise after ischemia may facilitate neurovascular remodeling in the hippocampus of the aged gerbil.
On the other hand, although we did not examine the relationship between neurogenesis and angiogenesis in the present study, some researchers have suggested that neurogenesis and angiogenesis are closely related to one another because angiogenic vessels provide neurotrophic support to newly generated cells51,52; this process is regulated by growth factor cascades that could be an important mechanism mediating exercise-dependent benefits in memory recovery after ischemia. 53
Many studies have demonstrated that BDNF and IGF-1 signaling play key roles in guiding neuroblast differentiation and in improving motor and sensory deficits after cerebral ischemia54-56 and that physical exercise enhances memory via upregulation of hippocampal BDNF and IGF-1 levels.53,57-59 In the present study, we found that treadmill exercise significantly increased BDNF expression in the CA1 region and DG of the TR groups in a time-dependent manner, but exercise did not influence the expression of IGF-1, the level of which is significantly lower in the hippocampus of the aged gerbil than that in the adult gerbil. 60 Therefore, we conclude that long-term exercise could have a considerable effect on memory capacity by upregulating hippocampal BDNF levels, not IGF-1, in the aged hippocampus after transient ischemia.
In brief, this study shows that 4 weeks of treadmill exercise facilitated memory recovery despite neuronal death in the aged gerbil hippocampus subjected to transient cerebral ischemia and that long-term treadmill exercise enhanced cell proliferation, neuroblast differentiation, and neuronal maturation, and increased the density of myelinated nerve fibers, oligodendrocytes, and microvessels with an increase in BDNF expression. These results suggest that long-term exercise after ischemic stroke could restore ischemia-induced impaired memory function through the cumulative effects of multiple neurorestorative processes and that prolonged exercise could be an effective strategy for promoting neural plasticity after ischemic insult in the elderly.
Footnotes
Authors’ Note
Authors Ji Hyeon Ahn and Jung Hoon Choi contributed equally to this study.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was supported by the National Research Foundation of Korea (NRF) funded by the Ministry of Education, Science and Technology (2010-0010580), and by Basic Science Research Program through the National Research Foundation of Korea funded by the Ministry of Science, ICT and Future Planning (NRF-2013R1A2A2A01068190 and NRF-2014R1A1A3051721).