
Abstract
In this review, the movement characteristics of the human masticatory system are discussed from a biomechanical perspective. The discussion is based upon the three fundamental laws of mechanics applied to the various anatomical structures that are part of the masticatory system. An analysis of the forces and torques applied to the mandible by muscles, joints, articular capsules, and teeth is used to assess the determinants of jaw movement. The principle of relating the interplay of forces to the center of gravity of the lower jaw, in contrast to a hinge axis near its joints, is introduced. It is evident that the muscles are the dominant determinants of jaw movement. The contributions of the individual muscles to jaw movements can be derived from the orientation of their lines of action with respect to the center of gravity of the lower jaw. They cause the jaw to accelerate with six degrees of freedom. The ratio between linear and angular accelerations is subtly dependent on the mass and moments of inertia of the jaw, and the structures that are more or less rigidly attached to it. The effects of articular forces must be taken into account, especially if the joints are loaded asymmetrically. The muscles not only move the jaw but also maintain articular stability during midline movements. Passive structures, such as the ligaments, become dominant only when the jaw reaches its movement boundaries. These ligaments are assumed to prevent joint dislocation during non-midline movements.
Introduction
There is a large body of literature describing the dynamics of the human musculoskeletal system (for a review, see, for instance, Nigg and Herzog, 1999). In this large and mostly well-developed field, the human masticatory system occupies a relatively small place. One of the reasons for this underexposure is probably its relative complexity, which makes it more difficult to analyze than, for instance, the system of the shoulder, arm, hip, knee, or leg.
There are several reasons why masticatory dynamics are difficult to analyze. First, the masticatory system consists of a large number of muscles of various shapes and sizes, making it impossible to determine, unambiguously, how they might cooperate to perform a certain task (Wood, 1987; Lund, 1991; Hannam and McMillan, 1994). Second, they have a complex architecture (Schumacher, 1961; van Eijden et al., 1997), and their actions cannot be determined from their overall orientation only (van der Helm and Veenbaas, 1991). Third, the upper and lower jaws articulate through two very complexly shaped incongruent temporomandibular joints (Werner et al., 1991; Wish-Baratz et al., 1996). Any simplification of these joints based on concepts usually used for other joints (like hinges or balls-and-sockets) leads to considerable loss of functionality (van Loon et al., 1999). Furthermore, the articular surfaces are separated by a cartilaginous articular disc which is able to move more or less freely between these surfaces (Bade et al., 1994; Schmolke, 1994) and are influenced by, and affect, the movements of the jaw (Rees, 1954; Isberg and Westesson, 1998).
Apart from the intrinsic complexity of the system, there are several limitations to the collection of experimental data on masticatory function. For instance, some of the masticatory muscles run deep and are partially hidden behind bony structures, which prevents easy access for electromyographic (EMG) measurements (Wood et al., 1986; Koole et al., 1990; Murray et al., 1999a). Furthermore, many jaw movements are relatively small, posing stiff challenges to experimental systems designed to record relevant properties adequately (Naeije et al., 1996).
In summary, there are many factors that impede assessment of the mutual contributions of the relevant active and passive structures to jaw movements. Recently, the application of biomechanical models has provided an adequate experimental framework to explore masticatory dynamics without several of the drawbacks that accompany experiments with human subjects. They are powerful tools for establishing causal relationships in this field and have led to updates of or new formulations on various insights into the function of the masticatory system.
Generally, monographs on jaw movement (Griffin and Malor, 1974; Brown, 1975; Goodson and Johansen, 1975) have been written from a more clinical perspective, and have provided information mainly about the position, and positional changes, of the lower jaw. The dynamic aspects and their consequences were rarely taken into account. Recent developments in these areas have provided new insights. The present review is based upon a selection of studies on jaw movement analysis, from both clinical and basic perspectives. Its purpose is to establish an updated overview of the fundamentals of jaw movement, and it focuses on the contributions of muscles and the influence of passive constraints. It has been written from a biomechanical perspective and with an emphasis on masticatory dynamics. Since relevant topics such as neural control and feedback as well as properties of masticatory motor units have been reviewed relatively recently (Lund, 1991; Hannam and McMillan, 1994; van Eijden and Turkawski, 2001), they will not be discussed here.
In section 1, the relevant anatomical properties of the human masticatory system are reviewed briefly. They serve as a basis for all aspects of jaw movement and its determinants. Jaw movement is discussed in section 2, where its physics and its properties are reviewed. Interactions among anatomical structures and jaw movement properties are reviewed in section 3. An attempt is made to analyze the causal relationships between the two components. Finally, relevant issues that have not yet been resolved, but are assumed to be of critical importance for jaw movement analysis, are discussed in section 4.
(1) The Human Masticatory System
The human masticatory system consists of a mandible which is able to move in relationship to the skull and is guided by two temporomandibular joints through contractions of the masticatory muscles. To establish the contribution of each individual structure to jaw movements, one must explore the construction of the joints and the muscular system as well as the mechanical consequences of this construction. The morphology of the human masticatory system will be summarized very briefly. While there is a large quantity of literature in this area, the list of relevant citations in this article is far from complete.
Joints
Morphology
The anatomy of the temporomandibular joint has been described thoroughly (Rees, 1954). Mandibular movements are guided by its articular surfaces (Brown, 1975; Williams et al., 1995). These surfaces reside on the temporal bone of the skull, involving an articular eminence and a mandibular fossa, and on the roughly ovoid condylar head of the mandible. They are both irregularly shaped (Wish-Baratz et al., 1996), covered with fibrocartilage, and inaccessible for direct measurements in vivo.
The articular surfaces are separated by a cartilaginous articular disc with non-uniform thickness (Bade et al., 1994; Schmolke, 1994). This disc is able to move together with the mandibular condyle along the articular eminence while simultaneously rotating on the condyle (Rees, 1954). Disc movements generally run smoothly with respect to the articular surfaces.
The articular disc is connected superiorly to the temporal bone and inferiorly to the mandible by relatively loose fibrous structures. Together, these structures make up the articular capsule. It is reinforced laterally by the temporomandibular ligament, and is the only capsular structure that runs directly between the temporal bone and the mandible (Schmolke, 1994). Cadaver material reveals that the articular capsule is slack (Rees, 1954; Schumacher, 1983). There are two accessory ligaments: the sphenomandibular ligament, which runs medially from the mandibular ramus; and the stylomandibular ligament, which attaches to the mandibular angle from behind.
Mechanical consequences
The articular surfaces are highly incongruent, which means that the shapes of the upper and lower surfaces differ considerably. This allows for a large amount of motion at the cost of a lessened joint stability and relatively small areas of joint contact. The articular disc is supposed to reduce joint incongruency and increase joint stability by enlarging the contact area (Williams et al., 1995).
A second consequence of the incongruency of the joint, in combination with the slackness of its capsule, is that the movements in the joint are not restricted to rotations about more or less fixed joint axes, as in classic joints (Andrews and Hay, 1983). The condyle and temporal bone can be regarded as two separate bodies in space, usually held in appositional contact when the jaw moves. As a consequence, the mandible may be able to move with six degrees of freedom. Theoretically, it may rotate about an axis through, for instance, its incisor point. Therefore, the motion of this point per se bears no relationship to condylar motion. This property is known as kinematic redundancy. Furthermore, if the incisor moves from one point to another its path is not necessarily defined a priori. It may choose to move along a straight path or along a detour. In principle, the number of possible paths is infinite.
Muscular system
Morphology
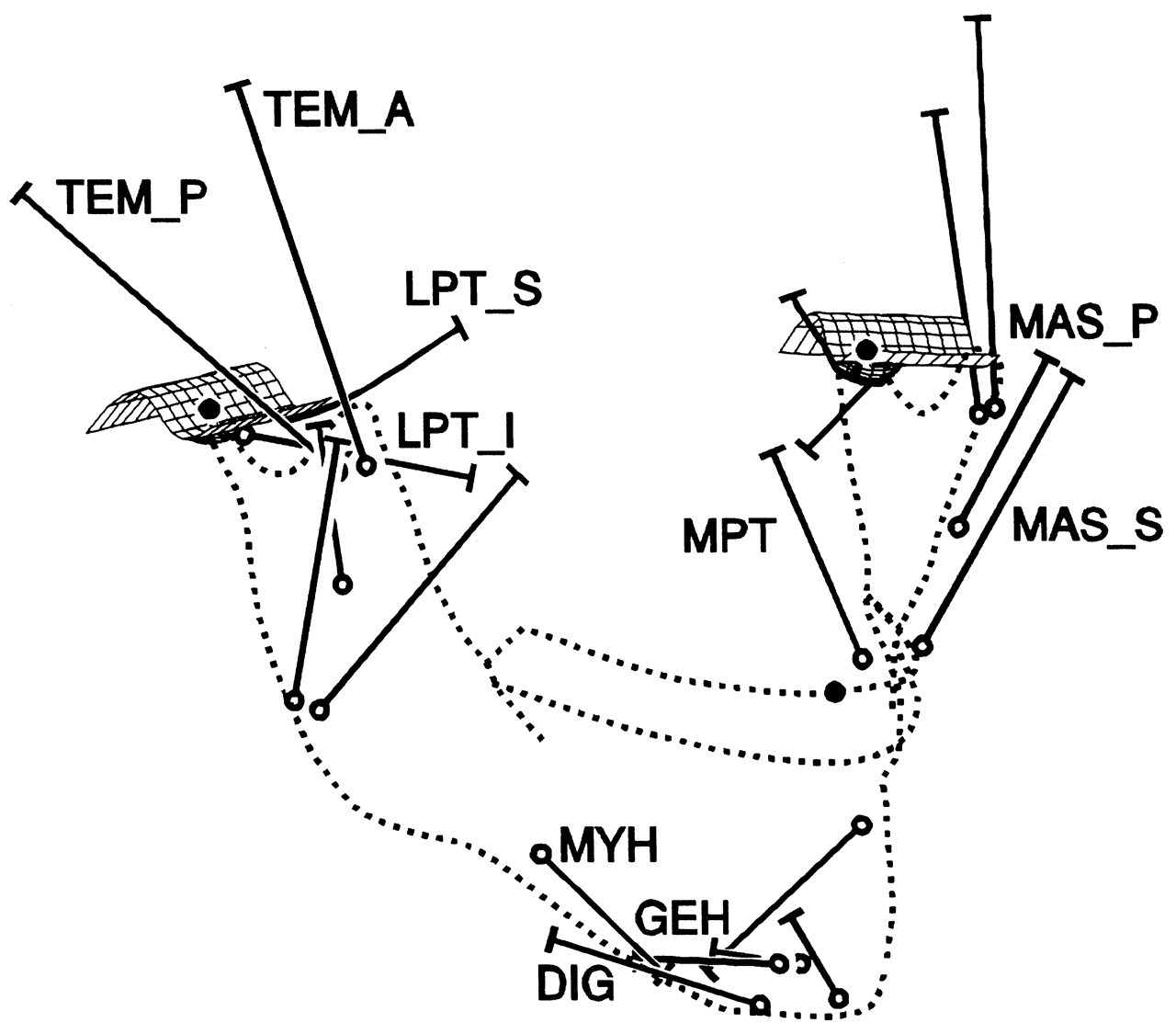
From a classic anatomical perspective, the masticatory muscles are divided into elevator and depressor groups. The elevator group consists of the masseter and temporalis muscles, which are located more or less superficially, and the medial pterygoid muscle, which is located more deeply. The muscles of the depressor group are located in the floor of the mouth. This group consists (from superior to inferior) of the geniohyoid, the mylohyoid, and digastric muscles. The geniohyoid and mylohyoid muscles connect the hyoid bone with the body of the mandible. The digastric muscle connects the mastoid process of the skull with the body of the mandible and is attached to the hyoid bone via a fibrous loop which runs around its intermediate tendon. The lateral pterygoid muscle completes the muscular system. It consists of a superior and inferior head running from the mandibular neck in forward and medial directions. Since both heads are considered to have different actions, they cannot be regarded exclusively as elevator or depressor (Juniper, 1981).
The elevator muscles are heavily pennate (Hannam and McMillan, 1994; van Eijden et al., 1996, 1997). They have relatively large physiological cross-sectional areas and are suitable for the generation of large forces. The fibers are short, which limits their capacity for active shortening during contraction. The depressor muscles and the lateral pterygoid have more or less parallel fibers and are therefore able to contract over a longer distance with less force.
Mechanical consequences
The human masticatory system contains more muscles than are apparently necessary to accomplish its tasks. This seems to be unnecessary from a mechanical perspective, but it must be noted that there are also spatial requirements to the construction of the muscular system. For instance, a muscular system that is mechanically optimal probably violates spatial requirements with respect to the adjacent airway and alimentary tract. The muscles can perform almost any task in various ways. Although the system is able to generate cyclic movements controlled by a central pattern generator (Lund, 1991; Ottenhoff et al., 1993), its muscles cannot be lumped into a limited number of alternating muscle groups. One of the reasons for this is that they have to adapt constantly to the texture of the food between the teeth (Thexton, 1992). The system is mechanically redundant, which means that there is an infinite number of muscle contraction patterns which can cause the same movement.
It has been demonstrated that various masticatory muscles have the capacity to deploy regionally different portions for different tasks (Møller, 1966; Wood, 1986; Miller, 1991; Blanksma and van Eijden, 1995; Blanksma et al., 1997; Murray et al., 1999b). Such functional heterogeneity, in combination with a relatively large attachment area, may cause the direction of the line of action of such a muscle to vary as well (van der Helm and Veenbaas, 1991). While there is no a priori evidence of co-activation between different muscle portions, the system is capable of fine-tuning, to a large extent, the orientation of the required muscle force (van Eijden et al., 1988) by selective activation of motor units (for reviews, see Hannam and McMillan, 1994; van Eijden and Turkawski, 2001). The relatively large extensive nature of some muscles may also cause spatially distant fibers within a muscle to shorten to various degrees during mandibular movements (van Eijden and Raadsheer, 1992; Hannam and McMillan, 1994; van Eijden et al., 1996, 1997). This may cause shifts in muscle lines of action which are not caused by the nervous system.
The depressors are directly or indirectly attached to the hyoid bone. When this bone moves downward through action of the infrahyoid muscles during wide jaw-opening (Muto and Kanazawa, 1994), the jaw depressors are stretched, which, in turn, lengthens their possibilities for active shortening. This may help in obtaining wider jaw gapes.
(2) Jaw Movement Basics
Degrees of freedom for jaw movement
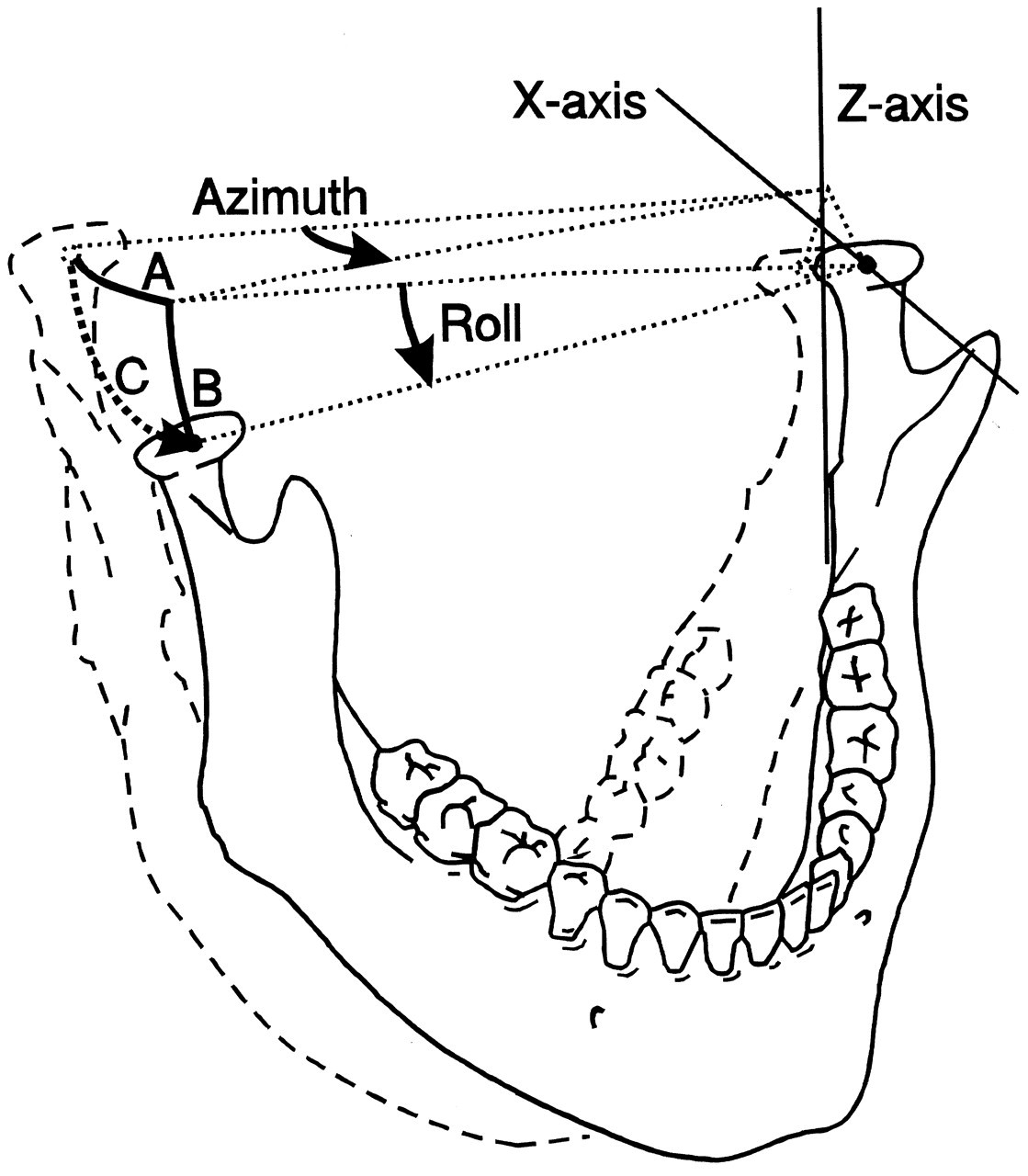
In three-dimensional space, a body able to move freely may perform translations and/or rotations. This applies to the lower jaw, although the degrees of the various movements are limited. Translations can be performed along, and rotations about, three independent axes. The translation axes and the rotation axes are not necessarily the same, but usually the three orthogonal axes of some Cartesian systems aligned to anatomical planes are used for this purpose. Translations can be described along axes which are, for instance, anteroposterior or X, mediolateral or Y, and supero-inferior or Z. Rotations can be defined by terms such as azimuth (about the Z-axis), elevation (about the Y-axis), and roll (about the X-axis) (Fig. 1) or yaw, pitch, and roll (Baragar and Osborn, 1984). It must be noted that there are many other conventions about sets of axes which are applicable. Independent of the applied set of axes, every movement can be expressed by a unique combination of the six independent fundamental movements (which are known as the six degrees of freedom of movement). The lower jaw does not move freely but is guided by its joints. These structures, therefore, may reduce the number of degrees of freedom (vide infra). Although translations and rotations relative to any of the three independent axes remain possible, they are no longer independent. For instance, if the joints should restrict one degree of freedom, the movement is completely determined by the other five.
If the movement of a point is studied—for example, the incisor point of the lower jaw or a point representing the condyle—it should be recognized that a point does not have an orientation. Rotations about axes through this point are thus meaningless, and it should be recognized that movements accomplished by rotations about distant axes can also be performed by translations. A point, therefore, can move with, at most, three degrees of freedom. The consequence is that the movement of any point on the jaw can be reconstructed from the movement of the jaw, but not the other way around.
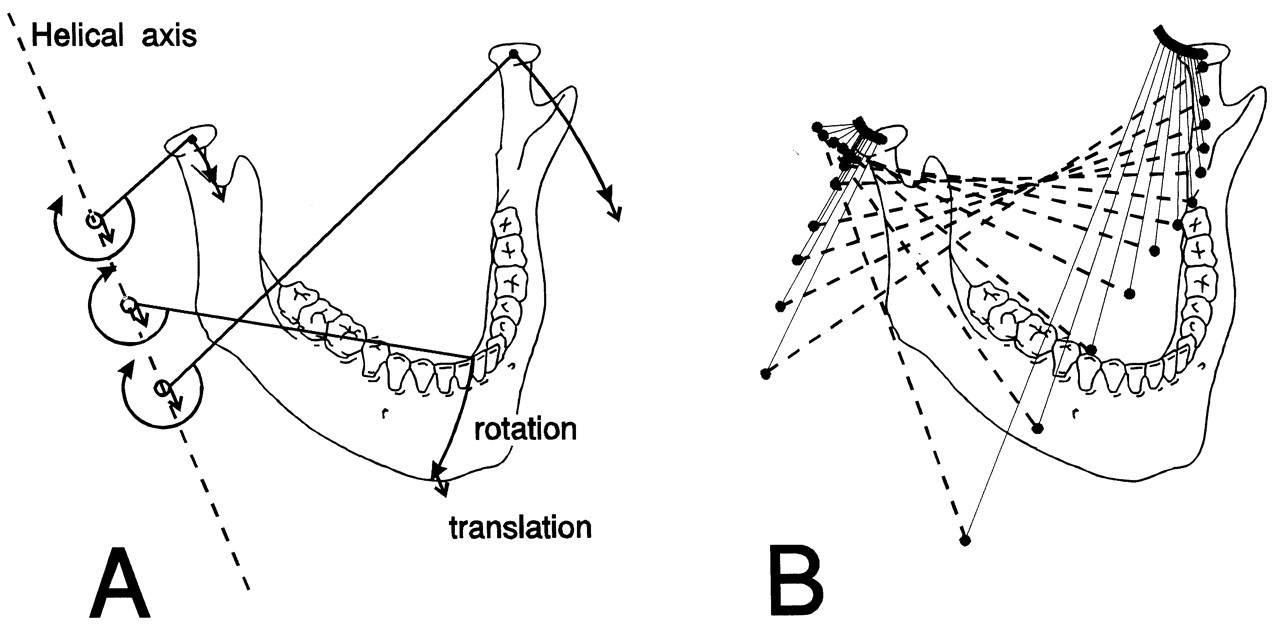
It is not easy to recognize functional aspects of jaw movement from a combination of translations and rotations with respect to pre-defined axes. An alternative way to describe a movement is by a rotation about and a translation along a so-called helical axis or screw axis (Kinzel et al., 1972). A movement can be described by subsequent (six degrees of freedom) small displacements. The orientation and location of the helical axis related to such a displacement (Fig. 2) provide information as to how it took place, while the moving body translates along and rotates about this axis. Generally, the helical axis is not stationary and may itself undergo displacements during movement. Therefore, each instant of movement is connected to a unique instantaneous helical axis. For sagittal movements, this axis is directly analogous to the instantaneous center of rotation for plane motion (Wu et al., 1988; Lindauer et al., 1995; Chen, 1998). It should be noted that the location and inclination of the helical axis, and the amounts of rotation about and translation along this axis, contain six independent variables according to the six degrees of freedom for movement.
Instantaneous helical axes thus provide a complete overview of jaw movement (Gallo et al., 1997, 2000; Koolstra and van Eijden, 1997b; Chen and Katona, 1999; Gal et al., 2000). The movement of teeth and condyles can be derived from them (Fig. 2). The relative contributions of rotations and translations of the mandibular condyle, for instance, can be determined from the distance to the axis. If, at a certain instant, the condylar movement is characterized primarily by a rotation, the helical axis will be situated close to the joint. If, in contrast, the translation component is dominant, the helical axis will be located at a distant location. These differences were demonstrated for jaw-opening movements performed with different muscle recruitment patterns showing no clear visual differences from normal movements in terms of displacement of teeth and condyles, but great differences in terms of helical axis positions (Koolstra and van Eijden, 1997b). This emphasizes that, from a clinical perspective as well, this approach can be relevant, for example, to an enhanced possibility of discrimination among different translations of the same type of movement by quantification of the inclination and remoteness of the helical axis. Furthermore, it enables on to discriminate between and among apparently similar movements caused by different muscle contraction patterns.
Physics of jaw movement : Newton ’s laws
The dynamics of a moving lower jaw are expressed by its position, its velocity, and its acceleration. According to the six degrees of freedom for movement, each of these three properties also consists of six independent variables. In a Cartesian system, the position is not defined only by the (X, Y, and Z) position of the center of gravity with respect to the origin of this system, but also by the orientation (azimuth, elevation, and roll) of the jaw. The velocity and the acceleration also have three linear and three angular components. For each of the six components, velocity is the (time) derivative of position and acceleration the derivative of velocity.
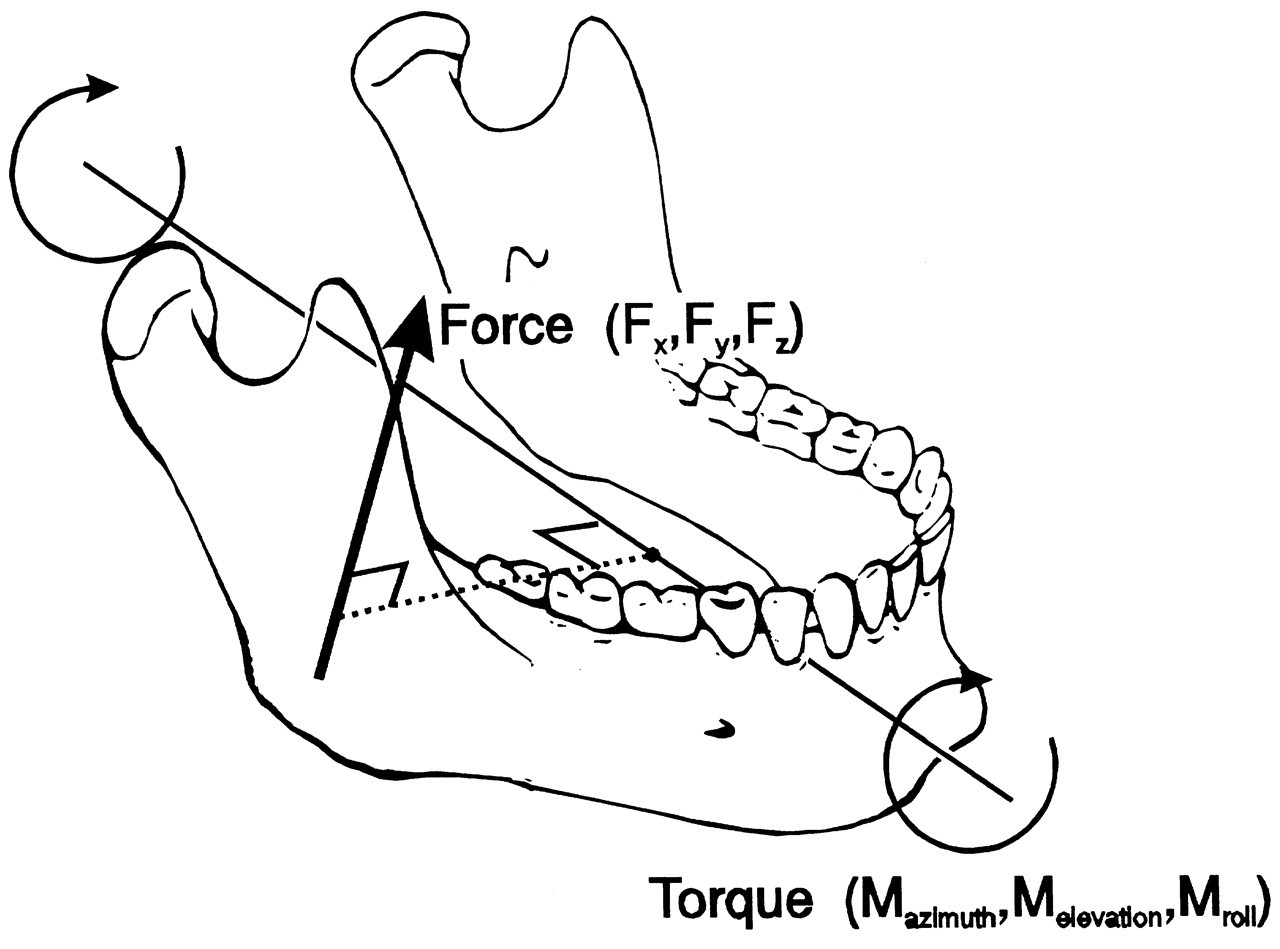
Every moving body, including the lower jaw, obeys Newton’s laws. Movements are caused by forces acting on the jaw. They may be active muscle forces and also passive (reaction) forces generated by joints, ligaments, and dental elements. The forces also have six components. Each linear force (Fx, Fy, Fz) is accompanied by a moment (angular) or torque (Mazimuth, Melevation, Mroll). The resultant forces and torques generate accelerations according to Newton’s second law (acceleration equals force divided by mass) (Fig. 1). This accounts for each degree of freedom, emphasizing the fact that the mass of the jaw also consists of three linear and three angular components. The three linear components of the mass of the lower jaw are equal to the common mass. The three angular masses (moments of inertia) are dependent on the distribution of mass about the axis under consideration and therefore on the shape of the lower jaw and adhering structures. The moment of inertia about an axis is defined as the sum of the mass of each particle multiplied by the distance between this particle and the relevant axis to the power of two (Nigg, 1999). For a lower jaw of about 0.44 kg, the moments of inertia have been estimated as 8.6 kg.cm2, 2.9 kg.cm2, and 6.1 kg.cm2 for Iazimuth (about the z-axis), Ielevation (about the y-axis), and Iroll (about the x-axis), respectively (Koolstra and van Eijden, 1995). This means that it requires about three times less muscle torque to accelerate the jaw for open-close movements than for latero-deviations. The accelerations cause changes in (linear and angular) velocity, and the velocities cause changes in (linear and angular) jaw position.
Influence of joints
The degrees of freedom of articulating bones are often reduced due to the construction of the connecting joint. An ideal ball-and-socket joint, for instance, does not allow for translations. Therefore, such a joint allows for movements with a maximum of three degrees of freedom. In contrast, the degrees of freedom in the temporomandibular joint are not reduced by its construction. While its articular capsule is relatively slack and its articulating surfaces incongruent, the mandibular condyle is able to move almost freely in the three-dimensional half-space bounded superiorly by the articular surface of the temporal bone. Articular contact is not necessarily maintained, although the distance along which the condyle is able to move perpendicular to the articular surfaces is relatively short. Furthermore, the articular surfaces are not rigid. The articular cartilage and the articular disc are deformable such that the distance between the bony surfaces will be proportional to the joint load (Huddleston Slater et al., 1999). Consequently, the mandible is also able to move with six degrees of freedom. If the joints are assumed to maintain articular contact all the time, and the joint contact is assumed to be rigid, a translation of the condyle in a direction perpendicular to the articular surface of the temporal bone is restricted, and the number of degrees of freedom for condylar movement is reduced to five. Furthermore, if both joints are assumed to be connected rigidly through the mandibular symphysis, the rotation of the lower jaw about an antero-posterior axis is restricted. In this (simplified) situation, it is able to move with four degrees of freedom (Schumacher, 1961).
Influence of muscles
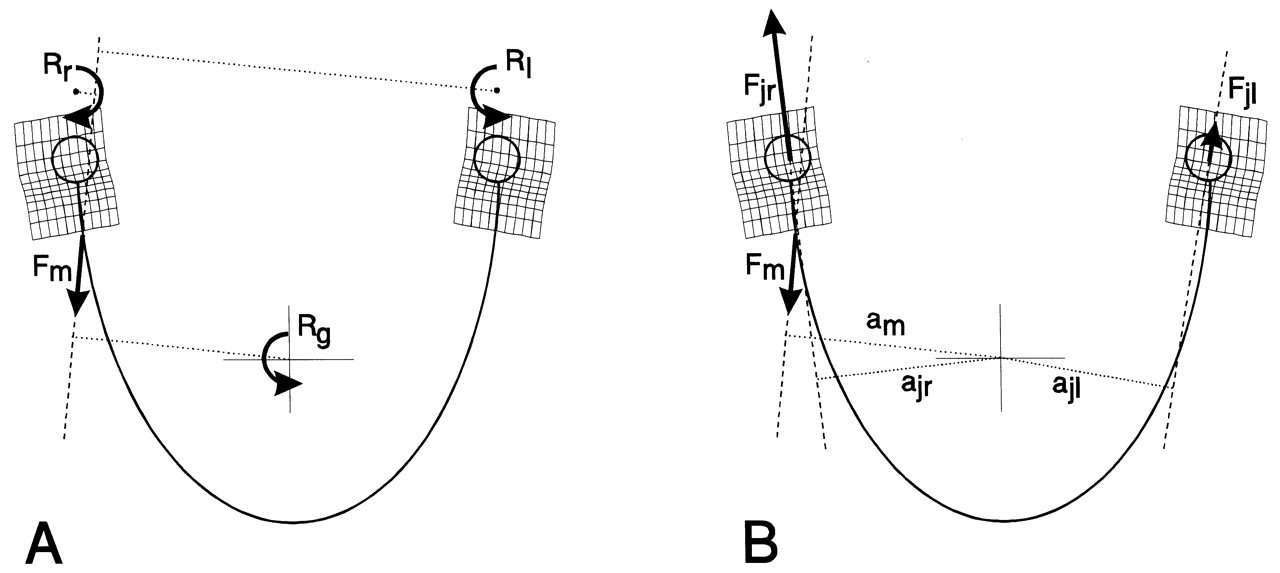
The jaw moves through contractions of the masticatory muscles. Each muscle contraction is associated with a force which is expressed by three independent variables: its magnitude, its point of application, and its orientation. The latter two are determined by the muscle’s line of action, defined by the geometry of the system. Each muscle can produce a translation of the lower jaw along its line of action, and a rotation about an axis perpendicular to it and running through the jaw’s center of gravity (Stern, 1974; Koolstra and van Eijden, 1995), as illustrated in Fig. 3. The translation and rotation caused by a muscle are not independent, and they express only one degree of freedom. Therefore, if such a muscle is activated homogeneously, the nervous system is able to influence only one degree of freedom through the magnitude of its force. If the muscle can be activated heterogeneously, and is represented by more than one independent line of action, it can influence more than one degree of freedom. Conversely, if separate muscles or muscle portions cannot be activated independently, then, together, they are able to influence only one degree of freedom. A system of muscles, therefore, is represented by a number of degrees of freedom equal to the number of independent lines of action. The masticatory system contains at least 20 muscle portions which may be activated independently (Fig. 4). The number of degrees of freedom of the muscular system, therefore, is considerably larger than the (maximum) six degrees of freedom of jaw movement. This causes a mechanical redundancy in the masticatory system.
(3) Determinants of Jaw Movement
Jaw movements caused by the masticatory muscles are guided by passive structures; thus, both passive and active elements generate forces and torques which accelerate the jaw (Fig. 5). Because of the large number of these forces, their changes during jaw movements, and their strong interdependency, it is difficult to separate these influences.
Active elements
Although passive structures in the masticatory system may act as constraints for jaw movements and guide the mandible along its path, active masticatory muscles are the prime movers in this system. Therefore, it can be expected that they will be the dominant determinants of jaw motion.
Muscle lines of action
While muscle lines of action differ considerably between muscles, each contributes to masticatory movements in a unique manner. Furthermore, the lines of action depend on the position of the lower jaw with respect to the skull. This causes continuous changes in the interplay of muscle forces and torques.
Investigators have estimated the lines of action of the masticatory muscles, in vitro, by connecting the centers of the attachment areas (centroids) on the skull and the mandible (Baron and Debussy, 1979). However, functionally different muscle portions that share attachment areas cannot be discriminated with this approach. This drawback can be overcome by measurement of the orientation of fiber bundles and incorporation of the influence of tendinous sheets (van Eijden et al., 1997). Furthermore, this method facilitates the estimation of fiber length as a function of jaw position (van Eijden et al., 1996; Koolstra and van Eijden, 1997b). The drawback of all in vitro methods is that results are not necessarily applicable to individual subjects. Therefore, muscle lines of action have been estimated in vivo by a determination of putative muscle attachment points (Goto et al., 1995, 2001) or by fitting the long axis through “Magnetic Resonance Imaging” sections (Koolstra et al., 1990). Although this enables individual characteristics to be incorporated, the estimates of the lines of action remain coarse.
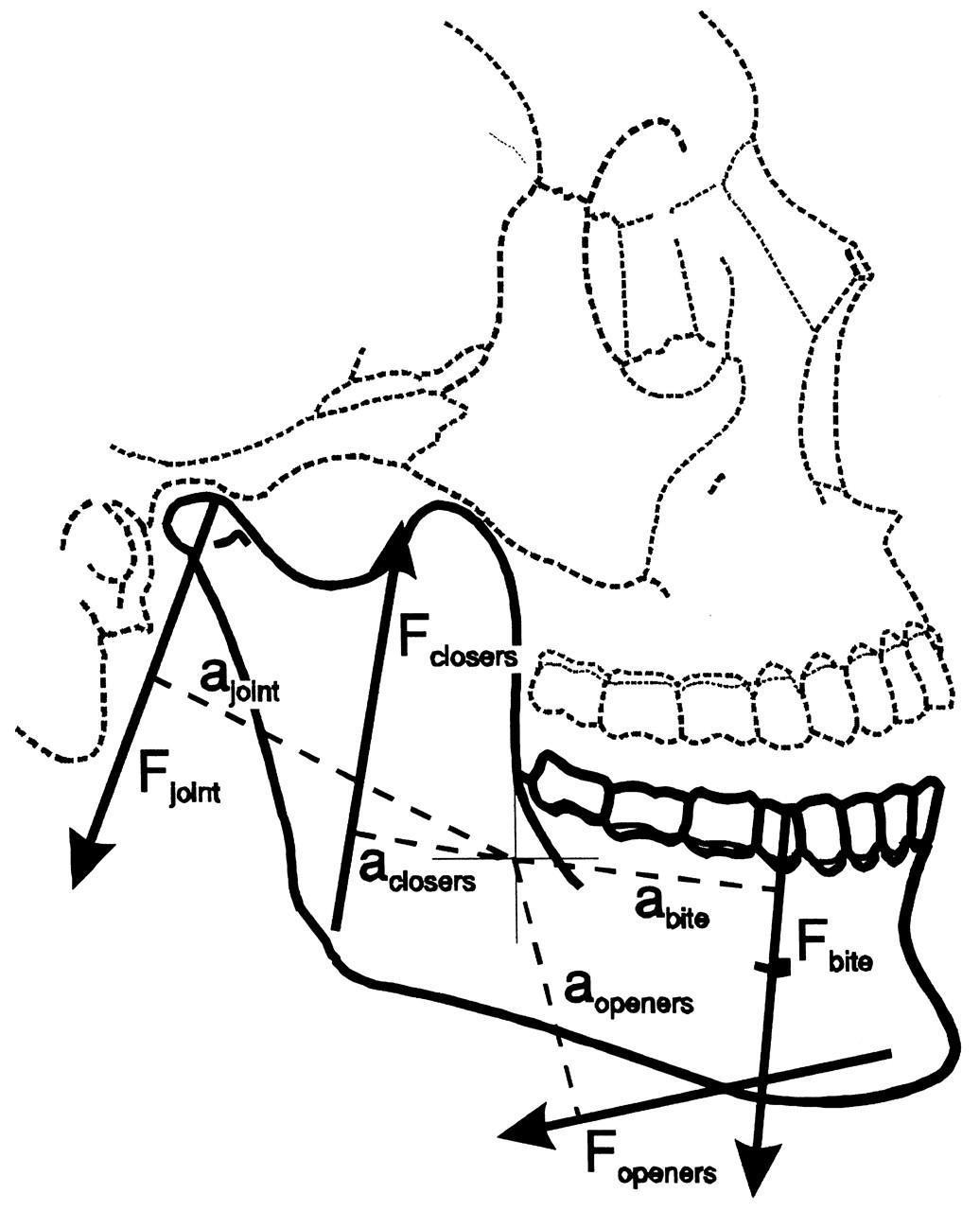
Generally, the contribution of a muscle to jaw movements can be established by the direction of its line of action and the position of this line with respect to the center of gravity of the lower jaw. It accelerates the jaw in the direction of the line of action according to:
In a sagittal plane analysis, the lines of action of most jaw-closers are directed upward, and those of the jaw-openers, downward and backward (Fig. 5). However, in both cases, each line of action has a similarly directed moment with respect to the sagittal axis through the center of gravity of the lower jaw. Jaw-closers and -openers are able to produce a similarly directed torque about this axis which leads to an angular acceleration in the “negative elevation” (opening) direction. Consequently, almost every muscle pair that is activated symmetrically attempts, aside from its specific action, to perform an opening rotation about the center of gravity. It is through this mechanism that both jaw-closers and -openers, despite their difference in orientation, are able to maintain articular contact while performing unloaded (symmetrical) jaw movements.
Muscle dynamics
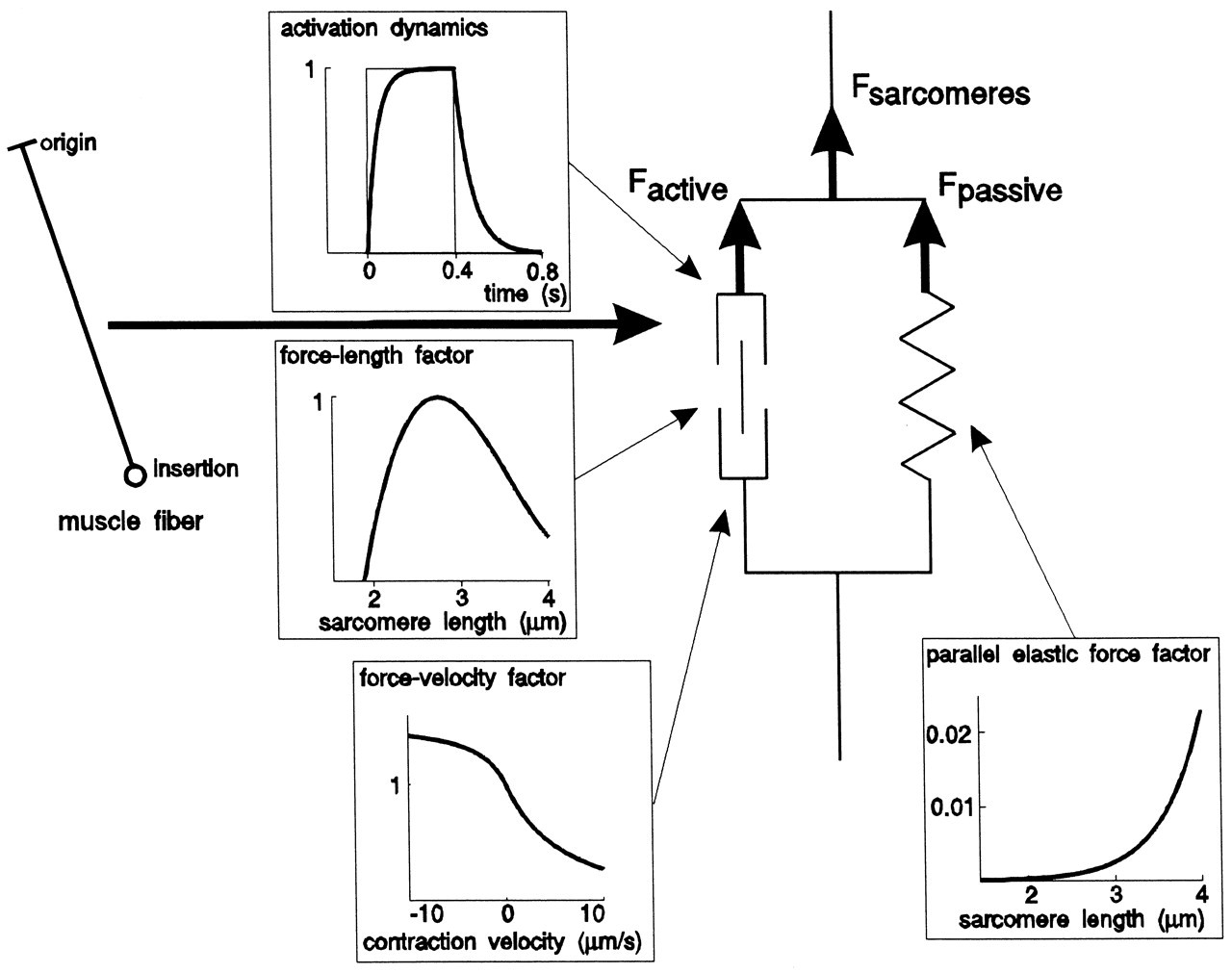
The optimum isometric force produced by a muscle Fopt is proportional to its physiological cross-section S (in cm2) and its activation A (in %), as denoted in the equation Fopt = 37 x S x A (Weijs and Hillen, 1985). Due to the dynamic muscle properties (Fig. 6), the instantaneous force of a concentrically contracting muscle is less than Fopt, and an eccentrically contracting muscle may produce an instantaneous force larger than Fopt. Due to these properties, the forces produced by masticatory muscles may change constantly during a movement, even though their activation levels remain constant (Koolstra and van Eijden, 1997a). The force-length relationship quantifies the property that enables a muscle to produce a force when its sarcomeres are not shortened below, or elongated beyond, certain lengths, and this property has been demonstrated to be an important limiting factor for masticatory muscle force development. For instance, the limited amount of maximum shortening of the fibers of the lateral pterygoid muscles prevent protrusion of the jaw beyond its normal limits (Koolstra and van Eijden, 1996). Furthermore, maximum jaw-opening is limited by the maximum shortening of the jaw-openers, which is counteracted by the passive forces of the elevators (Koolstra and van Eijden, 1997b; Langenbach and Hannam, 1999). The latter effect would be even more dramatic if the instantaneous center of rotation remained close to the joint (Weijs et al., 1989).
Muscle force is also dependent on the shortening velocity through the force-velocity relationship (Fig. 6). For jaw open-close movements, where all jaw-opening or jaw-closing muscles were activated simultaneously, it was demonstrated that the trajectory of movement is not very dependent on the speed of movement (Koolstra and van Eijden, 1997b). While this trajectory depends on mutual muscle forces, the force-velocity relationship does not considerably affect the mutual ratio of instantaneous muscle forces. Consequently, the possibilities for force production are affected similarly in all contributing muscles (Koolstra and van Eijden, 1997a). In contrast, the force-velocity relationship does appear to assist in deceleration of the lower jaw after a sudden disappearance of resistance during forceful biting (Slager et al., 1997). When such an event occurs, the force of the closing muscles can disappear instantly through the suddenly large shortening velocity, which may be considered profitable when the teeth are near occlusion, and there is little time to activate the jaw openers to decelerate the jaw.
Contribution of muscle action to jaw movements
Jaw movements are performed through co-contraction of various muscles. Electromyographic (EMG) measurement of the masticatory muscles during various jaw movement tasks (Carlsöö, 1952, 1956a,b; Møller, 1966; Wood, 1987), therefore, cannot be used to establish the individual contributions of the various muscles to a movement. For instance, it does not provide a means to decide whether two or more active muscles assist each other to perform a certain movement or work against each other to stabilize the system. EMG measurement has been used to detect functional heterogeneity in the activation of different muscle portions (e.g., Møller, 1966; Wood, 1986; Blanksma and van Eijden, 1990; Miller, 1991; Blanksma et al., 1992; Murray et al., 1999b). For example, it has been shown that the temporalis muscle shows a gradual heterogeneity in the antero-posterior direction, and that the masseter muscle can be subdivided functionally into a superficial, an anterior deep, and a posterior deep portion.
EMG registrations of masticatory muscles correlate muscle activity with jaw movements or static bite tasks. These correlations, however, do not necessarily reflect causal relationships. For causal relationships between muscle contraction and jaw movement to be established, the influence of the passive constraints must also be taken into account. Generally, this influence can be simplified to a screw displacement axis defined by the movement constraints in the joint (Andrews and Hay, 1983). The function of a muscle can then be defined by its moment with respect to this pre-determined axis. This concept was adopted by Grant (1973) and applied to the instantaneous center of rotation during jaw movements. Unfortunately, the method applies to joints influenced by only one degree of freedom by any muscle. Since the temporomandibular joint allows for movements with at least four degrees of freedom, this kind of analysis is irrelevant both for static situations (when the influence of the joint force is omitted) and for dynamic situations (when torques are expressed with respect to the center of gravity; Stern, 1974). In contrast, masticatory muscle function is not dependent on the location of the center of rotation. Conversely, the location of the instantaneous center of rotation (or instantaneous helical axis for non-midline movements) is dependent on the actions of the masticatory muscles. For instance, it has been demonstrated that jaw-open movements can be performed with different muscle recruitment patterns. Despite the relatively similar appearance of these movements, the trajectories of the instantaneous centers of rotation are very different (Koolstra and van Eijden, 1997b). If one wishes to estimate the work done by a muscle force during a movement, then its position with respect to the instantaneous helical axis is relevant (Gal et al., 2000).
A causal relationship between masticatory muscle contraction and jaw movement can be demonstrated experimentally only when muscles are activated independently. This is not an option in a regular experimental setting, nor is it likely even with intensive training, due to the central organization of motor control in the jaw muscles (Lund, 1991). An effective method, however, is registration of jaw movements evoked by electrical stimulation of isolated muscles. This can be done with in-dwelling electrodes (Zwijnenburg et al., 1996, 1999) or by electromagnetic stimulation of selected portions of the motor cortex (McMillan et al., 1998). The amplitude of such stimulation is necessarily restricted, to protect the experimental subject. Thus, the evoked jaw movements are small and require a very sensitive jaw-tracking device to be recorded. When the masticatory muscles are not fully relaxed, the other muscles may easily disturb the measurements. Even if complete rest can be obtained (for instance, in the unconscious state), the passive muscle forces may be relatively large compared with the evoked force and cannot be ignored. Consequently, direct muscle stimulation has not yet convincingly demonstrated the contributions of individual masticatory muscles to jaw movements.
Though direct experimental methods have failed to disclose the full functional potential of the masticatory muscles, biomechanical modeling approaches can help to develop reliable predictions on this subject. Such models are always simplifications; care must be taken that all relevant properties be included, and correct assumptions regarding model behavior must be made. In one model, for instance, it has been predicted that the jaw-opening muscles have the tendency to dislocate the temporomandibular joint (Osborn, 1993), while in another, it has been suggested that they have the tendency to stabilize the joint (Koolstra and van Eijden, 1997b). In the former model, torques were computed not with respect to the center of gravity (Stern, 1974) but with respect to the joint, thereby neglecting the law of conservation of angular momentum (Nigg and Herzog, 1999). In contrast, in the latter model, all relevant properties of (Newtonian) rigid body dynamics were implemented, strengthening the reliability of its predictions. According to these rigid body dynamics, the temporomandibular ligaments play an insignificant role during symmetrical jaw movements. It was demonstrated that activation of all jaw-closing muscles simultaneously leads to natural-looking jaw-closing movements, including a condylar movement similar to that observed experimentally (Koolstra and van Eijden, 1995). Conversely, activation of all jaw-opening muscles resulted in normal jaw-opening movements (Koolstra and van Eijden, 1997b). It is possible that the varying instantaneous center of rotation can be used as a means for the assessment of different muscle recruitment patterns during apparently similar jaw movements.
With the exception of the anterior temporalis and the superficial masseter, the principle that muscles, when activated unilaterally, generate a translation along their line of action and a rotation about the center of gravity has been confirmed. When their torque about the vertical axis through the center of gravity is considered, a contralateral latero-deviation would be expected, but instead, an ipsilateral latero-deviation occurs (Zwijnenburg et al., 1996, 1999). This apparent paradox can be resolved if one takes into account that joint loads caused by muscle forces also contribute to jaw movement (Fig. 7). Both the anterior temporalis and superficial masseter tend to tilt the contralateral condyle from the articular eminence because of a large ipsilateral joint load. The ipsilateral joint reaction force results in an ipsilaterally directed joint torque about the vertical axis through the center of gravity (Koolstra and van Eijden, 1999), which overcomes the muscle torque to cause an ipsilateral latero-deviation.
Passive structures
Passive structures contribute to jaw motion because they have the ability to resist its movements along one or more degrees of freedom. This resistance is expressed by the structure’s ability to generate a reaction force and/or torque. Furthermore, the jaw itself contributes to its movements through its inertial properties. The ratio between linear and angular accelerations effected by a muscle is subtly dependent on the mass and moments of inertia of the jaw, and all structures that are more or less rigidly attached to it. This attachment may include the part of the masticatory muscles attached to the mandible, the tongue, skin, and other soft tissues. It has been demonstrated, however, that the influence of these inertial properties on the final movement is small (Koolstra and van Eijden, 1995).
Articular surfaces
When the temporomandibular joints are loaded, reaction forces are transferred to the mandibular condyles. The cartilaginous structures in the joint have very low friction, and their deformation is fairly independent on loading direction (Beek et al., 2000), so it is probable that these reaction forces are directed perpendicular to the contacting articular surfaces. In a sagittal plane, they are directed inferiorly and pass posteriorly to the center of gravity of the lower jaw. These forces, therefore, have the tendency to lower the condyle and separate the articular surfaces (Fig. 5). The joint reaction forces also apply torques with respect to the center of gravity of the lower jaw. While the line of action of the reaction forces passes posteriorly to this center, the joint reaction torques lead to an angular acceleration about the sagittal axis through the center of gravity, which is bound to produce an elevation movement in the positive direction. Consequently, the reaction forces attempt to perform a closing jaw rotation about this axis (Koolstra and van Eijden, 1995).
The cartilaginous temporomandibular joint disc and the cartilage lining of the articular surfaces are deformable, with a finite and non-linear elasticity (Beek et al., 2001). This elasticity causes the joint reaction force to be dependent on the deformation. The more both bony articular surfaces move toward each other, the more the cartilaginous structures between these surfaces are compressed, and the larger the magnitude of the reaction force. Consequently, even if articular contact is maintained during static and dynamic situations, some movement perpendicular to the articular surfaces occurs. This is supported indirectly by the finding that the condylar path during an unloaded opening lies superior to the path during an unloaded closing movement (Yatabe et al., 1997; Huddleston Slater et al., 1999). The consequence of this observation is that joint loading during (unloaded) jaw opening is most likely larger than that during unloaded jaw closing. Furthermore, it supports the suggestion that the joints are loaded by the torque of the jaw-opening muscles (Koolstra and van Eijden, 1997b).
The movement range of the condyle is not limited to any great extent by the articular surfaces of the skull. Only when the joints are compressed in an upward or obliquely backward direction by manipulation can the fossa restrain jaw movements in the region of the condyles. The articular surfaces do not restrict protrusive and medio-lateral translations or rotations about any of the three axes. If the mandible performs a latero-deviation, the contralateral condyle has to move forward relative to the ipsilateral one, due to the interconnection of both condyles. During this movement, it is forced to move downward along the articular eminence. Consequently, this movement includes not only an azimuth rotation, but also a roll rotation (Fig. 8). While these rotations are interdependent, they nevertheless imply only one degree of freedom.
Articular capsule and ligaments
The bony parts of the temporomandibular joint are connected by an articular capsule composed of relatively loose collagenous fiber bundles organized in a more or less parallel fashion (Schmolke, 1994). On the lateral side, it is reinforced by a temporomandibular ligament. The fibers are able to withstand some stretching. They deform according to their (non-linear) elastic properties (Woo et al., 1993) and, in doing so, generate tensile forces. These forces may decelerate the attached condyle when it moves away from the articular surface of the temporal bone. A similar mechanical function can be attributed to the accessory ligaments, though these are very thin (Williams et al., 1995) and their function is probably negligible.
For quite a while, the temporomandibular ligaments have been considered a dominant constraint for condylar movement and, therefore, for jaw movement. This has been illustrated by the construction of more or less fixed axes for mandibular rotations, especially in the final phase of jaw closure, and the idea has been applied widely in the areas of prosthetic dentistry (Ramfjord and Ash, 1966) and temporomandibular dysfunction rehabilitation (Crawford, 1999). Presently, there is a considerable amount of evidence that does not support the presence of a fixed hinge axis (Lindauer et al., 1995; Chen and Katona, 1999) in normal functional movements. Another dominant role for the temporomandibular ligaments has been proposed for symmetrical jaw-opening movements. Here, it has been assumed that the temporomandibular ligament is always taut and forces the condyle to slide down the articular eminence (Osborn, 1993). However, if the temporomandibular ligament is taut, it cannot allow for habitual latero-deviation movements unless there is a mechanism for tightening the ligaments during midline movements and for slackening them during non-midline movements. It is very unlikely that such a mechanism exists. These considerations favor a less dominant role for the temporomandibular ligaments in controlling habitual jaw movements.
The temporomandibular ligament may play a dominant role in preventing the mandibular condyle from moving beyond the limits of the articular surface of the temporal bone. It has been shown that the masticatory muscles could pull the condyles a few mm beyond these limits if the temporomandibular ligaments were not present (Koolstra and van Eijden, 1999; Koolstra et al., 2001). Due to the interconnection of left and right condyles through the mandibular symphysis, the two ligaments are sufficient to limit both antero-posterior and medio-lateral condylar movements, during protrusive and retrusive mandibular movements, and during latero-deviations, respectively. Furthermore, when the mandible reaches these positions, the contracting masticatory muscles have almost reached the length where they become insufficient for force production (Koolstra and van Eijden, 1999). The forces applied to the ligaments will then probably not be large.
Teeth and food
The direct influence of teeth and food on jaw movements is due to the reaction forces occurring when the upper and lower teeth come into direct contact with each other or with a bolus of food in between. Through the interplay of muscle and joint forces, these reaction forces will be predominantly directed downward (Koolstra et al., 1988) and will be accompanied by an opening torque with respect to the center of gravity (Fig. 5), thus causing a joint loading. There is also an indirect influence, since the central nervous system is able to detect forces on the dental elements through mechanoreceptors in the periodontal ligament (Lund, 1991). This system, therefore, is able to adapt muscle activation as required by the presence of food. Furthermore, the nervous system is able to react even faster through reflexes.
The reaction forces are due to the resistance to deformation by the underlying structures. The dental elements are very hard, and should they come into contact with each other with some velocity, even a small deformation could result in a very large reaction force. Fortunately, the dentition is connected to the mandible via a deformable, collagenous periodontium. When a tooth of the lower dentition comes into contact with its upper neighbor with a certain velocity, it undergoes little deformation but is pushed into its socket. Due to the elasticity of the periodontium, a reaction force occurs which becomes larger as the tooth is pushed further into its socket. This reaction force acts on the mandible and causes deceleration until the movement stops. If food is compressed between the teeth, the reaction force increases more slowly, while the bolus itself is deformable.
The direct influence of the teeth on jaw movements is reflected by the superior portion of the Posselt envelope of incisal point motion, but the dentition can also have an indirect influence on jaw movements. It has been demonstrated that subjects with malocclusion have a more irregular chewing pattern than normally found (Gibbs et al., 1971; Lewin, 1985). It is not clear whether these aberrant patterns are due to tooth contacts themselves or to external factors.
During mastication, food is compressed and/or fractured between the dentition to reduce the particle size and facilitate swallowing. This compression and fracturing take place in the slow-closing phase of the masticatory cycle. It is apparent that the movement in this phase will be dependent on the mechanical properties of the food. For tough foods, the compression will be slower than for soft foods. Notably, the muscles are able to generate larger forces when contracting slowly. The peak velocity that follows the fracturing of hard, brittle food is much greater than that for soft food (Peyron et al., 1997). This peak velocity, however, is less than might be expected, possibly due to a decrease in muscle force as a consequence of the force-velocity relationship (Slager et al., 1997). Also, the size of the food affects mandibular movements, since the mandible has to be opened wider for larger pieces of food to be chewed. In the frontal plane, it has been observed that subjects chewing hard food tended to perform larger lateral excursions than when chewing soft food (Lewin, 1985).
Impact loads on the dentition may have consequences for the joints, since they transfer to the joints via the mandible. A healthy periodontium partially absorbs impact loads, and thus, it may prevent peak loads on the joints. This property does not exist if the dentition is connected with the mandible through an artificial implant. The mandible itself, however, is deformable (van Eijden, 2000), so it is possible that the transfer of impact loads of the teeth to the joints may be reduced by its elasticity. At present, there are no quantitative data on this subject.
Muscles
When inactive, the masticatory muscles generate passive forces which are dependent on the instantaneous length of their sarcomeres (Epstein and Herzog, 1998). When the sarcomeres are at or below optimum length, estimated at 2.73 μm (van Ruijven and Weijs, 1990), they are negligible, but increase exponentially if they are stretched beyond this length. Apart from these passive forces, muscle stretch can, indirectly, cause reflexes, because it is detected by muscle receptors (Lund, 1991).
The passive forces of the jaw-closing muscles are believed to decelerate the jaw at the end of jaw opening during mastication (Ostry and Flanagan, 1989) and become significant when the jaw nears its maximum opening (Koolstra and van Eijden, 1997a). It has been proposed that they are a determinant of maximum jaw opening (Langenbach and Hannam, 1999; Peck et al., 2000; Koolstra et al., 2001). Mathematical models applied to the study of the passive forces of the masticatory muscles have been unable to open the jaw more than about 3 cm, whereas an opening of 6 cm is frequently observed in vivo (Posselt, 1962; Brown, 1975). Therefore, the quantitative nature of these predictions is disputable. Due to the proposed exponential relationship between the passive muscle forces and their sarcomere lengths, small errors in the constants that determine this relationship may lead to relatively large errors in the projected passive forces. As long as there are no accurate quantitative data on the relationship between sarcomere length and passive force of the human masticatory muscles, this issue remains uncertain.
Interplay of passive and active structures
The most dominant determinants for jaw movement are the forces generated by active muscles. Passive forces may modulate jaw movements, but become dominant as the jaw reaches its movement boundaries. Axes of rotation of the jaw during free jaw movements are not primarily related to passive structures of the masticatory system, but are determined by muscle use. Nevertheless, in clinical practice, axes of rotation which were assumed to be connected to the temporomandibular joint have been measured and applied successfully for the diagnosis and treatment planning of masticatory dysfunction (Roth and Williams, 1996). This indicates that conclusions drawn from studies which were performed to search for practical solutions to a clinical problem cannot be automatically extrapolated to other problems in particular or to jaw movements in general.
The influence of the passive constraints appears to be more dominant as jaw movement deviates from the midline. Dynamic biomechanical analysis has demonstrated that the masticatory muscles are capable of maintaining the integrity of the masticatory system, in most cases, without the need for an articular capsule with ligaments to maintain articular apposition (Koolstra and van Eijden, 1997b). In contrast, they appear to play a role in reducing the medio-lateral movements of the mandibular condyle during latero-deviation (Koolstra and van Eijden, 1999). If the joints are loaded asymmetrically, the influence of their reaction forces on jaw movement has to be considered. When a muscle is activated unilaterally, the condylar reaction forces may produce a reverse movement compared with the one expected from the muscle’s line of action (vide ante). In practice, however, the muscles contract as groups rather than in isolation. For both midline and non-midline jaw movements, dynamic muscle properties should be taken into account, since they limit the force-producing capacities of the muscles, thereby restricting jaw movement possibilities.
(4) Final Remarks
Jaw movement analysis has evolved from early observation (Ulrich, 1896; Bennett, 1908) to experiments designed to formulate and validate or falsify testable hypotheses. In particular, the availability of dynamic biomechanical modeling methods has accelerated our understanding of jaw movements and the masticatory system (Koolstra and van Eijden, 1995, 1997b, 1999; Hannam et al., 1997; Langenbach and Hannam, 1999; Peck et al., 2000). It is now possible to predict the actions of the different muscles in this complex system by applying Newton’s laws. It has become clear that the masticatory muscles not only control jaw movements, but also maintain the physical integrity of the masticatory system. However, during relatively large medio-lateral excursions, the muscles may fail to keep the articular components in apposition, at which time the articular ligaments may be presumed to perform this role.
Although these developments have improved our knowledge of the working of the human masticatory system, the persistence of some outdated theories is striking. A prominent example is related to the articulation (Gysi, 1910), where there is a need to find a simple, reliable method for describing jaw movements near dental occlusion in anatomically different patients. Often, researchers and clinicians have attempted to use hinge axes to describe this movement. However, it has been known, since the end of the 19th century, that such axes are non-existent during habitual jaw movements (Ulrich, 1896). The concept of a hinge axis may have been revived by the demonstration that rotary jaw movement near occlusion can be accomplished through manipulation on cadaverous material (Rees, 1954). This concept has survived in clinical practice, where, despite a lack of a scientific basis (Mohl et al., 1990; Lindauer et al., 1995), it has been applied successfully for a long period (Roth and Williams, 1996).
While, for instance, with the use of dynamic biomechanical models, hypotheses regarding muscle actions in the functioning masticatory system are being validated (Koolstra and van Eijden, 1999), joint load predictions have not yet been verified satisfactorily. This is a consequence of the fact that direct measurement of temporomandibular joint loading without disturbing articular integrity has remained impossible. Unfortunately, this parameter is considered as a major influence on the development of wear and degeneration of the cartilaginous and bony structures of this joint. Insight on temporomandibular joint loading is, therefore, still limited to model predictions, and the reliability of these predictions is directly related to the assumptions and parameters built in such models. These include joint morphology and the material properties of its deformable structures that contribute to load distribution. Furthermore, muscle tensions applied during joint loading are required. In each subject, these parameters may be different, leading to the need for in vivo measurement. Progress has been made in reconstructing the relevant muscle lines of action (Koolstra et al., 1990, 1992) and bony parts of the joints (Krebs et al., 1994) in vivo, but, to date, no reliable method is available to create reconstructions of the cartilaginous tissues in the joints. For a complete overview of the applied muscle tensions to be acquired, their physiological cross-sections, architecture, and degree of activation must be estimated. Physiological cross-sections have been estimated from anatomic cross-sections (Weijs and Hillen, 1984; Koolstra and van Eijden, 1992) and muscle activation from EMG recordings (Carlsöö, 1952, 1956a,b; Møller, 1966; Lehr et al., 1971; Widmalm et al., 1988; Murray et al., 1999a,b), but it is questionable whether such methods can be applied routinely to all muscles involved. Masticatory muscle architecture has been studied in vitro (van Eijden et al., 1997), but the influence of individual variations on model predictions and the possibility of applying relevant corrections have not been established.
A start has been made on assessing the dynamic material properties of the cartilaginous structures in the human temporomandibular joint (Beek et al., 2001), but the nature and influence of individual variations are subject to speculation. Consequently, although much is known qualitatively, quantification of joint forces that incorporate individual variation still cannot be performed unambiguously.
The ultimate limiting factor for reliable masticatory function analysis incorporating biological variation is the lack of knowledge about masticatory muscle recruitment patterns. The mechanical redundancy of the masticatory system prevents their unambiguous prediction (Koolstra and van Eijden, 2001). The forces generated by the active muscles are the most dominant determinants of jaw movement and joint loading. The search for a rational way to predict muscle recruitment patterns remains a dominant challenge in the field of jaw movement analysis.
Six degrees of freedom for jaw movement. Dashed lines: principal axes. a: (linear) accelerations. F: (linear) forces. m: mass. α: angular accelerations. M: torques. I: moments of inertia. Helical axis. Force and torque generated by a muscle (arrow) with respect to the center of gravity of the lower jaw. Overview of the masticatory system. Ventro-lateral view. Continuous lines: muscle lines of action. Cross-bar: muscle origin. Circle: muscle insertion. MAS_S: superficial masseter. MAS_P: deep masseter. MPT: medial pterygoid. TEM_A: anterior temporalis. TEM_P: posterior temporalis. LPT_S: superior lateral pterygoid. LPT_I: inferior lateral pterygoid. DIG: digastric. GEH: geniohyoid. MYH: mylohyoid. Dots: position of centers of right and left condyle and incisor point. Forces acting on the lower jaw in the sagittal plane. Cross-hairs: center of gravity. Fclosers: mean force of the jaw-closing muscles. Fopeners: mean force of jaw-opening muscles. Fjoint: joint force. Fbite: bite force. a: moment arm of the different forces. Dynamic muscle properties. Total force is the sum of the forces produced by the sarcomeres (Fsarcomeres) The active force (Factive) is dependent on the activation through the activation dynamics, the instantaneous sarcomere length, and contraction velocity. The parallel elastic force (Fpassive) is dependent on the instantaneous sarcomere length. Schematic overview of the possible actions generated by force Fm of the superficial masseter or anterior temporalis muscle viewed in the horizontal plane. Rotations about vertical and horizontal axes during a latero-deviation movement.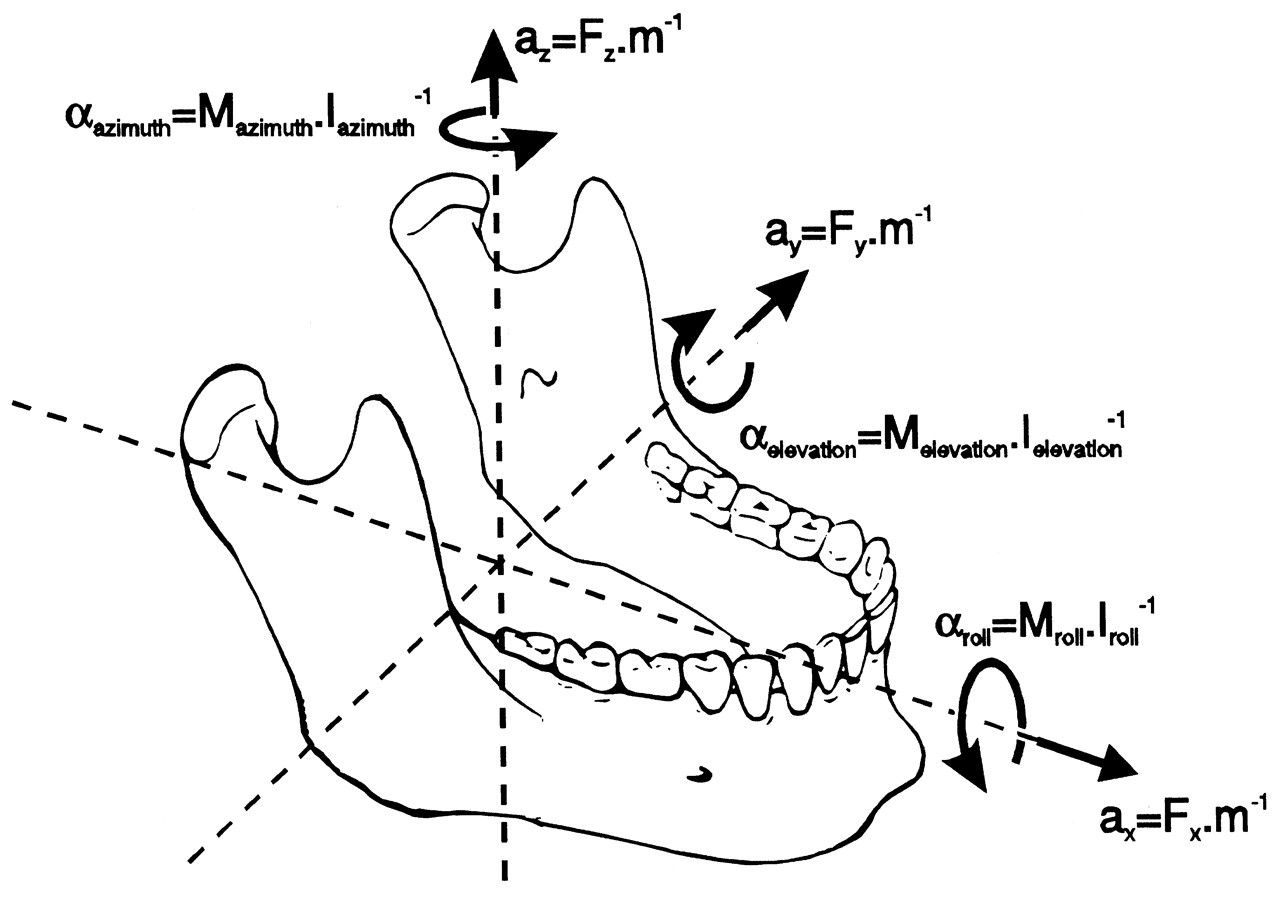
Footnotes
Acknowledgements
This research was supported by the Interuniversity Research School of Dentistry, through the Academic Center of Dentistry Amsterdam. I am grateful to Prof. T.M.G.J. van Eijden for his critical suggestions.