
Abstract
Sjögren's Syndrome (SS) is a chronic autoimmune disease characterized by histological and functional alterations of salivary and lacrimal glands that result in a severe dryness of the mouth and the eyes. The etiology of SS has remained undefined despite investigators' significant efforts to identify the mechanisms of initiation. Based on histopathology, several animal models are available—such as MRL/lpr, NZW/NZB, NFS/sld, graft vs. host, transgenic mouse expressing viral surface antigen, and the non-obese diabetic (NOD) mouse—for investigation of the etiology of SS. Biochemical and immunological similarities between human SS and autoimmune exocrinopathy (AEC) in the NOD mouse, including the loss of secretory function, establish the NOD mouse as an appropriate model to unravel the underlying pathophysiology of SS. Recently, several NOD congenic partner strains have been developed to investigate the roles of genetic intervals, cytokines, and autoantibodies in the disease pathogenesis. Studies on NOD-scid suggest that the pathogenesis of SS occurs in two phases: an asymptomatic phase, in which epithelial cells of exocrine tissues undergo dedifferentiation accompanied by elevated apoptosis; and a second phase in which autoaggression is mounted against target organ autoantigens, resulting in the activation of T- and B-cells, and the generation of autoantibodies. The presence of autoantibodies on the cell-surface signaling receptor, the muscarinic3 receptor, in both SS patients and the NOD mice correlates with the hallmark clinical symptom of secretory dysfunction. Additionally, the NOD mouse model provides an important example of how both Th1 and Th2 cytokines, as well as non-immune genetic loci, are involved in the maintenance of and progression to the overt disease state. Ultimately, analysis of these data provides insight into potentially novel therapeutic interventions.
(I) Introduction
Autoimmune diseases occur when self-tolerance breaks down and results in pathological destruction of one's own tissue. Whether the attack is against specific target organs, such as the thymus in Graves' disease or the pancreatic islets in type I diabetes, or against targets that are ubiquitous, such as connective tissues in systemic lupus erythematosus or rheumatoid arthritis, the tissue damage is mediated by effector functions of phagocytes, natural killer cells, T-cells, B-cells, and/or autoantibodies. Up to approximately 15% of America's population suffer from some form of autoimmune disease.
Sjögren's syndrome (SS) is a chronic autoimmune disorder in which patients present clinical symptoms of dry eyes and dry mouth due to inflammatory reactions against lacrimal and salivary glands, respectively. Dryness of the mucosal surfaces serviced by exocrine glands, such as skin, gastrointestinal tract, lungs, and vagina, has also been observed in patients with SS (Constantopoulos and Moutsopoulos, 1986; Tsianos et al., 1986). Involvement of the heart, blood vessels, muscles, kidneys, pancreas, liver, and brain, possibly due to epitope spreading, is also common in these patients. SS exists in two forms: primary SS, unassociated with other connective tissue autoimmune diseases; and secondary SS, accompanied by scleroderma, rheumatoid arthritis, or systemic lupus erythematosus (Fox and Kang, 1992).
Several classification criteria, summarized in Table 1, have been developed for the diagnosis of SS. These parameters include histological evaluation of lip biopsy for infiltrating leukocytes and the assessment of oral and ocular symptoms, as well as the presence of autoantibodies. Histopathological analysis of the minor salivary gland lip biopsy from patients usually exhibits lymphocytic infiltration, with the majority of lymphocytes being CD4+ T-cells. Infiltrates appear as peri-ductal and peri-vascular foci within the glandular architecture of the salivary and lacrimal glands. Focal scores classified as 1 or above, which consist of clusters of > 50 lymphocytes in a 4-mm2 area from the lower lip biopsy, are considered abnormal (Greenspan et al., 1974). Oral exocrine tissue is detected by the use of sialography for the image of one parotid gland, scintigraphy for the dynamic image of the function of all the major salivary glands, and/or the evaluation of stimulated saliva production. Recently, ultrasonography has been applied in place of salivary sialography or salivary scintigraphy (Salaffi et al., 2000). Ocular involvement is assessed by a Shimer's test to measure tear secretion and/or a Rose Bengal test to measure the ocular surface abrasion in patients. Serological evaluations identify the presence of rheumatoid factor, hypergammaglobulinemia, and specific anti-nuclear antibodies to nuclear protein, such as SS-A/Ro and SS-B/La (Fox and Kang, 1992). Antibodies against α-fodrin (Haneji et al., 1997), carbonic anhydrase II (Kino-Ohsaki et al., 1996), and acetylcholine muscarinic3 receptor (Bacman et al., 1996; Robinson et al., 1998d) have also been observed in the sera of SS patients.
(II) The Mouse as an Experimental Model
The pathogenesis of SS still remains to be determined, due to the mutifactorial nature of the disease, including genetic, hormonal, environmental, viral, and immunological factors (Fig.). In addition to the complexity of identifying etiological factors, the use of human subjects to study the etiology of SS has its limitations, due to ethical issues, dietary and environmental diversity, as well as the heterogeneous genetic backgrounds of human populations. These problems in the study of human disease have led to the development of animal models, especially rodents, exhibiting autoimmune exocrinopathy (AEC). Advantages of mouse models include their convenience in breeding, their short lifespan, their uniform genetic backgrounds, and the relative ease in manipulation of their environment. Although none of the mouse models that have been developed provides a complete profile of SS complications in humans, utilizing various mouse models for this autoimmune disease has given researchers insights into the disease pathogenesis of SS in humans.
Table 2 lists the various mouse models that have been studied. MRL/lpr, NZB/W (NZB/NZW F1 hybrid), NFS/sld, and NOD mice have been studied as the most notable animal models for their resemblance to autoimmune connective tissue diseases. All of these mice show immunological characteristics in common with SS (e.g., hypergammaglobulinemia, polyclonal B-cell activation, autoantibody production, and mononuclear leukocytic infiltration in the lacrimal and salivary glands). Like SS, the AEC of mice shows the majority of infiltrating cells to be CD4+ T-cells that express a pro-inflammatory cytokine profile. Despite heavy infiltrates within the glands, keratoconjunctivitis sicca (dry eyes) and xerostomia (dry mouth), which are the hallmarks of SS in humans, have not been observed in mouse models, with the exception of the NOD mice (Yamano et al., 1999).
(A) MRL/lpr strain
A defect in the lpr gene on chromosome 19, which encodes Fas protein, results in the accumulation of CD4-, CD8-, and CD3+ T-cells in the lymphoid tissue of MRL mice, due to the failure of lymphocytes to undergo programmed cell death. MRL mice without the lpr mutation (MRL/+) develop a late-onset autoimmune syndrome, indicating that the mutation accelerates rather than causes SLE-like autoimmune disease (Theofilopoulos and Dixon, 1985). In MRL/lpr mice, pro-inflammatory cytokines (Th1 profile) were detected in the spleen, lymph node, and submandibular glands by the polymerase chain-reaction (PCR) technique (Takahashi et al., 1996) and in situ hybridization (Mustafa et al., 1998). In the lacrimal gland, however, a Th2 profile predominated when assesed by immunohistochemistry (Jabs et al., 2000). This may suggest differences in the target organ microenvironment that guides immune cells to commit to be either Th1 vs. Th2 cytokine-releasing cells.
(B) NZB/W strain
NZB/W mice manifest AEC later than MRL/lpr mice and apparently have a longer lifespan. While the 2 strains have similar percentages of T-helper cells, NZB/W mice have a lower percentage of suppressor/cytotoxic and a greater percentage of B-cells compared with MRL/lpr mice (Jabs and Prendergast, 1988). In both strains, the lacrimal glands contain more B-cells in the infiltrate than in the salivary glands. Again, this indicates that the underlying pathogenesis may be different in different organs and in different strains, although they show similar phenotypes of autoimmune disease. MRL/lpr may be T-cell-dependent (Seaman et al., 1983), and NZB/W mice may be B-cell-driven (Ono et al., 2000).
(C) NFS/sld strain
NFS/sld mice bear an autosomal-recessive gene resulting in sublingual gland differentiation arrest. Salivary and lacrimal glands are infiltrated with autoreactive T-cells following neonatal thymectomy. The inflammatory infiltrates in the salivary and lacrimal glands consist mainly of CD3+ and CD4+ T-cells. Autoreactive CD4+ T-cells comprising a unique cytokine profile in the salivary glands exhibited high levels of interleukin (IL)-2, interferon (IFN)-γ, IL-10, and IL-12p40 mRNA (Hayashi et al., 1996). When the repertoire of TCR Vβ genes expressed within the inflammatory infiltrates was analyzed in NFS/sld mice with autoimmune lesions, a considerable preferential use of TCR Vβ8 was detected at the onset of disease (Haneji et al., 1994). A protein that has sequence identity to the human cytoskeletal protein α-fodrin was purified from these mice as a candidate autoantigen for SS (Haneji et al., 1997). Human patient sera also contain immune reactivity toward α-fodrin. One of the interesting findings of NFS/sld mice was an apparent loss of salivary secretory function observed at 18-20 months of age (Ishimaru et al., 2000). Reactivity to α-fodrin appears very early (1-2 weeks of age) with a corresponding elevation of apoptosis and immune infiltrates. Thus, the observation of secretory dysfunction in NFS/sld mice could be due to the age-related loss of exocrine function or tolerance rather than the direct consequence of the autoimmune disease, as proposed by this group.
(III) Non-obese Diabetic (NOD) Strain
NOD mice, a mouse strain that has been used for the study of type I diabetes (Makino et al., 1980), demonstrates a temporal lymphocytic infiltration of the exocrine tissues at 10-12 weeks of age that correlates with a loss of secretory function at 16 weeks of age. Since this model was suggested as the most appropriate animal model for the study of secondary SS (Humphreys-Beher et al., 1994), considerable work has been conducted to characterize unique features of NOD mice in association with disease pathogenesis. A predominance of CD4+ to CD8+ lymphocytes and a TCR Vβ repertoire dominated by Vβ6 and Vβ8 have been described in both the submandibular and lacrimal gland infiltrates. Analyses of cytokine mRNA expression in the submandibular glands favored an increase in several proinflammatory cytokines, including IL-1β, IL-6, IL-7, IL-10, IFN-γ, and TNF-α (Robinson et al., 1998a,b). Similarities between SS in human patients and autoimmune exocrinopathy in NOD mice are summarized in Table 3.
(A) Genetic control of autoimmunity in NOD mice and human patients
The NOD mouse genome is permissive for the development of type I diabetes due to the unique H2g7 major histocompatibility (MHC) haplotype (Leiter and Atkinson, 1998). Expression of I-Ag7 with no concomitant surface expression of an I-E molecule leads to differences in binding affinity and, subsequently, in antigen presentation. This complex locus on mouse chromosome 17 is estimated to contribute more than 40% of the relative risk for diabetes development. However, its congenic partner strain NOD.B10.H2 b, whose diabetogenic MHC locus has been replaced by the non-diabetogenic MHC locus of C57BL/10, shows no development of diabetes, yet continues to develop autoimmune exocrinopathy accompanied by loss of secretory function (Carnaud et al., 1992; Robinson et al., 1998c). This has led to the proposal that this mouse model is appropriate for primary SS. The association of certain HLA haplotypes, such as DR3 and DQ2, with SS has been reported in human patients (Arnett et al., 1988) and may suggest a difference in SS and AEC of NOD.B10.H2 b mice.
Other important genetic polymorphisms that distinguish NOD mice from other mouse strains include multiple mutations in the promoter regions for Fcγ receptor 1 (low affinity) and 2 (high affinity), a null mutation in hemolytic complement, polymorphisms in the IL2 gene, natural killer cell antigen 1, and prostaglandin synthase 2. These result in low expression of the Fcγ receptor in macrophage and impaired binding or clearance of certain IgG isotypes, lack of complement-dependent lysis due to lack of C5a component, less potent IL-2 cytokine, lack of functional NK1+ T-cells, and high constitutive expression of prostaglandin synthase 2 in NOD macrophage (Leiter and Atkinson, 1998). Different diabetes incidence rates between male and female mice in different mouse colonies indicate interactions of endocrine factors, environmental factors, and epistatic interactions of multiple genes that may be susceptible to or protective of the development of the disease.
To date, 19 susceptibility loci contributing to insulin-dependent diabetes (IDD) in NOD mice have been identified. Recent genetic studies (Brayer et al., 2001) with a C57BL/6 recombinant strain, containing the Idd5 genetic region derived from chromosome 1 of the autoimmune NOD mouse, showed that this mouse exhibited pathophysiological markers of autoimmune exocrinopathy, but not the loss of secretory function. This indicates that chromosome 1 controls biochemical changes and other loci which are necessary for immune infiltrates and the loss of saliva production. C57BL/6 carrying either the NOD-derived genetic interval on chromosome 3 encompassing Idd3, 10, 17, 18, or the Idd6 genetic intervals on chromosome 6 retained normal disease-free physiology. Conversely, NOD mice, containing chromosome regions of Idd5 and Idd3 derived from C57BL/10 and C57BL/6 mice, respectively, exhibited a reduced pathophysiology associated with SS-like autoimmune exocrinopathy. Analysis of these data suggests that genetic intervals on chromosomes 1 and 3 contain major determinants for the development of both IDD and SS-like disease in NOD mice. It will be interesting to see whether these two genetic intervals derived from NOD mice together in a normal healthy genetic background can duplicate the complete disease phenotype of SS-like autoimmune exocrinopathy. Genetic intervals in MRL/lpr mice (Nishihara et al., 1999) controlling sialadenitis were recently mapped to chromosomes 18, 10, 1, and 4, supporting the contribution of chromosome 1 as a major locus for the development of AEC.
As noted above, the major histocompatibility complex appears to play a critical role in determining an individual's susceptibility to the development of SS in human patients. Additionally, the frequencies of a new allele of the TAP2 (transport associated with antigen processing) gene, which had a unique substitution at codon 577, and GSTM1 null (Glutathione S-transferase M1) null genotype were higher in SS patients than in normal subjects (Kumagai et al., 1997, and Morinobu et al., 1999, respectively). In these patient populations, greater frequency of SS-A autoantibody was found compared with normal subjects. In the diabetic NOD model, a rare allele at the TAP1 locus has also been reported (Faustman, 1993).
The possible involvement of genetic loci encoding non-immune factors was first proposed in studies using the NOD-scid congenic mice (Robinson et al., 1996). Although the scid mutation prevents the spontaneous development of sialadenitis, insulitis, and diabetes in these mice, due to the absence of functional lymphocytes, several biochemical markers of differentiation (e.g., amylase or parotid secretory protein [PSP] expression) are diminished or aberrantly expressed and processed in exocrine glands of NOD mice. Amylase is one of the most abundant proteins in saliva, and in mice, PSP is a transiently expressed protein in the submandibular glands until 5 days of age in mice (Mikkelsen et al., 1992; Mirels and Ball, 1992; Ball et al., 1993). Changes in the expression of these molecules may indicate dedifferentiation in the glands of mice with the NOD genetic background. Elevated levels of caspase and matrix metalloproteinase (MMP) activity accompany morphological loss of submandibular gland acinar cell structures (Yamachika et al., 1998). Aberrant proteolytic processing may play a role in the generation of otherwise hidden cryptic antigens, thereby priming the immune system for an autoimmune response (Casiola-Rosen et al., 1995). Analysis of these data indicates the presence of underlying non-immune components associated with regulation of glandular homeostasis that may initially trigger the disease independently of immune cells, followed by loss of secretory function mediated by immune cell components. A recent report indicates that reactive oxygen species (ROS) can play a role in exposing cryptic epitopes of type IV collagen hidden within basement membranes and result in autoimmune Goodpasture syndrome (Kalluri et al., 2000). ROS could elicit apoptosis by induction of mitochondrial membrane permeability transition and release of cytochrome c (Lee and Wei, 2000).
The possible involvement of genetic factors encoding non-immune components has been investigated with the use of neonatal submandibular glands from NOD mice (Cha et al., 2001), to exclude immune responses and to explore the identification of early triggers for autoimmunity in disease-prone mice. Interestingly, cell proliferation rates examined by bromodeoxyuridine (BrdU) labeling and gelatinase activity assayed by degradation of collagen type IV were significantly reduced at the time of birth in the submandibular glands of NOD mice compared with normal mice. In contrast, there was elevated gelatinase activity in NOD submandibular glands around the time of weaning (21 days) compared with control mice. This enhanced MMP activity has also been detected in the adult onset of AEC in NOD mice. Extracellular matrix proteins, such as decorin and biglycan, were degraded by saliva and exocrine gland lysates from NOD mice and the congenic partner strains, NOD-scid and NOD.B10.H2 b, while no degradation was observed in control mice. This proteolytic activity for proteoglycans was inhibited by the MMP inhibitors, EDTA, GM6001, and 1,10-phenanthroline (Yamachika et al., 2000). Whether the disturbance in the cell proliferation in neonatal NOD mice is due to the intrinsic problems in the cell-cycle clock apparatus or to those in exogenous signals (e.g., growth factors, cytokines, or matrix-degrading enzymes) (Fig.) and how these changes affect the initiation of the disease remain to be answered.
Branching morphogenesis of the mouse submandibular gland (SMG) is dependent on cell-cell contact between and within epithelium and extracellular matrix. Such interactions are mediated by hormones, growth factors, cytokines delivering the signals into specific gene responses regulating cell division, apoptosis, and histodifferentiation (Jaskoll and Melnick, 1999). Specific cytokines can have multiple roles in regulating genes that encode for MMPs (Mengshol et al., 2000), genes that are involved in programmed cell death (Gannot et al., 1998), genes that influence neuronal responses (Soliven and Wang, 1995; Lu et al., 1997), and genes that modulate immune responses. Potentially, one of the interesting pathways controlling this response is the insulin-like growth factor (IGF) system and its binding proteins (IGFBP), where the genes are located on mouse chromosome 1. These molecules are involved in the maintenance of calcium homeostasis and protection from oxidative stress and apoptotic stimuli (D'Ercole et al., 1996; Dore et al., 1997) as well as the early development of salivary glands (Jaskoll and Melnick, 1999). When TNF-α and IFN-γ were applied in combination with IGF-I, these cytokines induced specific IGFBP synthesis which modulated human salivary gland (HSG) cell growth (Katz et al., 1995). Interestingly, patients with rheumatoid arthritis have lower concentrations of IGF-1 and IGF-2 than healthy controls (Fan et al., 1995; van de Loo et al., 1998). Due to the complex interactions of multiple genes and proteins, thorough screening of differentially expressed genes in NOD mice by means of microarray techniques may give us a list of possible candidate genes to be examined in human patients.
Altered glandular homeostasis in human SS patients has also been reported. Biopsies from SS patients showed significant increases in laminin protein and laminin mRNA expression preceding lymphocytic infiltration compared with normal controls. This study suggests that an alteration in basement membrane synthesis is an early event associated with salivary gland pathology in SS prior to the attack by lymphocytes (McArthur et al., 1997). Interestingly, IFN-γ was shown to up-regulate laminin and MMP-9 expression in the HSG cell line when the cells were treated with IFN-γ in vitro (Daniels et al., 1999; Wu et al., 1997, respectively). Elevation in MMP concentration in the saliva from patients with SS may be responsible for the increased remodeling of basement membrane proteins, such as laminin, and/or structural destruction of the basement membrane scaffolding in salivary glands in SS (Konttinen et al., 1998). Indeed, recent analysis of MMP-9 activity in gland biopsy material has provided evidence for the production of proteases in acinar cells of salivary glands removed from sites of lymphocytic foci (Pérez et al., 2000). Additionally, higher levels of MMP activity correlated with more severe histopathology.
(B) Immune cell infiltraton in the glands of NOD mice and human patients
The majority of infiltrating immune cells in submandibular glands of NOD mice are CD4+ T-cells, comprising 45% of the infiltrates. B-cells account for 20%, and CD8+ T-cells were 15% of the infiltrates. The percentage of B-cells is higher (30%) in the lacrimal glands than in the submandibular glands. Focal T-cells preferentially express Vβ6 and Vβ8 TCRs in these tissues (Robinson et al., 1998a,b). Adoptive transfer of splenic T-cells from NOD diseased mice into syngeneic healthy neonates can apparently transfer the histopathologic disease phenotype (Goillot et al., 1991). NOD.Igμnull mice, which lack B-cells, fail to lose secretory function, suggesting a direct role for the B-cell and/or autoantibodies in mediating clinical symptoms of exocrine dryness (Robinson et al., 1998d). Analysis of recent data shows that mature dendritic cells are detected by immunohistochemical methods in the NOD submandibular glands beginning at 5 weeks of age, suggesting the possibility of dendritic cells as the antigen-presenting cells for the disease (van Blokland et al., 2000). Interestingly, increased RNA expression of co-stimulatory molecules B7.1 and B7.2 on the ductal and acinar epithelial cells of salivary gland biopsy tissue from patients with SS was detected. Furthermore, IFN-γ is able to up-regulate B7.1, B7.2, and HLA-DR expression in salivary gland epithelial cell lines from SS patients (Manoussakis et al., 1999). This suggests that active participation of epithelial cells in the disease process occurs through the expression or up-regulation of co-stimulatory molecules for the presentation of antigen to the T-cell or through the up-regulation of apoptotic molecules such as Fas ligand (FasL) to induce apoptosis in the diseased glands.
(C) Role of apoptosis in the development of autoimmunity of NOD mice and human patients
Various ratios of apoptotic cells in the salivary glands of animal models and human minor salivary gland biopsy samples have been detected by the terminal UTP nucleotide end labeling method (TUNEL). Molecules generated by apoptotic processes are not immunogenic in normal situations. However, timing, localization, proteolytic processing, or the amount of apoptotic events at one point in time in the glands could certainly play a role in the pathogenesis by exposing cryptic antigens or altered antigens for presentation. It is also possible that a defect may exist in the clearance of apoptotic cells by phagocytic cells in the glands. A recent study indicates that germline mutations in Fas and FasL were not detected in adult patients with primary SS (Bertolo et al., 1999). Nevertheless, elevated glandular expression of the Fas-FasL and apoptosis has been detected in patients with SS in many studies, suggesting an involvement of Fas and FasL-mediated apoptosis in the pathogenesis of SS. The majority of infiltrating lymphocytes in SS are Fas+, bcl-2+, and Bax+, and they are resistant to apoptosis. The up-regulation of bcl-2 in lymphocytes of SS may inhibit apoptosis, leading to B-cell lymphoproliferation, since bcl-2 is known as an anti-apoptotic molecule by prohibiting cytochrome c release from mitochondria and neutralizing the function of Bax by forming heterodimeric complexes (Humphreys-Beher et al., 1999). In contrast, elevated Bax expression in SS salivary gland acinar cells may be associated with ongoing apoptosis through the formation of homodimeric complexes (Kong et al., 1997, 1998b).
A study of NOD and NOD-scid mice supports the idea that a potential apoptotic process in epithelial cells is dependent on Fas:FasL interactions (Kong et al., 1998a). FasL mRNA and protein were detected by reverse-transcription/polymerase chain-reaction (RT-PCR) and immunohistochemistry, respectively, in salivary and lacrimal gland epithelial cells from eight- and 18-week-old NOD, NOD-scid, and BALB/c mice, while Fas protein and mRNA were expressed only in the exocrine glands from 18-week-old NOD and NOD-scid mice. Glandular secretory epithelial cell apoptosis increased in NOD-scid in the absence of lymphocytic infiltration, suggesting the involvement of non-immune factors in the initiation of disease. Despite evidence for elevated apoptosis, controversy surrounds the actual levels reputed in both animal models and patients. This may be in part due to variability among laboratories in the use of the TUNEL method.
(D) Cytokines in NOD mice and human patients
Various studies indicate that human minor salivary gland biopsy tissue and salivary glands from mouse models of AEC exhibited Th1-type cytokine profiles at the sites of target organs regardless of biopsy classification. In addition, a significant increase in Th1 cytokine expression was associated with longer disease evolution (Kolkowski et al., 1999). Th2 cytokine mRNAs, such as IL-4 and IL-5, were detected occasionally in association with strong B-cell accumulation in the labial salivary glands (Ohyama et al., 1996). However, IL-4 is generally not detected at the site of the lesion in both human patients and animal models. Temporal analysis of cytokine mRNA expression in the submandibular glands of NOD mice demonstrated an increase between 10 and 12 wks of age in several pro-inflammatory cytokines, including IL-1β, tumor necrosis factor (TNF)-α, IL-6, IL-7, IL-10, IFN-γ, and inducible nitric oxide synthase (iNOS). Cytokine mRNA detected in lacrimal tissue was similar to that seen in the submandibular glands but appeared both earlier and more intensely (Robinson et al., 1998a,b).
IL-1β, IL-6, and TNF-α produced by epithelial cells were also detected, indicating that salivary gland epithelial cells may participate in the pathogenesis of SS (Fox et al., 1994). Up-regulation of IL-1β and down-regulation of IL-6 in a primary parotid gland acinar cell line from NOD mice at 10 weeks of age, preceding the infiltration, may suggest involvement of epithelial cells in the initiation of the disease, and alterations in the target environment independent of B- and T-lymphocytes (Yamakawa et al., 2000).
The contribution of an individual cytokine can be studied with the use of cytokine-knockout mice or transgenic mice exhibiting high or aberrant expression of the cytokine. Transgenic expression of IL-10 under the control of the amylase promoter induced apoptosis and lymphocytic infiltrates, which were composed mainly of FasL-expressing CD4+ cells, in the glands of normal mice (Saito et al., 1999). Despite this observation, IL-10-deficient NOD mice still develop normal disease progression (Brayer et al., submitted for publication). This may be due to the redundancy of functions that various cytokines share. Thus, blocking the function of one cytokine may not be enough to prevent the disease phenotype. In the development of diabetes in NOD mice, IL-10 appears to promote precocious disease progression (Wogensen et al., 1994), but has a protective role in an established diabetic condition by down-regulating pro-inflammatory Th1 cytokines (Zheng et al., 1997).
Further evidence for a role of humoral immune regulation of the loss of secretory function was revealed by the use of IL-4 knockout mice and IFN-γ knockout mice. Although IL-4 message is not detected in the NOD exocrine glands, genetic ablation of IL-4 affected the levels of IgG subclasses but not the numbers of B-cells and T-cells infiltrating the tissue. In addition, there was no loss of fluid secretion in these mice (Brayer et al., submitted for publication). In contrast, the IL-4 knockout had no effect on the timing or frequency of the transition to overt diabetes in NOD mice (Wang et al., 1998). This argues for the need to examine global roles of the cytokine in the mice and not just in the affected tissue, since immune cell proliferation and differentiation occur in draining lymph nodes upon activation.
Similarly, NOD mice deficient in IFN-γ did not exhibit the loss of secretory function. Interestingly, the salivary glands in NOD.IFN-γ-/- mice remained free of lymphocytes past 35 weeks of age, while lacrimal glands showed normal disease progression and immune cell composition. The architecture of salivary glands in these mice more closely resembled that of the normal control mice, and less apoptotic activity was detected in the gland, suggesting the possible involvement of IFN-γ in triggering the epithelial cell pathology of salivary glands at a pre-clinical, asymptomatic phase in NOD mice (Brayer et al., submitted for publication). Additionally, human salivary gland cells can be induced to express HLA-DR mRNA along with other inflammatory cytokine gene expression such as IL-1β, TNF-α, and IL-6, following stimulation with IFN-γ (Hamano et al., 1996). This could explain the participation of glandular acinar cells in the chronic re-stimulation of autoreactive T-cells in the glands when the acinar cells are in contact with the right signals for the up-regulation of class II molecules and the production of cytokines. In contrast, genetic absence of IFN-γ does not prevent either insulitis or diabetes in NOD mice. Instead of an expected increase in the expression of the Th2 T-helper cell-related cytokines, NOD IFN-γ-/- mice were shown with a profound decrease in the expression of IL-4 and IL-10 (Hultgren et al., 1996). Blocking the function of a cytokine by injecting antibody, soluble receptor, or receptor antagonist after maturation of the immune system has an effect different from that caused by inhibiting the function by gene knockout, in the sense that compensatory immune regulatory mechanisms may be favored in the gene knockout mice. Also, examining the roles of cytokines can be complicated, because of the dose and frequency of administration of cytokines and the degree of the disease as well as the age of the mice.
Data from the NOD.IL-4-/- and NOD.IFN-γ-/- mice reveal that both IL-4 (Th2-type cytokine) and IFN-γ (Th1-type cytokine), and the Th1/Th2 balance, may be essential for the disease progression and clinical manifestation of the autoimmune exocrinopathy in NOD mice. How they influence disease progression still needs to be further determined. A recent paper (Drakesmith et al., 2000) proposed that imbalances in cytokine environments can influence dendritic cell differentiation and antigen presentation, or they can affect glandular organogenesis to be favorable for immune cell infiltration, indicating important roles for cytokines in autoimmune diseases.
Studies of the cytokine gene knockout in NOD genetic background mice clearly indicate that the pathogenesis of SS is different from that of Type I diabetes. Thus, it is not surprising that tolerance induction of diabetogenic autoantigen does not prevent AEC (Leiter, 1993). In addition, the diabetogenic MHC, I-Ag7, seems to be essential for the development of diabetes but not AEC.
(E) Autoantibodies and pathogenesis
Various autoantibodies reactive with epitopes expressed in target organs have been detected in the sera from patients with autoimmune diseases (Bottazzo et al., 1974; Lernmark et al., 1978; McCarty, 1986; Hardin and Mimori, 1985; Bacman et al., 1996; Haneji et al., 1997; Robinson et al., 1998d). These antibodies, directed against nuclear, cytoplasmic, and cell-surface proteins, have been used as diagnostic tools for disease onset and progression (Tan, 1989; Fox, 1992; Haneji et al., 1997). One of the first-described antibodies associated with connective tissue autoimmune diseases was antinuclear antibodies (ANA) (Tan, 1989). Antigens for these ANA include the ribonucleoprotein SS-A/Ro and SS-B/La, in addition to Sm and Scl70 (Tan, 1989; Bachmann et al., 1990). Other sets of autoantigens appear to be generated during the process of apoptosis through the exposure of potential cryptic antigens following cleavage by activated caspases. For example, a 150-kDa proteolytic product of caspase cleavage of α-fodrin leads to the generation of autoantibodies in Sjögren's syndrome of potential diagnostic value (Haneji et al., 1997), although the role in disease initiation or progression has yet to be defined. A growing list of cellular target proteins has been identified which includes other cytoskeletal proteins (e.g., focal adhesion kinase), as well as signal transduction kinases (Reyland et al., 1999; Simbulan-Rosenthal et al., 1999; van de Water et al., 1999).
Autoantibodies to the muscarinic3 (M3) receptor, the acetylcholine receptor responsible for fluid secretion from the salivary glands, have been detected in sera from NOD mice and SS patients (Robinson et al., 1998d; Bacman et al., 1996, respectively). Furthermore, infusion of purified serum IgG or F(ab′)2 fragments from parental NOD mice or human primary SS patients alters stimulated saliva production in NOD.Igμnull mice which lack B-lymphocytes (Robinson et al., 1998d). Recently, an anti-rat M3 receptor monoclonal antibody was generated and reacted with mouse M3 receptor, in vivo, resulting in a reduction in secretory response in NOD-scid mice, further indicating the important role of autoantibodies in SS-like autoimmune exocrinopathy (Nguyen et al., 2000). Another study indicated that SS patient sera containing anti-M3 antibody or purified IgG from patient sera acted as an antagonist for M3-muscarinic receptors on isolated bladder smooth muscle, resulting in the inhibition of bladder contraction (Waterman et al., 2000). Receptor desensitization by repeated interaction with anti-M3 antibody may potentially result in changes in the normal signaling pathway, possibly through the regulation of downstream signaling targets. One potential target identified so far is the translocation of aquaporin protein to the apical plasma membrane of acinar cells for fluid secretion (Nguyen et al., 2000). Interestingly, when aquaporin5 null mice were generated by targeted gene disruption, defective saliva secretion by salivary glands was observed in these mice, despite the fact that tear secretion by lacrimal glands was not affected (Moore et al., 2000; Ma et al., 1999, respectively). Possibly, this may be due to differences in the mechanics of fluid secretion, such as α-adrenergic receptor as the major neuroreceptor for fluid secretion in the lacrimal glands (Dartt, 1994), as opposed to muscarinic receptor in the salivary glands. Additionally, the presence of other isoforms of aquaporins in rate-limiting steps for tear or saliva production and the different underlying mechanisms of disease pathogenesis in the two target organs may account for the observed results.
(IV) Summary
Studies on the NOD mouse and its congenic partner strains have begun to provide us with invaluable insights into the pathogenic mechanism(s) underlying the development of SS (Table 3). The NOD strain revealed the existence of two phases in the pathogenesis of SS. First, there is the disruption of homeostasis in the target tissue, resulting in increased apoptosis as well as cysteine proteinase activity, aberrant protein expression, and loss of amylase activity. Second, the involvement of immune components leading to activation of T- and B- cells and the generation of autoantibodies (e.g., anti-M3 receptor autoantibody) provide the hallmark clinical symptoms of dry eyes and dry mouth. The fact that severity of the symptoms does not always correlate with the degree of glandular pathology (Barchan et al., 1998) supports the concept of involvement of soluble factors such as cytokines or dysregulation of the parasympathetic nervous system by autoantibodies (Fu et al., 1993; Goin et al., 1994; Bacman et al., 1996; Ishikawa et al., 1998; Robinson et al., 1998d).
The M3 receptor is present in many tissues as well as in the lacrimal and salivary glands. Multiple organs with parasympathetic neural innervation include intestinal, vascular, bladder, and exocrine pulmonary smooth muscle, vaginal and dermal sweat glands, and iris sphincter muscle. All these tissues are reported to be affected in patients with secondary SS (Fu et al., 1993; Rose and Bona, 1993), supporting a role for the M3 receptor antibodies. Importantly, full secretory response was re-established in NOD Igμnull mice receiving either the patients' or NOD IgG following a one-week washout period (Robinson et al., 1998d). This could explain the importance of autoantibody in the disease pathogenesis and the possible treatment strategy of regulating the humoral arm of the adaptive immune response.
From the perspective of cytokine regulation of the immune response, the NOD mouse model provides an important example of how both Th1 and Th2 cytokines are involved in the initiation and progression of autoimmunity. Regulation of the cytokine environment and preferential activation of Th1 or Th2 cells can be achieved by direct manipulation of the cytokine environment or by administering antigen by particular routes, such as oral adminstration or IV injection, before the full-blown disease state. Another manipulation of the cytokine environment would be the use of soluble cytokine receptors. For example, soluble IFN-γ receptors to block the function of IFN-γ on the cells, as one potential immunomodulatory strategy, can be utilized once the drawback of its short half-life is overcome. Recently, the application of IFN-α via the oral route is in Phase II trial (Ship et al., 1999). The application resulted in decreased immune cell infiltration and increased secretory response. IFN-α may directly increase the density of aquaporin5 in salivary gland tissue by up-regulating the gene and the protein expression (Smith et al., 1999). Antagonizing the effects of IFN-γ by IFN-α/β may indirectly alter the outcome of pathological changes in patients with SS, as we observed in NOD.IFN-γ-/- mice.
The identification of non-MHC candidate genes in AEC in mice can be approached by microarray techniques and/or differential displays. Having high synteny between humans and mice, these candidate genes can be used for screening human libraries to determine how those genes will affect organogenesis and subsequently lead to the uniqueness of target tissue in the pathogenesis of SS. This could open up the possibility of gene therapy to alter the intrinsic problems in the glands. Additionally, once the primary neuroreceptor autoantigen initiating the clinical features of disease is confirmed, more reliable diagnostic tools for the disease will be developed, and intervention strategies to cure the disease will be readily available for patients with SS.
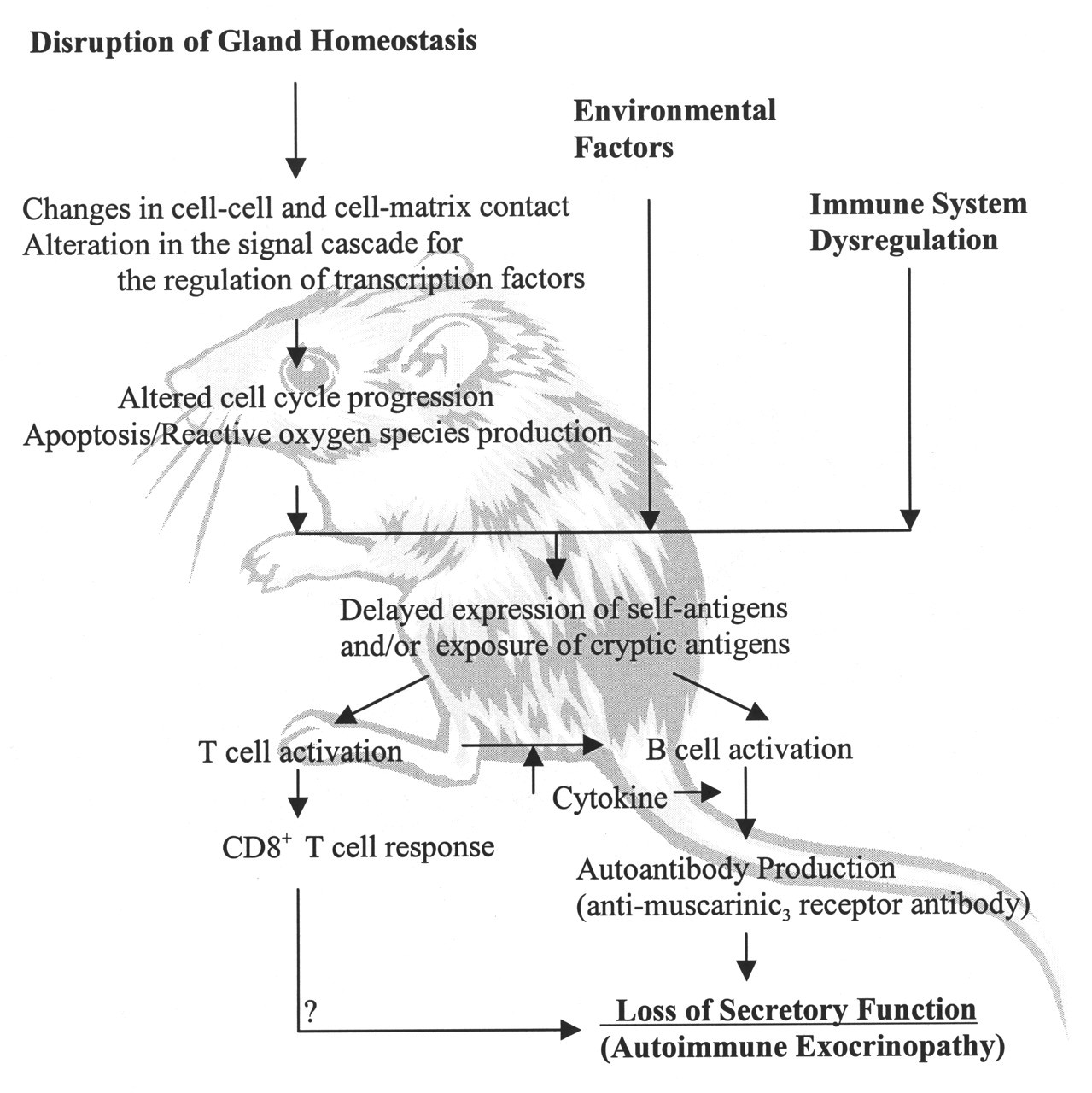
Proposed hypothesis of the initiation and the progression of Sjögren's syndrome-like autoimmune exocrinopathy in the NOD mouse.
Footnotes
†
Dr. Humphreys-Beher died while this article was in press.
Acknowledgements
This work was supported by NIDCR grants DE13290 and DE08778 to the late MHB. Dr. Seunghee Cha was further supported by the Department of Oral Biology at the University of Florida.