
Abstract
The recent success of autologous T cell-based therapies in hematological malignancies has spurred interest in applying similar immunotherapy strategies to the treatment of solid tumors. Identified nearly 4 decades ago, natural killer (NK) cells represent an arguably better cell type for immunotherapy development. Natural killer cells are cytotoxic lymphocytes that mediate the direct killing of transformed cells with reduced or absent major histocompatibility complex (MHC) and are the effector cells in antibody-dependent cell-mediated cytotoxicity. Unlike T cells, they do not require human leukocyte antigen (HLA) matching allowing for the adoptive transfer of allogeneic NK cells in the clinic. The development of NK cell-based therapies for solid tumors is complicated by the presence of an immunosuppressive tumor microenvironment that can potentially disarm NK cells rendering them inactive. The molecular imaging of NK cells in vivo will be crucial for the development of new therapies allowing for the immediate assessment of therapeutic response and off-target effects. A number of groups have investigated methods for detecting NK cells by optical, nuclear, and magnetic resonance imaging. In this review, we will provide an overview of the advances made in imaging NK cells in both preclinical and clinical studies.
Keywords
Introduction
Natural killer (NK) cells are potent immune sentinels constantly guarding our bodies against foreign invaders. Natural killer cells are cytotoxic lymphocytes of the innate immune system that do not require antigen priming and kill in the absence of major histocompatibility complex (MHC) presentation. 1,2 Their mechanism of action is fundamental in cancer immunosurveillance. 3 Natural killer cells are also effector cells for antibody-dependent cell-mediated cytotoxicity (ADCC)—the main mechanism of action for a number of therapeutic monoclonal antibodies. 4 In humans, NK cells are defined as CD3− CD56+ with further subdivision based on CD56 expression. 5 Natural killer cells recognize targets through families of activating (NKp46, NKG2C, CD84, CD16, etc) and inhibitory receptors (NKG2A, TIGIT, PD-1, etc). 2,6 The balance between these receptors determines their function. 2 Once an activating receptor is stimulated, a signaling cascade is initiated that results in the release of cytolytic granules and cytokines such as interferon gamma (IFN-γ) and Tumor necrosis factor (TNF)-α that recruit additional immune cells. 7,8
Natural killer cells are of immense clinical interest in part because they are easy isolated from the blood and can be expanded to clinically relevant numbers in vitro using antigen-presenting cells for patient infusion. 9 Natural killer cells also do not require donor matching and do not contribute to graft-versus-host disease. 1 Since allogeneic NK cells can be used for adoptive transfer, NK cell-based immunotherapies represent affordable treatment options that are truly off-the-shelf. The adoptive transfer of both autologous and allogeneic NK cells to improve immune function has become routine for leukemia and lymphoma patients. 10 This strategy has been successful when used in conjunction with lymphodepleting chemotherapy and cytokines such as IL-2, IL-15, or an IL-15 super agonist (ALT-803) for in vivo NK cell expansion. 10 -12 There has been a recent dramatic growth in the number of NK cell-based therapies undergoing clinical investigation. Several of the new therapies on the horizon involve the adoptive transfer of adaptive NK cells—NK cells that possess elements of memory—and activated NK cells coadministered with monoclonal antibodies to enhance ADCC. 13,14 Additionally, there are a number of ongoing clinical trials evaluating the therapeutic potential of NK cells in numerous solid tumors ranging from neuroblastoma to breast cancer.
A major challenge to implementing NK cell-based therapies in solid tumors is the immunosuppressive tumor microenvironment that could limit the antitumor activity of NK cells. 15 -17 The microenvironment of solid tumors is rich with elements of immunosuppression including MHC class I chain-related protein A and B (MICA and MICB), programmed death-ligand 1 (PD-L1) expression on tumor cells and macrophages, and high levels of myeloid-derived suppressor cells (MDSCs). 18 -20 For the development and successful administration of NK cell-based immunotherapies in solid tumors, the ability to accurately and quantitatively assess tumor homing and biodistribution by molecular imaging will be key. Imaging NK cell-based therapies will allow for an immediate assessment of potential therapeutic efficacy, off-target effects, severity of the immunosuppressive microenvironment, and quantification of NK cell expansion. This review highlights advances that have been made in the development of methodologies for the in vivo tracking of NK cells by molecular imaging.
Molecular Imaging Overview
Molecular imaging allows for the visualization of cellular and subcellular processes at both the molecular and anatomical levels. 21 It can provide not only the visualization of biological and pathophysiological processes, but also characterization and quantification of these processes. 22 The techniques used in molecular imaging typically rely on exploiting disease-specific molecular probes and intrinsic tissue characteristics. Currently, molecular imaging is used for diagnostic/prognostic purposes and to measure therapeutic efficacy in preclinical models and patient populations in the clinic. 23 Because NK cell-based immunotherapies are still in early stages of development, molecular imaging of NK cells is essential for the development process. Investigators have thus far developed a number of methods for the molecular imaging of NK cells using optical (bioluminescence, fluorescence, intravital microscopy [IVM]), nuclear, and magnetic resonance imaging (MRI; Table 1). Many of the techniques developed have focused on the direct labeling of the NK cell surface with fluorophores or the loading of NK cells with cell-permeable fluorophores, contrast agents, and radioisotopes. 23,24 These strategies have been of limited clinical utility because of the diffusion of imaging agents out of the cell and cell division diluting the signal beyond the detection limit.
A Summary of all of the Studies Mentioned.
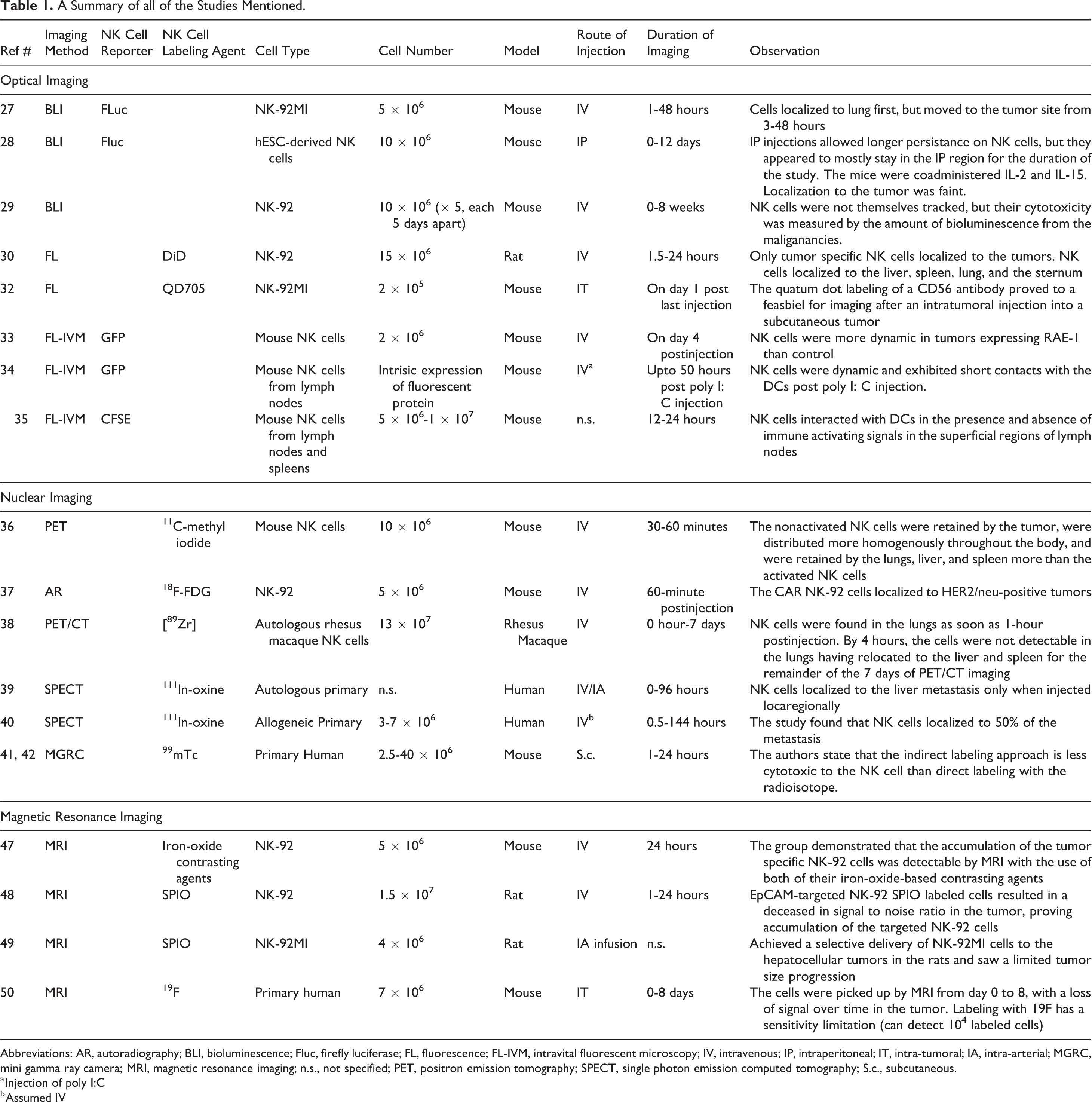
Abbreviations: AR, autoradiography; BLI, bioluminescence; Fluc, firefly luciferase; FL, fluorescence; FL-IVM, intravital fluorescent microscopy; IV, intravenous; IP, intraperitoneal; IT, intra-tumoral; IA, intra-arterial; MGRC, mini gamma ray camera; MRI, magnetic resonance imaging; n.s., not specified; PET, positron emission tomography; SPECT, single photon emission computed tomography; S.c., subcutaneous.
a Injection of poly I:C
b Assumed IV
Reporter gene imaging is potentially the ideal strategy for monitoring NK cell localization and tumor infiltration in vivo. This strategy will require the direct modification of the NK cell genome to express reporter genes that are uniformly passed down each cell division and are retained within the cell to maximize signal and sensitivity. Unlike primary T cells, primary NK cells are notoriously difficult to modify using traditional retro- and lentivirus methods. 2,25 This has led many investigators to use NK cell lines, such as NK-92 cells, that are easy to culture and genetically modify for research purposes. 25 It should be noted that many investigators refer to NK cell lines generically as “NK cells.” This is technically incorrect since NK-92 is a cancer-derived NK-like cell line. NK-92 cells lack the expression of NK cell activating receptors such as CD16, NKp44, and NKp46. 26 There have been proposals to use them in clinical trials to treat cancer, but since NK-92 cells are cancer-derived, possess cytogenetic abnormalities, and are latently infected with Epstein-Barr virus, they must be irradiated prior to patient infusion making them replication incompetent and of questionable clinical utility. A majority of the imaging studies to date have used the NK-92 cell line because of their ease of use compared to primary NK cells.
Bioluminescence Imaging
Bioluminescence imaging (BLI) is a commonly used tool for the quantification of tumor growth in either dissemination or intraperitoneal tumor models that have been engineered to express luciferase (firefly or renilla). The direct BLI of NK cells is a relatively unexplored field. Studies involving NK cells and BLI in the past have been mostly restricted to imaging luciferase expressing cancer cells and their response to NK cell therapy. The biodistribution of NK-92MI cells, a derivative of the NK-92 cell line, has been monitored by BLI. In this study, NK-92MI cells were transduced to express firefly luciferase and a thyroid cancer cell line CAL-62 was modified to express renilla luciferase. This allowed correlative BLI imaging to be performed on NK92MI cells localized to a pulmonary metastasis model derived from the CAL-62 cell line. Luciferase expressing NK-92MI cells (5 × 106) were injected into mice via tail vein and imaging was carried out for 48 hours postinjection. The NK-92MI cells were observed to migrate to the sites of the metastasis model and reached maximum intensity at 24 hours. 27
Knorr et al performed a similar study using human embryonic stem cell (hESC)-derived NK cells that were engineered to express firefly luciferase. Also for this study, leukemia cell lines were engineered to express membrane-bound Gaussia luciferase or a near-infrared fluorescent protein, TurboFP650. Repeated dual imaging experiments were performed and similar results using both the dual-bioluminescence and bioluminescence/fluorescence methods were obtained. Both methods showed localization of hESC NK cells to the tumor, but the group reported the dual bioluminescence method was difficult due to the timing of injections and the kinetics of the substrates. Localization of NK cells to the tumors was also confirmed with immunohistochemistry by staining for NKp46, a marker more specific than CD56. 28 However, in the localization in experiments, the luciferase signal from the NK cells did not appear strong in the tumor region. The group performed both intraperitoneal and intravenous injections of NK cells, but found that they lost the NK cell signal after the first time point by intravenous injection. The subsequent tumor localization studies were performed using intraperitoneal injections of the luciferase expressing NK cells. In another study, Swift et al assessed the effect of the NK-92 cell line on a human multiple myeloma cell line transduced to express green fluorescent protein (GFP) and luciferase. Mice with luciferase expressing multiple myeloma cells were imaged 4 weeks after multiple myeloma inoculation (3 weeks after last NK-92 injection). Mice treated with NK-92 exhibited lower disease burden compared to controls over a time course of 8 weeks. 29 This study did not involve the imaging of the NK cells, but rather only the tumor to quantify regression.
Fluorescence Imaging
Few literature reports exist on the fluorescence imaging of NK cells or NK cell lines. In 2009, Tavri et al used fluorescence to image an NK-92 cell line engineered with a chimeric antigen receptor (CAR) for the epithelial cell adhesion molecule (EpCAM). The targeted NK-92 cell line was labeled with a near-infrared dye 1,19-dioctadecyl3,3,39,39-tetramethylindodicarbocyanine (DiD)
Labeling of the cells with DiD had no effect on cell viability and subsequently 15 × 106 labeled cells were injected via tail vein into rats bearing subcutaneous DU145 prostate cancer tumors positive for EpCAM. 30 The study confirmed that the CAR NK-92 cells accumulated in the tumor, while the parental nontargeted NK-92 cells did not. The signal remained constant from hour 8 until the end of the study at 24 hours. The NK-92 cells in both the targeted and control groups were found to localize to the liver, spleen, lung, and the sternum after 24 hours. 31
A study by Lim et al involved the labeling of NK-92 MI cells with an anti-CD56 antibody coated with QD705, a quantum dot that emits in the near-infrared region. Using quantum dots for imaging has several advantages such as high quantum yield, color availability, good photostability, and small size. Quantum dots are particularly useful for NK cell imaging since they are not readily internalized by the cells. This study primarily focused on a proof-of-concept that a quantum dot labeling approach can be used for NK cell line imaging. The NK-92MI cells labeled with anti-CD56 antibody coated with QD705 were injected directly into a subcutaneous MeWo tumor (derived from human lymph node metastasis). The NK-92MI injections were performed on 2 separate days and imaged the day after the second intratumoral injection. The NK cells in the tumor were detected and tumor regression was observed in mice administered the NK cells. This study documented that the QD705 labeling had minimal toxicity on the NK cells as demonstrated by cell viability results done by fluorescence-activated cell sorting analysis. 32 The NK cells were also tested for IFN-γ production and cytolytic activity to assess for normal cell function. The labeled NK cells showed no significant difference from the control in these activities, therefore the quantum dot labeling also did not compromise the antitumor activity of the NK cells.
Intravital Microscopy Imaging
Multiphoton or “two-photon” IVM has had a dramatic impact on understanding cellular processes in living systems. Two-photon IVM uses a near-infrared excitation laser to excite common fluorophores leading to increased tissue penetration and decreased photobleaching and toxicity. Intravital microscopy allows for the facile monitoring of living tissue and cells, such as the highly dynamic immune system. Denguine et al performed two-photon IVM on GFP-expressing mouse NK cells to determine the effect the NKG2D receptor had on intratumoral NK cell dynamics. In addition to NK cells, the NKG2D receptor is expressed on NK-T cells and some T cells. The NKG2D receptor recognizes ligands related to MHC class I proteins, particularly retinoic acid early-inducible-1 (RAE-1) family of proteins. Such ligands are often overexpressed by mouse and human tumors. For this study, RAE-1β expressing tumors in mice were used for IVM. The GFP-NK cells (2 × 106) were injected via tail vein into mice with tumors overexpressing the RAE-1 ligand. Four days postinjection, IVM was performed. It was discovered that NK cells were activated and highly dynamic in the tumors overexpressing RAE-1β compared to the nonexpressing tumors. 33
Natural killer cell dynamics have also been studied by Beuneu et al using two-photon IVM. Natural killer cell activation by dendritic cells (DCs) in mouse lymph nodes was monitored using genetically modified mice that expressed GFP-NK cells and YFP-DCs. Natural killer cells can be activated by innate cytokines, in which DCs are able to secrete when challenged by a viral stimulant. The IVM studies indicated that in response to a synthetic analog of a viral double stranded RNA, NK cells were dynamic and exhibited intermittent contact with the DCs. Although it was previously known that DC interaction with NK cells is necessary for NK cell activation in response to viral stimulation, the details of this interaction were not established. The authors examined the NK cell–DC interactions in response to viral stimulation in vivo and found that the contacts last between 1 and 3 minutes. The length of NK cell–DC interaction is contrastingly short when compared to T-cell interaction with DCs in response to the same stimulant. The group found that the T-cells do not dissociate from the DCs throughout the duration of their experiment. 34 Bejénoff et al also studied NK cell behavior in lymph nodes via their interaction with DCs after an infection with Leishmania major. Their IVM data indicated that NK cells interacted with DCs in the presence and absence of immune activating signals in the superficial regions of lymph nodes. The infection, however, did recruit some NK cells to the paracortex and induce them to secrete IFN-γ. In this study, NK cells extracted from the spleens and lymph nodes of naïve mice were labeled with a fluorescein derivative and adoptively transferred (5 × 106-107) for IVM. 35
Nuclear Imaging
The 2 nuclear imaging modalities positron emission tomography (PET) and single-photon emission computed tomography (SPECT) have been employed to track radiolabeled NK cells with high sensitivity. In 1993, Melder et al labeled IL-2 activated and nonactivated murine NK cells with the PET tracer [11C]methyl iodide. After labeling, 10 × 106 NK cells were injected into tumor bearing mice via the tail vein and trafficking was monitored up to 1 hour postinjection. The findings of this study were surprising in that the nonactivated NK cells infiltrated the tumor, were distributed more homogenously throughout the body, and were retained by the lungs, liver, and spleen more than the activated NK cells. Though further longitudinal was not possible due to the short half-life of the 11C tracer, this pioneering study demonstrated that PET can be used to quantify NK cell tumor infiltration and biodistribution. 36 Since then, the NK-92 cell line has also been loaded with the commonly used PET tracer [18F]fluorodeoxyglucose ([18F]FDG). Meier et al confirmed the localization of [18F]FDG loaded NK-92 cells to human epidermal growth factor receptor 2 (HER2) positive tumors in mice by digital autoradiography. In this study, the NK-92 cells were modified with a CAR for HER2 prior to loading. 37 More recently, Davidson-Moncada et al used 89Zr-labeled autologous rhesus macaque NK cells that were expanded ex vivo for PET imaging. The 89Zr-labeled NK cells (13 × 107) were injected into a rhesus macaque and tracked by PET for 7 days. Natural killer cells were observed to localize in the lungs as early as 1 hour post-injection. By 4 hours, however, the cells were not detectable in the lungs having relocated to the liver and spleen for the remainder of the 7-day imaging experiment. This study also observed limited numbers of NK cells in the lymph nodes and bone marrow. 38
Single-photon emission computed tomography has been used in the clinic to image NK cell tumor infiltration in patients with cancer. In one study, autologous NK cells were labeled with 111In-oxine, activated ex vivo, and subsequently injected back into 3 patients with colon cancer that has metastasized to the liver. Patients were injected twice overall during the imaging study—once intra-arterially directly into the liver and once intravenously with 30 days between the injections. The results from the study showed that NK cells were confined to the liver and spleen when injected intra-arterially. However, when the same patients received an intravenous injection, the NK cells appeared first in the lungs prior to their trafficking to other tissues. It was determined that the 111In-oxine labeled NK cells localized to the metastatic lesions in the liver only after the intra-arterial administration. 39 This was not the first time this labeling approach was used. A few years prior to this study in metastatic colon cancer, primary allogeneic NK cells (3-7 × 108) labeled with 111In-oxine were injected intravenously into renal cell carcinoma patients with metastatic disease. This study found that the NK cells localized to 50% of the metastasis as confirmed by [18F]FDG-PET imaging. 40
More recently, Galli et al used indirect labeling of NK cells by targeting the CD56 receptor with a 99mTc-labeled antibody. Human NK cells injected intravenously into nude tumor-bearing mice were tracked by this method. The indirectly labeled NK cells localized to the tumor, liver, and kidneys as shown by imaging with a planar γ camera. Imaging was only performed out to 24 hours postinjection of the radiolabeled antibody due to the half-life of 99mTc. The authors state that the indirect labeling approach was less cytotoxic to the transferred NK cell population than direct labeling with auger electron emitting 111In-oxine. 41,42 In one of the previously mentioned studies, however, the viability of the NK cells labeled with 111In-oxine was 96% postlabeling. 40 Thus, it is difficult to the assess NK cell viability and expansion postinjection after increased exposure to a radioisotope such as 111In or 131I. It has been established that radionuclide labeling can be toxic to NK cells and that the toxicity is both radionuclide and dose dependent. 43 -45
Magnetic Resonance Imaging
Magnetic resonance imaging is one of the most commonly used imaging modalities in the clinic. It is noninvasive and can easily be translated from preclinical to clinical applications. The main drawback of MRI is the lack of sensitivity toward injected contrast agents. 46 One of the earliest MRI studies imaging NK cells was performed by Daldrup-Link et al where the biodistribution of HER2-targeted CAR NK-92 cells was determined using iron-oxide-based contrast agents. The CAR NK-92 cells were labeled with ferumoxides and ferucarbotran and intravenously injected (5 × 106) into mice bearing HER2 positive xenografts. Localization of the CAR NK-92 cells was detectable by MRI using both iron-oxide-based contrast agents. This study also determined that the CAR NK-92 cells could only be labeled efficiently using electroporation and lipofection, but not incubation. They found that simple incubation resulted in minor labeling with ferucarbotran and no labeling at all with ferumoxides, however, no insight was given as to why simple incubation was inefficient. Electroporation significantly lowered cell viability for a labeling efficiency of 50% to 60%, while lipofection was relatively nontoxic for the same degree of labeling. The salient purpose of this study was to provide a proof-of-concept that Food and Drug Administration approved contrast agents could be used to track NK cell distribution using a clinical 1.5 T MR scanner. 47
Half a decade later, CAR NK-92 cells targeting EpCAM were imaged using superparamagnetic iron-oxide (SPIO) ferumoxides in a DU145 prostate cancer xenograft model. The injected cells (1.5 × 107) were imaged 1- and 24-hour postinjection. Compared to nontargeted NK-92 SPIO labeled cells, the CAR NK-92 SPIO labeled cells resulted in a significant decrease in signal-to-noise ratio in the tumor, proving infiltration of the targeted NK-92 cells. The tumor infiltration by the CAR NK-92 cells was additionally confirmed by histopathology. 48 Sheu et al employed a similar approach using SPIO nanoparticles to label NK-92MI cells. Incubation of the NK-92MI cells with the SPIO nanoparticles resulted in decreased cell viability, but a labeling efficacy of 88%. The labeled cells were infused by intra-arterial delivery into the liver of rats implanted with hepatocellular carcinoma cells. This direct infusion method achieved selective delivery of NK-92MI cells to the tumors and limited tumor growth was observed compared to the controls. 49
Labeling with SPIO has proven to be a popular method for imaging NK cells with MRI. In 2012, Mallett et al labeled KHYG-1 NK cells, an NK cell line derived from a patient with aggressive NK cell leukemia carrying a p53 point mutation, 50 with SPIO to image a subcutaneous PC3-M prostate cancer model in nude mice. 51 The SPIO labeled KHYG-1 NK cells were injected using different routes of administration: tail vein, IP, and subcutaneous (20 × 106, 20 × 106, and 2 × 106 cells, respectively). Signal loss via MRI was only observed in the subcutaneous injected mice, however, histology detected NK cells in the tumors from all 3 routes of NK cell administration. More recently, there have been studies on imaging primary human NK cells by MRI rather than NK cell lines. In 2016, Bouchlaka et al labeled primary human NK cells with 19F perfluoropolyether. 19F is a nonradioactive, 100% naturally abundant spin ½ isotope of fluorine that can be formulated for incubation-based cell labeling 52 ; and 19F-MRI has the advantage of long-term tracking, but suffers from poor signal-to-noise. 53 Primary human NK cells were expanded ex vivo with 19F and injected the cells intratumorally into 2 different mouse xenograft models. The cells were observed by MRI from day 0 to 8, with a loss of signal over time in the tumor, suggesting NK cell migration to different organs. The authors state that labeling with 19F had a sensitivity limitation (can detect 104 labeled cells), which may prevent detection of cells delivered intravenously or percutaneously. 54
Conclusion
Molecular imaging is an important tool during both the preclinical and clinical stages of therapeutic development. Agents that specifically localize to the target tissue and demonstrate little secondary accumulation in nontarget tissues can be selected by employing molecular imaging to maximize therapeutic efficacy and minimize harmful side effects. With the number of NK cell products for clinical investigation increasing annually, molecular imaging is certain to play important role in their clinical development. Thus far, no strategy has emerged as the gold standard for imaging NK cell tumor infiltration and localization in vivo. Few of the imaging studies documented in this review even used primary NK cells, rather the vast majority used NK cell lines. Also, none of the strategies in this review can be used accurately for the longitudinal imaging of adoptively transferred NK cells weeks postinjection. The ability to detect NK cells by nuclear imaging is dictated by the half-life of the radioisotope and MRI may not be sensitive enough for most clinical applications. Optical imaging is an excellent laboratory tool, but it is not translatable. An ideal strategy for imaging NK cells in vivo would use a reporter gene such as herpes simplex thymidine kinase or human sodium iodide symporter for PET or SPECT imaging. 55 With the NK cell continuously expressing the reporter gene, radioactive substrate tracers could be administered hours, days, and weeks postadoptive transfer to monitor NK cell biodistribution and expansion in vivo. Since the reporter gene is stably integrated into the NK cell genome and passed down each cell division, multiple successive generations of adoptively transferred NK cells could be imaged using this strategy in theory.
To date, there are no published reports on the in vivo imaging of primary NK cells that have been genetically modified to express reporter genes for any imaging modality. The intransigence of primary NK cells to genetic modification has largely hindered exploration of reporter genes for in vivo imaging postadoptive transfer. Recently, we optimized methods to introduce large DNA constructs into the genomes of primary human NK cells. This allowed us to successfully engineer primary NK cells from a healthy donor to express a near-infrared fluorescent protein reporter gene. This protein was originally developed by Scherbakova and named iRFP682 based on its emission wavelength. 56,57 The expression of iRFP682 in our system was driven by a strong, constitutive MND promoter. 58 Flow cytometery and a LiCor Odyssey Infrared Imager were used to confirm expression of iRFP682 in modified primary NK cells that had expanded ex vivo (Figure 1A and B). Different numbers of primary NK cells expressing iRFP682 were mixed with matrigel (2.5 × 105-3.0 × 106) and injected subcutaneously above the left leg of the mice. Imaging on an IVIS Spectrum Optical Imaging System saw a cell number-dependent increase in fluorescence (Figure 1C).
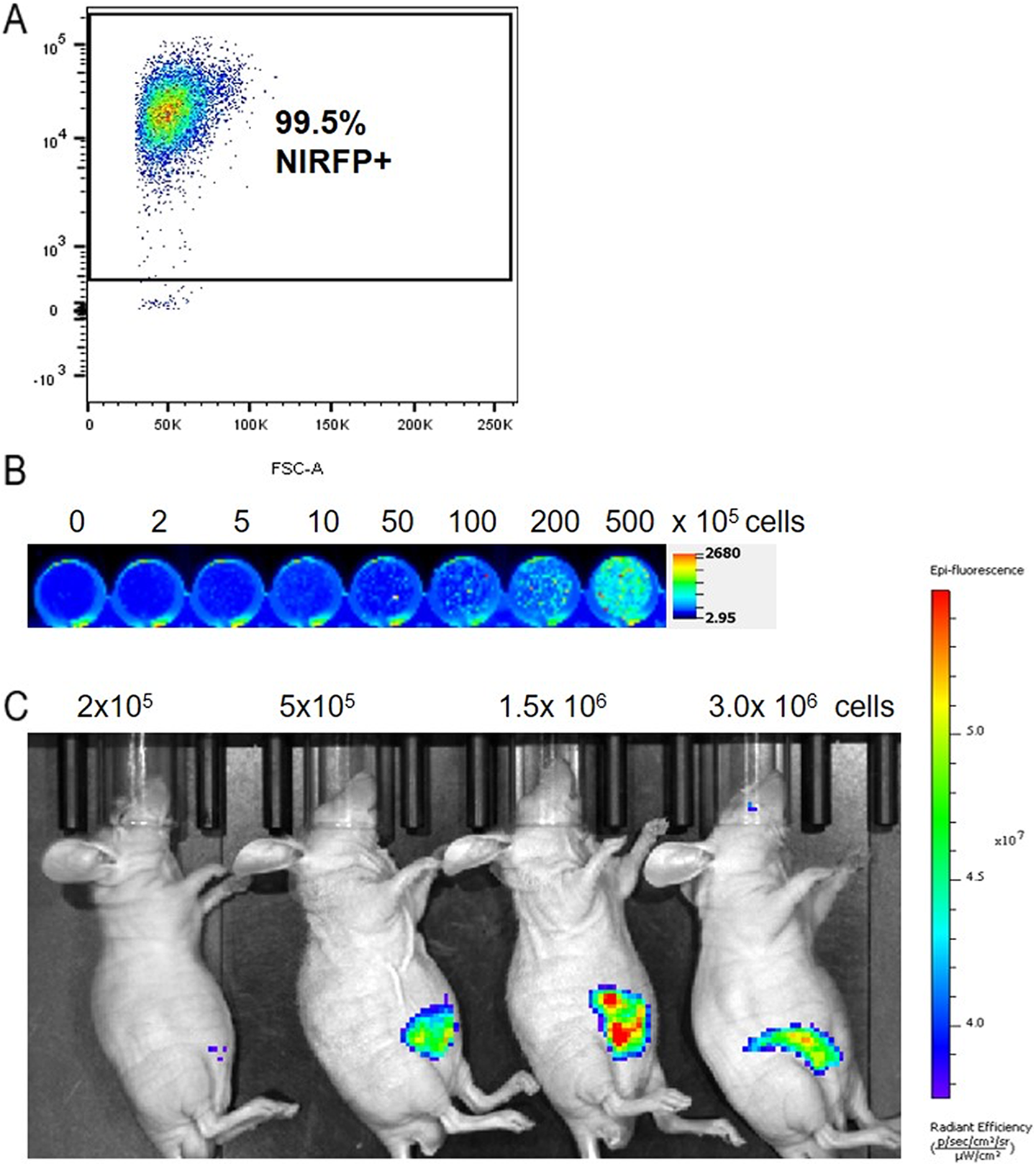
Establishment of iRFP682-expressing primary natural killer (NK) cells. A, Confirmation of iRFP682 expression in primary NK Cells by flow cytometry postmodification and sorting. Cells were also sorted for CD56 prior to transduction. B, NK cell fluorescence documented on the Licor Odyssey Infrared Imager using the 700 nm channel. C. NK cells were subcutaneously injected into mice and imaged 3 hours postinjection (ex/em = 675/720 nm)
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.