
Abstract
Bacillus subtilis is a successful host for producing recombinant proteins. Its GRAS (generally recognized as safe) status and its remarkable innate ability to absorb and incorporate exogenous DNA into its genome make this organism an ideal platform for the heterologous expression of bioactive substances. The factors that corroborate its value can be attributed to the scientific knowledge obtained from decades of study regarding its biology that has fostered the development of several genetic engineering strategies, such as the use of different plasmids, engineering of constitutive or double promoters, chemical inducers, systems of self-inducing expression with or without a secretion system that uses a signal peptide, and so on. Tools that enrich the technological arsenal of this expression platform improve the efficiency and reduce the costs of production of proteins of biotechnological importance. Therefore, this review aims to highlight the major advances involving recombinant expression systems developed in B. subtilis, thus sustaining the generation of knowledge and its application in future research. It was verified that this bacterium is a model in constant demand and studies of the expression of recombinant proteins on a large scale are increasing in number. As such, it represents a powerful bacterial host for academic research and industrial purposes.
Impact statement
The use of Bacillus subtilis to produce recombinant proteins with medical, biotechnological, and industrial value is a growing activity that presents an enormous potential for exploration. This host has been standing out and gaining notability because it offers laboratory safety and excellent yields. Such elements stimulate more and more the development of expression systems that use particular and unconventional genetic engineering strategies. We revised the wide availability and screening of several efficient promoters, in addition to the use of economically viable chemical inducers and self-inducible expression systems, which are increasingly in demand today for their practicality. We hope this article clarifies recent systems that comprise the technological arsenal available for this expression platform and that may be useful to various research groups as reliable alternatives to the worldwide need for bioactive produced by a scalable source of production.
Introduction
Bacillus subtilis is a Gram-positive bacteria host qualified by the Food and Drug Administration (FDA) as being a generally safe microorganism (GRAS) that is free of exotoxins and endotoxins. 1 The reading of its codons is remarkably diverse and contributes to the expression of heterologous genes that do not depend on additional steps. 2 The applications of B. subtilis include food fermentation, cell division studies, development of biofilms, production of secondary metabolites, and it is also a potential host for vaccine production, and a delivery vehicle.3–7
In addition, it has established itself as a robust cell host and an efficient technological platform for the expression of bioactive and high-yield protein production, with the potential for scaling in a bioreactor. The main challenge in the production of recombinant proteins in B. subtilis is that of how to choose a relevant expression system. 8 Different expression systems have already been developed, and proteins from prokaryotic or eukaryotic organisms have been synthesized and purified. 9 The best strategy depends on the protein to be expressed, though always with the aim of cost reduction, efficiency, and high-yield production. 10 New strategies include the use of constitutive promoters, double promoters, functional synthetics that are capable of directing transcription, efficient signal peptides, and inducible and self-inducing expression systems.11,12
These improved characteristics, together with the growing demand for biotechnology products, contribute to the increase in the use of this bacterium, thus endorsing its widespread use in the field of genetic and metabolic engineering.13,14 Since the first report of heterologous overproduction of recombinant proteins in this strain, the mass production of an infinity of valuable bioactive molecules has been performed on a relatively large scale. 15
Recognizing the importance of metabolic engineering, systematic biology, synthetic biology, and evolution-based engineering in B. subtilis, this review first summarizes the importance of technical knowledge and the construction of improved strains suitable for industrial production. Next, we present an overview of the B. subtilis secretion system and its crucial role in heterologous protein production. Finally, we review the versatility of the application of promoters and others expression mediators as strategies that aim at production efficiency, also recent advances inductor-free expression vectors construction.
Therefore, this review covers the latest and the principal expression systems developed in B. subtilis and the strategies inherent to the multivalence of this expression platform and confirm its wide capacity and its recognition as an important host in the modern biotechnological industry. The bibliographic survey of indexed publications was performed using the PUBMED, Google Scholar, and “Periódico Capes” databases. Only publications in the English language and from 2015 to 2020 were included. The keywords used for the search were “Bacillus subtillis”, “Expression Systems”, “Promoters”, “Induction”, “Self-induction”, and “Recombinant proteins”.
The B. subtilis expression platform
B. subtilis is constantly referred to as a reference system and a collection of biochemical, genetic, and physiological knowledge of Gram-positive bacteria. 16 Its sequencing was carried out by Kunst et al. 17 in 1997 using strain 168 and involved more than 30 laboratories worldwide. Subsequently, other research groups carried out a complete sequencing of the genome that included criteria that ensured the characterization of its genes and products. 18
Bacterial preservation centers around the world have conserved numerous strains of B. subtilis, including the auxotrophic strains 23, 122, 160, 166, and 168, which were derived originally from B. subtilis Marburg. 19 The 168 strain has been extensively used in academic research and industrial production. 20 Given the availability of an increasing number of cloning vectors and mutant host lines that are provided by the Bacillus Genetic Stock Center (BGSC), 21 many strains can be transformed using systematic metabolic engineering to obtain mutant strains that meet production requirements. 22 For example, B. subtilis WB600 and B. subtilis WB800 were constructed from B. subtilis 168 by knocking out six and eight protease genes, respectively.20,23 Although some proteases may be of interest to the industry, their activity generally limits the overall efficiency of heterologous protein production. 23 However, the use of these strains becomes attractive, as it partially solves the problem of proteolysis of secreted target proteins. 24
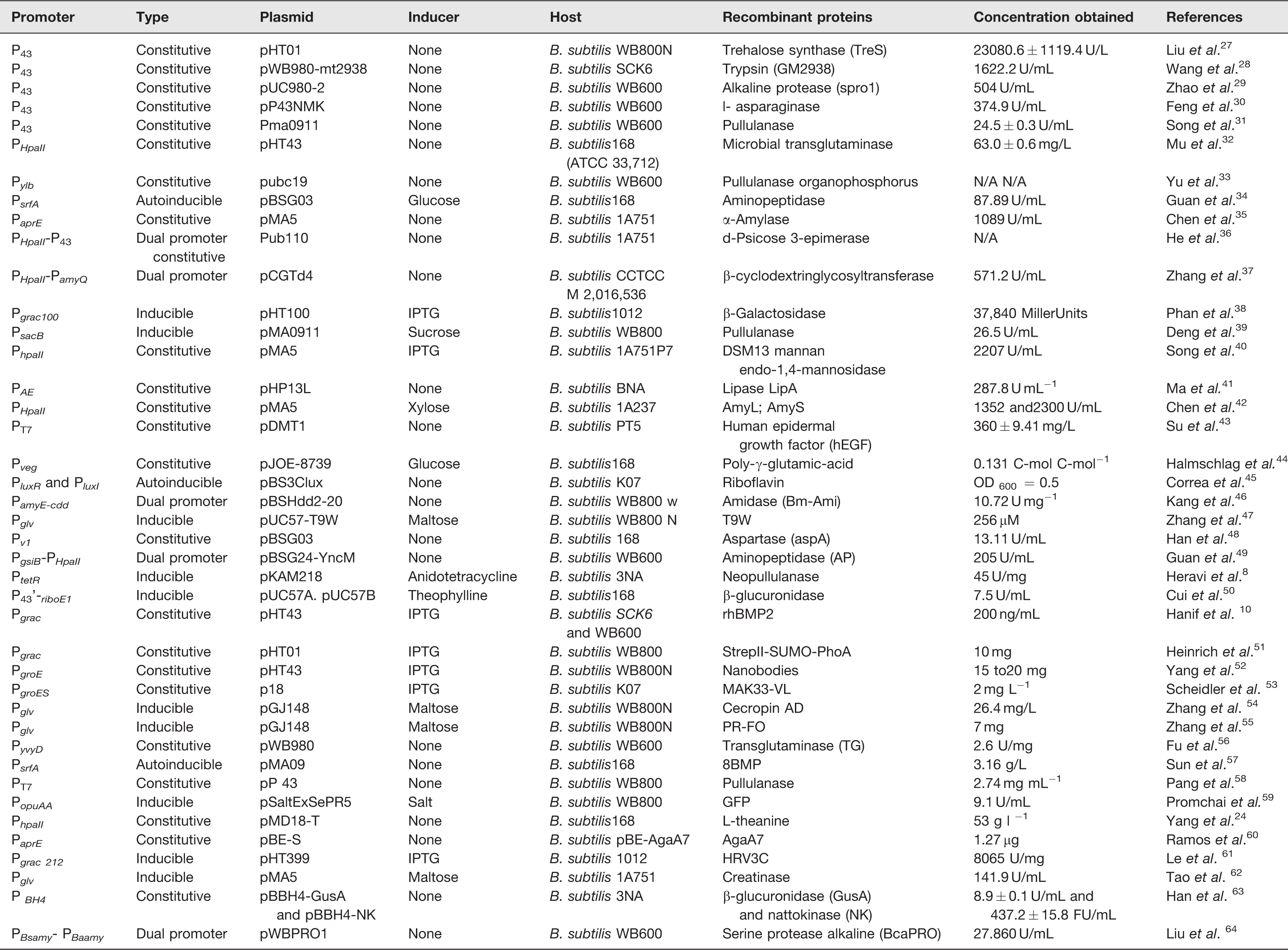
These sequencings and the emergence of powerful genetic tools culminated in the rise in popularity of B. subtilis, mainly in the industrial segment. 25 It is one of the most frequently used bacterial hosts for producing high-value recombinant proteins. 26 Furthermore, there has been an unprecedented increase in the number of molecules produced in this model, which has provided it with a prominent role in the industrial production of proteins worldwide and has transformed B. subtilis into the most studied species of the genus Bacillus. 23 Several examples are shown in Table 1.
Expression systems developed in Bacillus subtilis from 2015 to 2020.
The production of some protein classes can be difficult, for example when it comes to proteins with multiple subunits or with a high molecular weight that faces problems relating to transport across the cell membrane. This requires the use of alternative transport routes which the determining mechanisms related to these types of non-classical secretory pathways are still limiting and unclear.65,66 The sequences of synthetic genes also need to be optimized to achieve the best possible expression, 67 since, in the same way, that occurs in other species, some codons are rarely used, and the selection of synonymous codon is restricted by the efficiency of translation. Highly expressed genes are almost always more dependent on the tRNA content and tend to have a strong predisposition of codons usage.68,69 From a structural point of view, unlike other bacterial hosts, B. subtilis does not have an external membrane or periplasmic space, which gives it a greater capacity for protein secretion directly into its surrounding environment. 20 And based on proteomic analysis studies, B. subtilis has already demonstrated the potential to export approximately 300 proteins. 70
The secretion system in B. subtilis
The steps for producing recombinant proteins in B. subtilis are relatively simple (Figure 1(a)). To ensure that the proteins are properly directed to the pathways and thus initiate the translocation process, specific signal peptides are present at the N-terminus of each protein and must be cleaved by a peptidase for later release of the mature protein to be exported to the medium, retained in the cell wall, or retained at the cell membrane interface.70,71 B. subtilis contains a strict quality control system for protein-dependent on its machinery and performed by intracellular and extracytoplasmic chaperones, cell wall proteases, and extracellular proteases.20,72 And several different protein export systems have been identified in this bacteria so far. 73
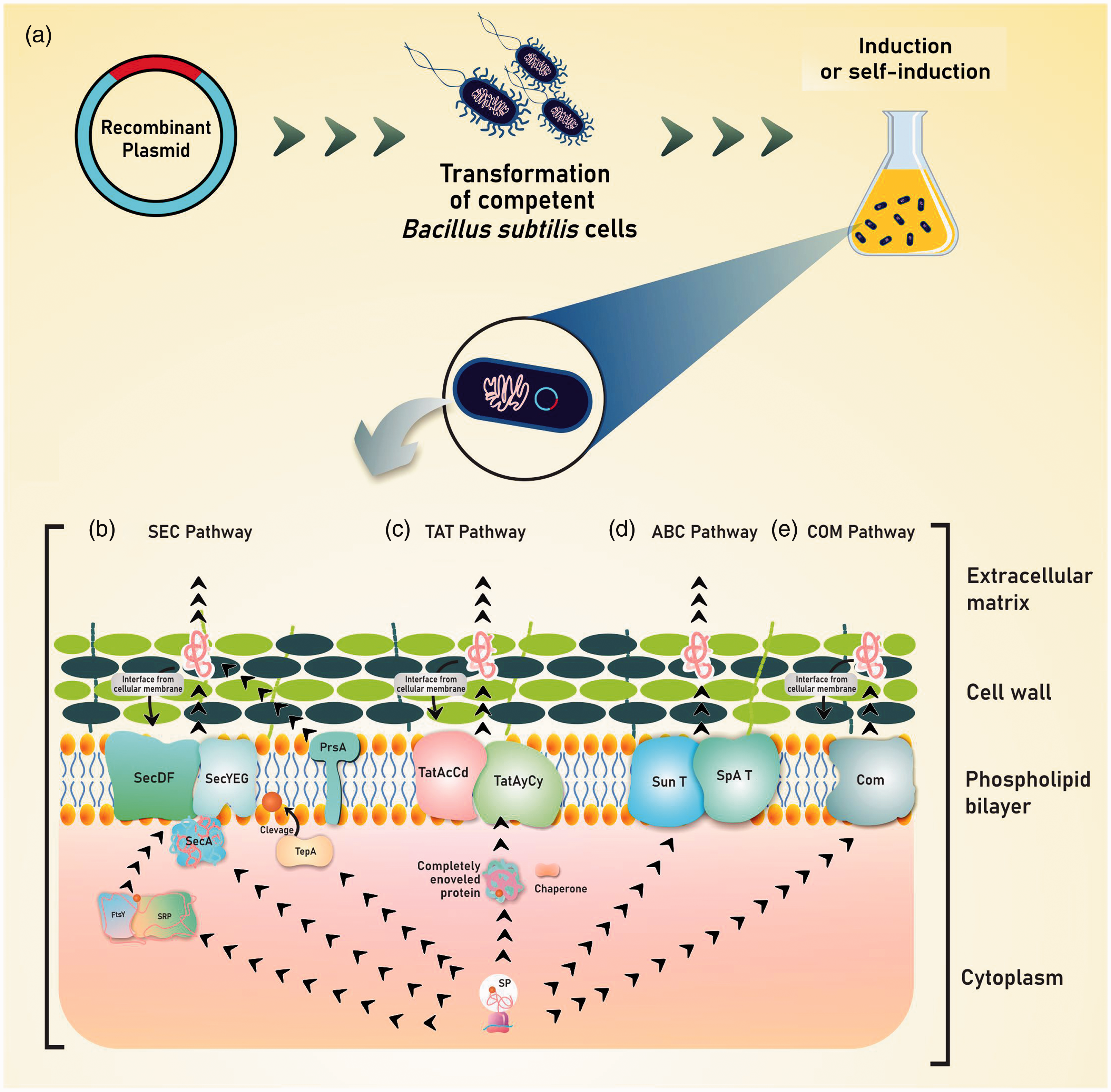
Steps for heterologous expression in B. subtilis and the four protein export pathways through the cytoplasmic membrane. (a) Binding of the gene of interest to an expression vector, a transformation of competent cells, and subsequent induction or self-induction of expression. (b) In the Sec path, the precursor protein containing a signal peptide is recognized by the signal recognition particle (SRP) and interacts with the FtsY protein, directing the protein to the membrane translocases (SecA, SecDF, and SecYEG). The TepA clidase peptidase, the signal peptide, and the PrsA lipoprotein aid in protein folding. Another form of transport is through a soluble SecA that guides the protein to the translocation channels, with subsequent removal of the signal peptide. (c) In the Tat pathway, the protein is entirely folding into the cytoplasm with the aid of chaperones, being leaded to two different translocation systems (TatAcCd and TatAyCy). (d) In via ABC, proteins can use the Sun T and SpaA T export systems to facilitate their secretion. These two classes of translocators have a dual function: proteolytic activity in addition to transport activity. (e) In the pseudopilin (Com) export pathway, proteins are exported in a specific way and can be retained at the cell membrane wall interface. (A color version of this figure is available in the online journal.)
The general secretion pathway (Sec)-dependent transport system and the “Twin-arginine” (Tat) translocation system are generally studied and used for the secretion of recombinant proteins in the extracellular medium in B. subtilis.20,25 The general secretion pathway (Sec) (Figure 1(b)) includes components that convert energy in the form of ATP and transmembrane protons as the driving force to direct proteins through channels incorporated in the membrane. 74 The general secretion system (SeC) is considered the main route for the secretion of proteins in the growth medium and has the characteristic of transporting proteins across the cytoplasmic membrane in an unfolded or weakly folded conformation.75,76 In the co-translational export mode, the protein is synthesized ribosomally as a precursor protein containing a signal peptide in the N-terminal region, being helped to maintain its competent state of translocation by cytoplasmic chaperones, with the subsequent recognition of the signal peptide by the particle signal recognition (SRP), as well as targeting membrane translocases. 77 The targeting of the pre-protein by this SRP complex also involves the presence of the FtsY protein, a highly conserved GTPase that also acts in targeting the SecYEG transmembrane channels. 78 In this way, the PrsA lipoprotein is necessary for the subsequent folding of the mature protein, in the assistance for providing stability, and avoiding non-productive interactions with the cell wall. 79 Another strategy in the post-translation export mode, SecA transfers the precursor proteins to the membrane translocation channel, then the SecA motor protein translocates the pre-proteins through SecYEG using metabolic energy from ATP hydrolysis. 80
In Tat (Figure 1(c)), before the translocation of the recombinant protein through the cytoplasmic membrane, they become completely bent in the cytoplasm with the help of intracellular chaperones, in a strategy capable of reducing proteolytic degradation. 21 On the other hand, in the Tat-dependent pathway, secreted proteins usually contain a typical highly conserved cleavable signal peptide that directly affects secretion. 81 The precursor protein strongly folded by intracellular chaperones is transported through a channel formed by the translocase complex. 82 In B. subtilis, two separate translocation systems work in parallel: TatAdCd and TatAyCy. TatAdCd is expressed as a translocase under phosphate-deprived conditions, while TatAyCy is expressed constitutively.20,71 TatAd and TatCd combine to form the TatAdCd complex, while TatAy and TatCy form the TatAyCy complex. As shown in Figure 1(d), proteins also can be exported to the medium via ATP binding cassette transporters (ABC). 77 In contrast (Figure 1(e)), small numbers of proteins are exported by the pseudopilin (Com) export route. 73
The constitutive promoters in B. subtilis
Notably, many efforts have been made to identify strong promoters for transcription control. In this sense, P43 is a promoter that meets widely used requirements. P43 was first described in the studies of Wang and Doi in 1984 and best characterized by Song and Neuhard in 1989.83,84 It was used to produce methyl parathion hydrolase and to develop an expression system for B. subtilis based on food-grade expression plasmids.85,86
In a recent study, the P43 promoter was successfully fused with other promoters to construct a series of plasmids to extend the screening of signal peptides (SPs) and to obtain promoters suitable for secretion of BcaPRO (alkaline serine protease) from B. clausii. The signal peptide “DacB” and the dual promoter “P Bsamy -P Baamy ” demonstrated the best performance, with a high expression level of BcaPRO (27,860 U/mL). 64 Liu et al. 27 regulated the extracellular activity of the TreS enzyme (trehalose synthase) using the P43 promoter to produce trehalose, a stable non-reducing disaccharide. In this study, a TreS mutant was successfully expressed and secreted in B. subtilis WB800N. 27 Currently, P43 is regularly used in comparative studies to measure the strength of different promoters in B. subtilis, showing that, although new stronger promoters can be identified, it is an important parameter for studies involving screening of promoters for the improvement of fermentation conditions and expression levels.28,29
Several other promoters have been characterized to increase heterologous expression in B. subtilis. These characterizations arise from the accumulation of genetic, biochemical, and structural information on the molecular mechanism underlying promoter stimulation. 87 It is important to emphasize that strong promoters are necessary since their characteristics lead to more efficient transcription of the genes encoding the protein of interest. Seo and Schmidt-Dannert in 2019 used the strong promoter P veg in combination with regulatory elements of Pseudomonas putida to investigate and control the expression of green fluorescent protein (GFP) in B. subtilis. 88 This new system has proved promising and useful for studies of metabolic engineering, synthetic biology, and production of proteins of industrial interest. 88
As a result of numerous studies, the repertoire of techniques has expanded, and improvement of the efficiency of promoters via chromosomal integration has become an option. For example, Zhou et al. 89 improved the activity of the promoter P ylb , and optimized the central region -35, -10, and the upstream sequence (UP) by replacing both sequences with consensus sequences. 89 The final promoter exhibited almost 26 times the activity of β-galactosidase (BgaB) and 195 times the intensity of the super-folded green fluorescent protein (sfGFP).
The dual promoters in B. subtilis
Another promising approach for increasing productivity that has received widespread attention due to its high efficiency and continuity is the use of dual promoters. 90 In the study of Guan et al., 49 satisfactory results were achieved with the dual promoter P gsiB -P HpaII , which showed better performance and increased the production of aminopeptidase (AP) after 45 h of fermentation. Another dual promoter system that presented remarkable applicability was P HpaII -P amyQ , which produced β-cyclodextrin glycosyltransferase (β-CGTase), and mediated the expression of substantial extracellular pullulanase and the expression of α-cyclodextrin glycosyltransferase (α-CGTase). 37 These enzymes were also produced using the PsodA-fusA- amyE triple promoter.91,92 Kang et al. 46 used a system containing a P amyE-cdd dual promoter and a (Pac) signal peptide to increase the expression of a recombinant amidase (Bm-Ami). The extracellular activity of Bm-Ami containing the plasmid pBSHdd2-20 reached 10.72 U/mg−1 DCW after 52 h in a scaling fermentation that, according to the authors, was the highest secretion obtained from Bm-Ami to date. 46
The functional synthetic promoters in B. subtilis
One technique that has proven to be useful is the development of functional synthetic promoters. Synthetic promoters are more resistant to imperfect growth conditions such as biotechnological processes that cause cellular stress or metabolic load and generate a strong transcription of target genes. 93 This makes them an important alternative since natural promoters are often not universally characterized due to their poor performance. 94 Han et al. 63 developed a new strategy called gradual evolution for screening robust B. subtilis bacterial promoters that was aimed at the promoter center spacing sequence (SETarSCoP). A series of mutant promoters was obtained that overexpressed β-glucuronidase (GusA) of E. coli and nattokinase (NK) of Bacillus natto. The mutant promoter P BH4 was identified and verified as an exceptionally strong synthetic promoter. In this same study, this technique was successfully used to increase the strength of the constituent promoter P ylbP and the inductive promoter by xylose P xylA and proved itself to be a new method for the manufacture and screening of promoters. 63
The inducer-dependent promoters in B. subtilis
The first inducible expression system reported in B. subtilis was described after pioneering studies by Yansura and Henner in 1984, in which the spac promoter was induced by isopropyl-ß-D-thiogalactoside (IPTG). 95 This system is a combination of a B. subtilis phage promoter SPO-1 and the lac operator from E. coli that deactivates the constitutively produced lac repressor (LacI). 8
Yang et al. 52 transformed the modified strain WB800N with the pHT43 vector, which contained a strong promoter derived from the B. subtilis operon groE converted into an IPTG-inducible promoter. 52 This study reported for the first time the secretion of fragments of heavy chain antibodies or nanobodies of camelids (Nbs). The yields of Nbs were estimated at 15 to 20 mg from one liter of bacterial culture, which is comparable to the yields in systems for producing Nbs in E. coli (15 to 20 mg). 96
Scheidler et al. 53 pioneered the development of an efficient amber suppression system in B. subtilis by using IPTG as an inducer. This system enables the expression, secretion, and direct purification of a target protein carrying non-canonical amino acids (ncAAs). The authors expressed the light chain variable domain of a murine monoclonal antibody fragment (MAK33-VL) and obtained 2 mg L−1 of culture. The reported expression system creates the possibility for producing therapeutically and biotechnologically significant proteins containing bio-orthogonal groups subject to chemical decoration. 53
Le et al. 61 used the robust promoter P grac212 , inducible by IPTG, to overexpress human rhinovirus protease 3 C (HRV3C) in the cytoplasm of B. subtilis cells. In the study, the proteases His-HRV3C and His-GST-HRV3C were overexpressed in the cytoplasm of B. subtilis in 11% and 16% of total cellular proteins, respectively. The largest specific protease activities were 8065 U/mg for His-HRV3C., 61 which indicates that it is an effective method for producing recombinant proteins in the cytoplasm of B. subtilis.
Although the IPTG inductor has been amply used, models with other chemical inducers, such as the carbohydrates sucrose, mannose, xylose, maltose, and starch, have been developed and are considered efficient and cheap. The IPTG inductor is not consumed by bacteria and this is an advantage over inducible sugar promoters such as P malA , P manP , and P mtlA .97–99 However, using the carbohydrate glucose as an inducer proved to be efficient in strains built to express biopolymers of industrial interest such as poly-γ-glutamic acid (γ-PGA). Using the constitutive Pveg promoter, the yield of γ-PGA increased by 129% when using glucose as the sole carbon source. 44
In the case of the maltose-inducible P glv promoter, which was used to secrete T9W, a variant of pig myeloid antimicrobial 36 (PMAP 36) with high antimicrobial activity, this system proved to be an economical and environmentally correct approach for biotechnological production. 47 Furthermore, maltose was likewise the inducer in a safe method to produce Cecropin AD (CAD), a hybrid peptide with biological activity. 54
Other important induction systems are the ones in which induction is based on the signal peptide, such as the system LIKE (from the German “LIa-Kontrollierte Expression”) that is based on the lial promoter, and the system SURE (subtilin-regulated gene expression). Both systems are strictly controlled by the addition of bacitracin and lantibiotic subtilin after the exponential phase of cell growth, thus preventing leakage of transcription in non-inductive conditions. 100
Studies involving the signal peptide have become increasingly viable since this mechanism can guide secretion of the heterologous protein to a specific area with more precision and thus effectively increase the solubility of the protein. 55 Previous studies have shown that the signal peptides SP sacB, SP amy, SP aprs, and SP aprl simultaneously guided the expression of pullulanase, with the guidance of SP sacB being the most efficient for secretory expression. 101 Su et al. 43 described six SPs to guide the secretion of the human epidermal growth factor (hEGF). The highest level of production was obtained when the hEGF gene was fused with SP xynD (360 ± 9.41 mg/L), which revealed an excellent system for the expression of proteins of cosmetic and pharmaceutical interest. 43
Heinrich et al. 51 identified a signal peptide coding region with a high capacity for protein expression and secretion. Based on the plasmid pHT01, an expression vector was constructed for encoding a fusion protein. The cassette constructed in this study demonstrated great efficiency in encoding the YoaW-StrepII-SUMO signal peptide in B. subtilis and gave rise to a system capable of maximizing yield as well as improving translocation, folding, and stability of heterologous proteins.
The inducible expression systems controlled by riboswitches in B. subtilis
As known inducers, there are a variety of synthetic riboswitches that have been developed to control heterologous expression and which represent relevant engineering targets for the biotechnological production of compounds.102,103 In this perspective, a system with great compatibility and robustness was obtained in the studies of Cui et al., 50 in which a new genetic element composed of the promoter P43 and a theophylline-riboswitch was developed and characterized in B. subtilis from the union of the constitutive expression characteristics of P43 for a dose-dependent induced pattern of theophylline. 50 The authors noted the system overexpressed β-glucuronidase with an induced expression level higher than three other strong constitutive promoters, including P srfA , P aprE , and the native P43.
The conditionally activating promoters in B. subtilis
One of the simplest induction systems already described are temperature change controlled. Li et al. 104 characterized two temperature-sensitive promoters, P2 and P7, isolated from B. subtilis. The production of β-galactosidase conducted by these promoters was higher at 45°C than at 37°C. In a later study, aiming to improve the overproduction of “difficult to express” proteins, Welsch et al. 105 developed a low-temperature expression system in B. subtilis that was based on the cold-inducible promoter of the des gene. The improved expression system was validated by the overproduction of xylanase, as well as an α-glucosidase from Saccharomyces cerevisiae, thus confirming the adequacy of this host organism for the overproduction of critical substances.
Recently, self-inducible expression systems have been attracting more attention due to the practicality of their services.93,106 Self-inducible promoters, such as P pst and P cry3Aa can express the target gene from the late log phase to the stationary phase without the need for an inducer, which facilitates the efficient production of low-cost proteins and peptides.57,107 An inductor-independent self-induction system was developed by Wenzel et al. 108 using the mutant strain TQ356 to further improve cost efficiency and product yield. In this system, glucose prevented induction through repression by carbon catabolism, creating ideal conditions for self-induction. This technique led to an almost three-fold increase in production and reached a 14.6% yield of recombinant protein (eGFP). 108
Guan et al. 34 built a self-inducing and self-regulating expression system by simply adding glucose, using the quorum detection-related promoter (P srfA ). In this case, the central sequence of -10 and -35 was replaced by consensus sequences. In the end, the system proved its efficiency by the successful production of aminopeptidase. In later studies, Guan et al. 109 demonstrated that it is possible to integrate the corresponding genes downstream of the promoter in the plasmid or chromosome in the BSG1682 strain of B. subtilis. The results revealed that the P23 promoter, derived from the P srfA promoter, presented the best performance, and was almost twice as strong as P srfA . Two heterologous proteins, aminopeptidase (AP) and nattokinase (NK) were overproduced in this study. 109
In addition, Correa et al. 45 developed a method of dynamic regulation by detecting quorum in B. subtilis, which was capable of self-monitoring and inducing expression without human supervision. The promoter response was 2.5 and 3.2 times stronger than the well-characterized promoters P srfA and P veg , respectively. The researchers even applied the strongest self-induction device to produce vitamin B2 and self-induction proved to be modular and adjustable. 45 Furthermore, Correa et al. 110 presented a set of data to analyze different growth parameters of OD600, fluorescence measurement of GFP, luminescence, and riboflavin production. The purpose of this validation was to support the efficiency of this self-inducing device for the expression and control of genes in B. subtilis. 110
The inducer-free expression vectors base on pgrac
The study of expression vectors without inducer is still a challenge due to a series of industrial production bottlenecks, but there are already some promising investigative studies. 111 In this regard, Tran et al. 112 generated inducer-free expression vectors using IPTG-inducible promoters in the absence of the LacI repressor and containing the strong P grac100 promoter. The levels of expression of β-galactosidase (bgaB gene) were at least 37 times higher when compared to the inducible constructions in the absence of IPTG. These plasmids may also be convenient for studying gene expression at different scales for recombinant protein production. 112
Tran et al. 113 demonstrated the great potential of using strong promoters from the P grac family in the overproduction of recombinant proteins through the construction of integrative inductor-free vectors, capable of integrating into the B. subtilis genome in both the lacA locus and the amyE locus. In this study, the promoters P grac 100- bgaB and P grac 212- bgaB yielded 20.9% and 42% of intracellular proteins, respectively. Whereby, after the incorporation of P grac 212- bgaB in both the amyE and lacA loci, the expression of β-galactosidase (BgaB) was observed in up to 53.4% of intracellular proteins 113
In another strategy that aims to support fundamental research studies in B. subtilis, Chu et al. 114 constructed inducer-free expression plasmids containing the hybrid promoter P spac , inducible by IPTG, suitable to express in a controlled way and at lower levels proteins with specific functions in the host cell and reporter genes. As expected, the BgaB and GFP + target genes were expressed at low levels compared to inducer-free plasmids with the strong promoters P grac01 and P grac100 , about 16.2 to 20.3 times for BgaB and 24.7 to 34.3 times for GFP +, respectively. The authors concluded that the use of these inducer-free vectors containing the P spac promoter enabled the constitutive expression of heterologous recombinant proteins at low levels desired in B. subtilis. 114
Conclusions
In summary, the literature consulted expands the knowledge regarding the functional expression systems that are commercially available and developed from different lineages of B. subtilis. We note the existence of a large arsenal of precursors and recent studies that accredit this bacterium as one of the main platforms for the expression of heterologous proteins, thus promoting progress in bioproduction. Furthermore, its wide use is justified due to its genetic and metabolic characteristics that allow proper planning and provide experimental security. The success of this model in expressing biocomposites is due to the development of massive strategies aimed at creating and constantly guaranteeing the efficiency of its promoters, plasmids, the use of low-cost varied inducers, and transcriptional self-regulatory elements, among others. As a result, the high technological status achieved increasingly encourages the improvement of existing genetic engineering techniques, as well as the search for the development of new efficient approaches, which makes the Gram-positive B. subtilis bacteria a robust and promising host with biotechnological excellence for the optimal expression of different recombinant proteins with valuable applications. Although the advances in omics studies are clear and decisive for the affirmation of this robust biotechnological host, and many strategies have culminated in the expression of various recombinant proteins, future studies need to focus on overcoming many limitations regarding the understanding and exploration of secretion pathways. New strategies must need to be developed to overcome current bottlenecks, e.g. expression of medical and industrial importance multiple subunits proteins.
Footnotes
AUTHORS’ CONTRIBUTIONS
All authors participated in the design and review of the manuscript. CCS, LAMM, and SDSP performed the literature review, CCS wrote the manuscript, and JMG provided the images.
ACKNOWLEDGMENTS
We thank the Universidade Federal do Amazonas (UFAM), Instituto Leônidas e Maria Deane (ILMD), Fundação Oswaldo Cruz (FIOCRUZ AMAZÔNIA) and the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) for their support.
DECLARATION OF CONFLICTING INTERESTS
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
FUNDING
The author(s) received no financial support for the research, authorship, and/or publication of this article.