
Abstract
Microglia begin colonizing the developing brain as early as embryonic day 9, prior to the emergence of neurons and other glia. Their ontogeny is also distinct from other central nervous system cells, as they derive from yolk sac hematopoietic progenitors and not neural progenitors. In this review, we feature these unique characteristics of microglia and assess the spatiotemporal similarities between microglia colonization of the central nervous system and embryonic neurogenesis. We also infer to existing evidence for microglia function from embryonic through to postnatal neurodevelopment to postulate roles for microglia in neurogenesis.
Introduction
Microglia are the tissue-specific macrophages of the central nervous system (CNS). Typically known for mediating inflammation in the CNS, continually emerging roles for microglia in physiological conditions emphasize their unique functional niche as both mononuclear phagocyte and glia (reviewed in Ransohoff and Cardona 1 ). Microglia actively survey the CNS to regulate homeostasis, provide trophic support for neurons and regulate synaptic development; functions that underscore their diverse roles. Microglia are also distinct in their origin. While neurons, astrocytes and oligodendrocytes develop from neuroepithelial cells, microglia are derived from yolk sac macrophages and appear in the CNS prior to the aforementioned cells.
Embryonic neurogenesis involves symmetric and asymmetric cell division for the purpose of expanding the pool of neural precursors and differentiating them. The progenitors that give rise to mature neurons of the CNS are not a single homogenous population, as their continuing division and differentiation generate a repertoire of neural progenitors at different developmental time points (reviewed in Gotz and Huttner 2 and Kriegstein and Alvarez-Buylla 3 ). As microglia share the same spatiotemporal compartment as neural progenitors, their influence on the development, survival and differentiation of neural progenitors is of great interest. Reviews on the effects of microglia in neurogenesis are available, with a focus on early postnatal and adult neurogenesis within the hippocampal dentate gyrus and lateral ventricles 4 or on the pathological or aging brain. 5 In this review, we describe microglial roles in embryonic neurogenesis, primarily focusing on their regulation of the neural precursor pool size.
Embryonic neurogenesis: A cell biology perspective
Our description on embryonic neurogenesis focuses on the mouse neocortex (unless specified otherwise) as the majority of the literature lies here. There are several key events that form the neocortex: (1) proliferation of neuroepithelial cells creates a multilayered neural plate, the innermost apical layer being the ventricular zone (VZ), (2) continued proliferation and differentiation of neuroepithelial cells and formation of a second germinal zone, the subventricular zone (SVZ), (3) formation of neurons and their positioning away from the neurogenic regions towards the pial surface, resulting in the six layers of the cortex and (4) emergence of astrocytes and oligodendrocytes during late embryogenesis, just prior to birth (reviewed in Kriegstein and Alvarez-Buylla 3 ). The size of the neural precursor pool is, therefore, not static, shifting as progenitors and their intermediates form, proliferate and differentiate. A slowdown in cell proliferation (largely due to the lengthening of the Gap 1 (G1) phase),6–8 developmental cell death,9,10 phagocytosis of precursors 11 and differentiation all may contribute to the retraction of the neural precursor pool, and this machinery requires tight regulation for sculpting of CNS circuitry.
The cellular development of the embryonic brain begins with neuroepithelial cells—the most primitive neural precursor that gives rise to all neurons and macroglia in the CNS. Neuroepithelial cells exhibit features that resemble epithelial cells, including expression of tight junctions and adherens junction proteins.12–14 They also express an intermediate filament protein, nestin 15 and the transmembrane protein prominin-1 (CD133). 16 First forming a single layer of cells called the neural plate; neuroepithelial cells symmetrically divide to expand the thickness of the neural plate. They then begin expressing features of astrocytes and assume a radial morphology, giving rise to a population of cells termed radial glia.
Radial glia are conserved across vertebrates and present in the embryo throughout neurogenesis. They are a heterogenous population of cells which interestingly share properties similar to astrocytes, for instance, they contain glycogen granules and express the brain lipid-binding protein, 17 astrocyte-specific glutamate transporter, 18 adhesion molecule tenascin-C, 19 calcium-binding protein S100β 20 and intermediate filament vimentin. 21 Radial glia signify a common progenitor for neurons, astrocytes and oligodendrocytes and appear to be a more fate-restricted progenitor compared to neuroepithelial cells. 18 It is most likely that CNS progenitors consist of populations of single-fate restricted progenitors, i.e. they are either of neuron-producing lineages or glia-producing lineages, and multipotent progenitors (a single progenitor capable of differentiation into multiple CNS cell types) as evidence exists for both. For further reading, Kriegstein and Alvarez-Buylla 3 and Malatesta et al. 22 offer detailed reviews on radial glia and neural stem cells. In mice, neuroepithelial cells transform to radial glia at around E9 to E10 and completes at E12.23,24 From E14 to E18, radial glial cells divide asymmetrically to generate neurons or produce intermediate progenitor cells that transiently increase the progenitor cells pool before terminally differentiating into specific neuronal cell types. 18 Throughout cortical neurogenesis, the somata of radial glia lie in the VZ, but their radial processes traverse to the basement membrane at the pial surface. Newborn neurons use these processes as scaffolds to migrate towards the cortical plate.25,26 Another important temporal event of CNS development is the switch from the neurogenesis to gliogenesis machinery (in mice at around E16 and until birth).18,27 At this point, radial glia cease producing neurons and began to generate astrocytes and later, oligodendrocytes.18,27 This diminishes the radial glia population in most regions of the brain, except for populations in the cerebellum and retina (known as Bergmann glial cells and Muller cells, respectively).
The distinct ontogeny of microglia
Given the fact that microglia share many phenotypic features with other myeloid cells in the body (including circulating monocytes and tissue macrophages), it is a general consensus that microglia are of hematopoietic origin. Nevertheless, the precise identity of microglia progenitors remained a great debate until recently. Microglia were initially thought to arise from circulating monocytes and bone marrow precursors that are recruited to CNS tissues. That notion was supported by in vitro studies demonstrating that monocytes from peripheral blood and the bone marrow can be induced to differentiate into microglia-like cells with comparable phenotypes and functional abilities in terms of secretory profiles, oxidative burst and phagocytic activities.28–30 Also, several studies exploited chimeric, transgenic and pharmacological techniques to show that circulating monocytes enter the CNS and differentiate into microglia.31–37 A noteworthy caveat associated with these approaches that ablate the local, in situ microglia population, alter phenotype of the remaining microglia, disrupt CNS homeostasis and/or compromise integrity of the blood brain barrier,32,36,38,39 is that it may trigger compensatory mechanisms to allow infiltration of monocytes into the CNS to reconstitute into microglia-like cells. In fact, it has been demonstrated that recipient mice with brains shielded from irradiation do not experience significant engraftment of bone marrow cells into the brain. 31 The parabiosis technique has greatly aided with this caveat. In non-irradiated parabiotic mice, the contribution of blood monocytes to the microglia population is negligible unless the test subjects were preconditioned with total body irradiation.38,40 Moreover, in a recent study using a CX3CR1CreER system that depletes microglia without perturbing the blood brain barrier, new microglia that repopulated the brain-derived exclusively from an internal CNS-resident pool. These newborn microglia were highly proliferative and their gene and protein profile placed them closer to microglia compared to bone marrow-derived brain macrophages. 36 Collectively, these studies suggest that under a normal physiological state, there is insignificant contribution (if any) of circulating monocytes to the microglia population. In the context of disease, however, it has been shown that the experimental autoimmune encephalomyelitis (EAE) model of multiple sclerosis causes a transient entry of monocytes, however, they do not become permanent members of the microglia population. 41 Seminal work by Butovsky et al. 42 demonstrates that even after infiltrating the inflamed CNS, monocytes retain a genetic and protein signature that is remarkably distinct from microglia; therefore, the lineage-based delineation between microglia and monocytes remains unperturbed even within the same tissue microenvironment.
Two waves of embryonic hematopoiesis give rise to macrophages–primitive hematopoiesis and definitive hematopoiesis. During primitive hematopoiesis, primitive macrophages develop from yolk sac progenitors from E8.5 to E9 and colonize the entire embryo beginning E9–E10. Definitive hematopoiesis generates hematopoietic stem cells (HSCs) that colonize the fetal liver around E10–E10.5 (reviewed in Ginhoux and Jung 43 ). From here, HSCs colonize tissues and replace the yolk sac macrophages. Microglia, however, are unique as the definitive wave of hematopoiesis does not contribute to their population. Primitive yolk sac progenitors are in the brain rudiment earlier than the emergence of monocytes.44,45 Instead, they emerge from c-kit+ yolk sac progenitors and begin migrating to the brain at E9.40,44,46 By E10.5, microglia are present in the cephalic mesenchyme and neuroepithelium and undergo intense proliferation.40,44,47 Although Langerhans cells of the skin do retain a small population of yolk sac macrophages, microglia are the only macrophage population believed to be derived predominantly from primitive haematopoiesis.40,47,48 In essence, the yolk sac progenitors that enter the developing brain at E9 are responsible for the population of microglia in the adult.40,47 Microglia also are maintained in the CNS without contribution from circulating monocytes as they are capable of in situ renewal. 38
The sole yolk sac-origin of microglia is conserved across vertebrate species.40,49–51 Importantly, unlike macrophages that form during definitive hematopoiesis, yolk sac macrophages develop independent of the transcriptional activator MYB 47 and instead are transcription factor PU.1- and interferon regulatory factor 8 (IRF8)-dependent. 46 MYB is required for stem cell development in the bone marrow, while PU.1 and interferon regulatory factor 8 (IRF8) both modulate the development of microglia. PU.1 is also exclusively expressed by all hematopoietic cells. This makes microglia the only adult macrophage population that are exclusively MYB-independent. 43 Differences between microglia and other tissue macrophages in terms of ontogeny and localization may be why microglia have low major histocompatibility complex class II (MHC II) expression and less antigen presentation capabilities compared to other macrophages.52–55 These appear to be a phenotype suited towards the relatively immunoprivileged status of the CNS.
These series of circumstances reveal the unique identity of microglia. Collectively, the brain comprises a distinct population of macrophages compared to other tissues in terms of ontogeny, lineage and phenotype. This begets us to question the reasons for the singularity of microglia and how it may affect embryonic neurogenesis. The early occurrence of microglia in the developing CNS is bound to have functional consequences, which we explore later in the next section.
Effects of microglia on neural progenitors in the developing brain
Intriguingly, the colonization of microglia in the developing CNS closely correlates with several developmental milestones of neurogenesis. The timeline of microglia invasion into the neural plate at E10 coincides with the emergence of radial glia in the embryonic brain.24,40,44 We do not believe that this spatiotemporal similarity is serendipitous, although there is yet to be a known role for microglia that this early in development. From E12 onwards, increasing numbers of microglia are progressively found throughout the developing cortex, with a dense population in the VZ and SVZ.56–59 This places microglia in regions of the brain that are key sites for neurogenesis and is an observation also made in other vertebrates including primates, rodents and humans. 11 The microglia in these areas express CD11b, MHC II and inducible nitric oxide synthase (iNOS) which are receptors and mediators involved in antigen presentation and inflammation. 11 Following cortical neurogenesis, microglia are evenly distributed throughout the cortex, potentially to prepare for homeostatic functions.
Overview of studies on microglia and embryonic neurogenesis
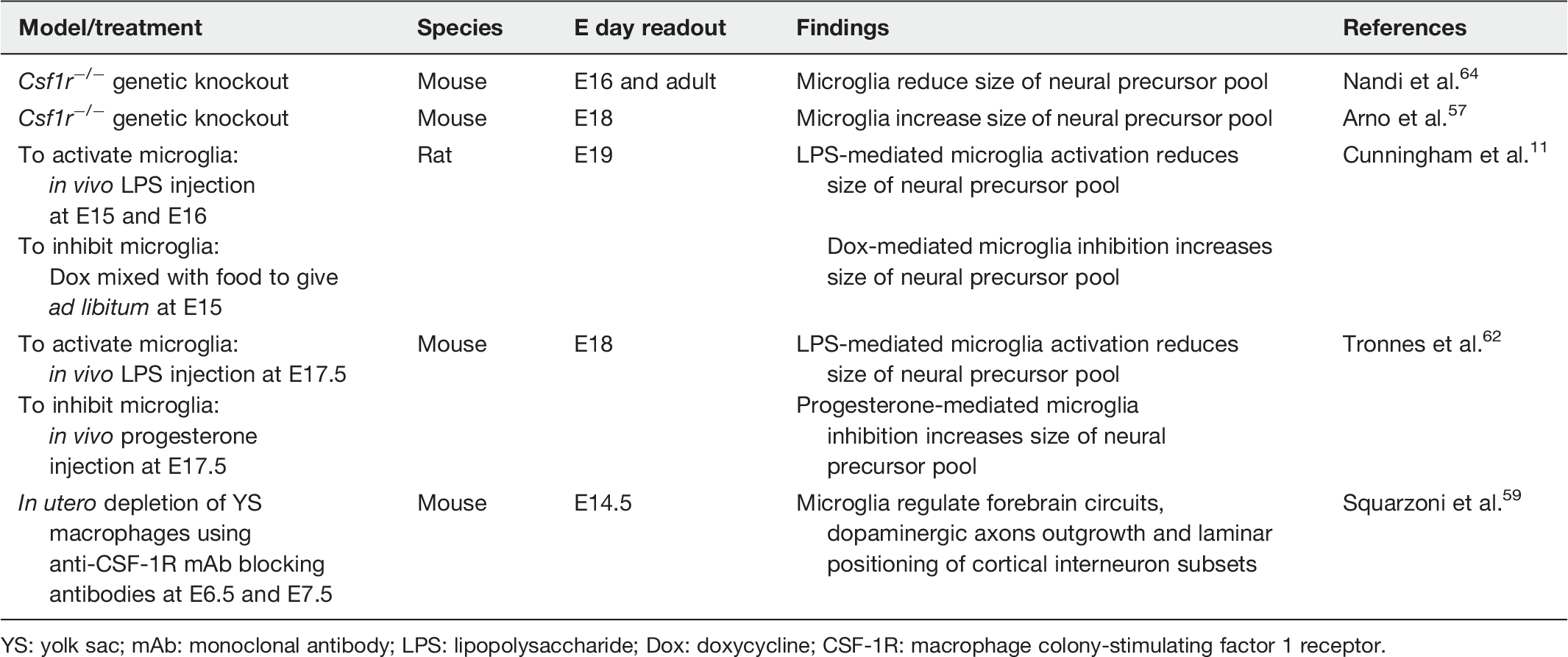
YS: yolk sac; mAb: monoclonal antibody; LPS: lipopolysaccharide; Dox: doxycycline; CSF-1R: macrophage colony-stimulating factor 1 receptor.
Genetic knockout models have also been useful to determine the role of microglia in neurogenesis. However, knockouts such as macrophage colony-stimulating factor 1 receptor (CSF-1R) and PU.1 do not solely target microglia as these are transcription factors required for monocytes and other tissue-specific macrophages, and their roles in development cannot be negated. Furthermore, deletion of these genes is embryonically lethal, making it difficult to irrefutably identify the role of microglia in any postnatal morphological or behavioral effects. Therefore, when examining data provided by these models, it is important to be aware of their limitations. A timely and valuable review on microglia depletion models discusses this in greater depth. 65 Nonetheless, interesting and valuable information regarding microglial functions in embryogenesis have emerged from these models. CSF-1R knockout mice which lack microglia display brain abnormalities such as reduced brain size with increased brain mass, olfactory bulb atrophy and increased ventricular size.64,66 The mice displayed expanded numbers of neural progenitors cells (nestin+, Pax6+, Tbr2+) in the neocortex compared to wild type animals, owing to the uncontrolled proliferation of neurogenic cells at both embryonic (E15.5) and postnatal (P20) time points. 64 This demonstrates that microglia potentially encourage neurogenesis by encouraging neural precursors to exit their proliferative mode. CSF-1R is also expressed by a small population of neural precursor cells. 64 However, selectively ablating CSF-1R in the neural precursor population only partially rescued the abnormality in the animal, indicating that microglia driven by CSF-1R signaling are required for development of the embryonic brain. 64 CSF-1R signaling in microglia also reduced the number of CTIP2+ and CUX1+ mature excitatory neuron subtypes and, therefore, implicates its role in neuronal differentiation. 64 A contradictory finding was reported in the CMVCre CSF-1Rflox/flox knockout model at E17.5 where deletion of CSF-1R gene resulted in reduced number of TBR2+ progenitors in the SVZ which was associated with reduced progenitor proliferation. 57 Although contradictory, nonetheless both studies demonstrate that microglia affect the number of neural progenitors during embryonic neurogenesis. The possibility of microglia influencing neurogenesis by encouraging neural precursors to exit the cell cycle is somewhat demonstrated in CSF-1R knockdown zebrafish where the absence of microglia in the retina resulted in retinal progenitor cells being in a continuous state of proliferation and delayed the onset of neurogenesis leading to dramatic gross abnormalities (microphthalmia). 67 The deficit of neurogenesis resulted from prolonged cell cycle periods (higher number of cycling cell pool) and/or disrupted cell cycle withdrawal in neural progenitor cells. When microglia reoccupied the retina, neurogenesis was partially recovered. 67 The effect of microglia on reducing neural progenitor numbers was also demonstrated in PU.1 knockout mice. 56 These mice also lack microglia and E12–E13 cortical precursor cultures from these mice showed significantly decreased Ki67+ proliferating cells. Although this did not appear to result in reduced neurons, astrocyte numbers were significantly reduced. This may affect overall neurodevelopment due to the imbalance of cell population number within the brain. Reintroducing microglia into these cultures restored precursor proliferation and rate of astrogenesis. 56 Another possible mechanism for regulating the size of the precursor pool is to induce cell death. Although yet to be observed in the neocortex, it has been demonstrated in the developing chick retina, interestingly an effect conferred by the trophic factor NGF 68 and TGF-β. 69
Beyond the effects of microglia on neural precursor pool size, embryonic microglia have also been shown to limit the outgrowth of dopaminergic axons into the forebrain and affect interneuron positioning in the laminar organization of the cortex. 59 These embryonic events affected dopaminergic innervation of the striatum and laminar organization of the cortex in newborn pups and, therefore, mark an embryonic activity of microglia that affects postnatal development of CNS circuitry. 59 Along with this finding, various other postnatal observations have been made that collectively trace the role of microglia throughout neurogenesis. Briefly, microglia have been shown to perform activity-dependent synaptic pruning at P5 70 and P15. 71 They also phagocytose apoptotic neurons in the cerebellum 60 and adult hippocampus, 72 promote developmental neuronal death60,61 and regulate neuronal migration to the olfactory bulb. 73 To conclude, the spatiotemporal similarities between microglia, neural precursors and neurons appear to have functional consequences throughout neurodevelopment. These physiological roles of microglia are increasingly being associated with neuropsychiatric and neurological disorders and have been reviewed by Prinz and Priller. 74 As the brain develops, microglia also appear to have phenotypes that shift from the embryonic and early postnatal phase to one that matches adult microglia.40,75 This may indicate the different functions of microglia during embryonic development compared to their housekeeping/inflammatory duties in the adult.
Conclusion
Numerous studies that report the effects of microglia throughout embryonic neurogenesis reinforces their role during development and better genetic and pharmacological animal models and fate-mapping have greatly advanced discoveries in this area. Microglia appear to keep the neural precursor pool size in check and influence neurogenesis during embryogenesis. What remains to be known is whether microglia have functions earlier in neurogenesis, an idea worth exploring due to their early colonization of the brain. Perturbing microglia during development leads to behavioral deficits and now more than ever, microglia dysregulation is implicated in the etiology for neuropsychiatric and neurodevelopmental disorders.
Footnotes
Authors’ contribution
Both CKT and SV wrote and reviewed the manuscript.
Acknowledgements
This work is supported by the Fundamental Research Grant Scheme [FRGS/2/2013/SKK01/UPM/02/15] awarded to SV from the Ministry of Education Malaysia. CKT is funded by the MyBrain 15 postgraduate scholarship programme, Ministry of Education Malaysia. We thank Prof. Dr Vidyadaran Menon and Dr Shinsmon Jose for comments on the manuscript.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.