
Abstract
Triple-negative breast cancer (TNBC) is the most aggressive type of breast cancer, with a high incidence of metastasis and chemoresistance. Epithelial-mesenchymal transition (EMT) is one of the molecular mechanisms that has been linked to the promotion of metastasis, and it can be promoted by several activators including the NF-κB signaling pathway. As a result, targeting EMT may be a potential strategy for treating TNBC. Pinostrobin is one of the important flavonoids found in the rhizome and rootlet of Boesenbergia rotunda (L.) Mansf. (fingerroot) that exhibits anticancer activities. However, the precise mechanism underlying the anticancer effect of pinostrobin on breast cancer remains unclear, and additional evidence is needed. In this study, the MCF-7 and MDA-MB-231 breast cancer cells were treated with various concentrations of pinostrobin. To determine the effect of pinostrobin on cell viability, an MTT assay was performed. Wound healing and Transwell chamber assays were conducted to examine the effect of pinostrobin on migration ability. RT-PCR was used to detect the expression of mRNA involved in NF-κB and EMT signaling pathways. The results revealed that low concentrations of pinostrobin did not affect cell viability, while higher concentrations produced an inhibitory effect on the viability of both cell lines. Pinostrobin also impeded migration and suppressed the expression of N-cadherin, a mesenchymal marker. Molecular docking analysis also suggested that the pinostrobin may target N-cadherin with higher binding affinity than IKK complex and NF-κB p65. These findings indicate that pinostrobin may serve as a potential treatment for TNBC.
Introduction
Breast cancer is the leading cause of cancer-related deaths among women worldwide and is classified into 4 main subtypes: luminal A, luminal B, HER2-positive, and triple-negative breast cancer (TNBC). While therapies for the first 3 subtypes have advanced significantly, TNBC remains the most challenging due to its aggressive nature, including invasiveness, high metastatic potential, and resistance to chemotherapy. 1 Current treatment options for TNBC primarily include chemotherapy, with immunotherapy still evolving. Emerging therapeutic strategies, such as fasting-mimicking diets targeting cancer stem cells, modulation of the tumor microenvironment, combination therapies, and enhancing chemosensitivity, show promise in improving TNBC treatment outcomes. 2
Epithelial-mesenchymal transition (EMT) is a key process contributing to breast cancer metastasis by enabling epithelial cells to acquire mesenchymal properties, thereby enhancing their invasiveness and migratory capacity.3,4 Activation of signaling pathways such as NF-κB has been identified as a major regulator of EMT, making it a potential target for therapeutic intervention. 5
In recent years, natural compounds from traditional herbs have gained attention as potential adjunctive therapies for cancer treatment due to their promising anticancer properties. One such compound is pinostrobin, a bioactive flavonoid found in the rhizomes and rootlets of Boesenbergia rotunda (fingerroot). 6 Pinostrobin has demonstrated antioxidant, anti-inflammatory, antibacterial, antiviral, and anticancer properties. Notably, toxicity studies have shown that it is non-toxic and non-genotoxic even at high doses, making it a promising candidate for cancer therapy.6,7
Pinostrobin has been reported to induce apoptosis, promote cell cycle arrest, and inhibit proliferation, invasion, and migration in various cancers.8,9 Specifically, in breast cancer, pinostrobin has shown no effect on the viability of the normal human breast cell line MCF10A but demonstrated inhibitory effects on the invasion and migration of TNBC cell lines such as T47D and MDA-MB-231. 10 Moreover, pinostrobin has been found to reduce the production of pro-inflammatory cytokines (TNF-α, IL-12, IL-1β) and modulate several signaling pathways. 11 However, the precise mechanisms through which pinostrobin exerts its anticancer effects, particularly in TNBC, remain unclear.
This study aims to investigate the anticancer potential of pinostrobin, focusing on its role in inhibiting EMT, cell proliferation, and the NF-κB signaling pathway, with the goal of contributing to the suppression of TNBC progression.
Methods
Reagents
(±)-Pinostrobin was obtained from Sigma-Aldrich (St. Louis, MO, USA). Pinostrobin (Figure 1) was dissolved in dimethyl sulfoxide (DMSO) and stored at −20°C.

Chemical structure of pinostrobin. 12
Cell Culture
Human breast cancer cells such as MCF-7 (ATCC HTB-22, RRID:CVCL_0031) and MDA-MB-231 (ATCC HTB-26, RRID:CVCL_0062) cells were purchased from the American Type Culture Collection (ATCC, USA). Cells were grown in Dulbecco’s Modified Eagle Medium (DMEM) supplemented with 10% fetal bovine serum, 1% penicillin, and streptomycin. Cultures were incubated at 37°C with 5% CO2.
MTT Assay
The effect of pinostrobin on the cell viability of MCF-7 and MDA-MB-231 cells was evaluated by 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT) reduction assay. MCF-7 and MDA-MB-231 cells were seeded at 7 × 103 cells/well in a 96-well plate and incubated at 37°C in a humidified atmosphere of 5% CO2 for 24 hours. The assay was performed for 24, 48, and 72 hours with varying concentrations of pinostrobin for 2 conditions such as low concentration (6.25, 12.5, 25, 50, and 100 µM) and higher concentration (250, 500, 750, and 1000 µM). Andrographolide 30 µM was used as a positive control (concentration was selected following a previous study 13 ). MTT was dissolved in phosphate buffer saline (PBS) to make a stock solution of 5 mg/ml. MTT stock solution was mixed with medium to make a final concentration of 0.5 mg/ml for the working solution. After treatment, the medium was discarded and 100 µl of MTT working solution was added to the cells in each well and incubated for the next 3 hours in the dark at 37°C in a humidified atmosphere of 5% CO2. Then, the medium was removed and 50 µl/well DMSO was added to the cells to solubilize the formazan crystals. The optical density (OD) was measured at 562 nm using a Varioskan Flash Multimode Reader (Thermo Fisher Scientific, Marietta, OH, USA). The cell viability of treatment groups was expressed as a percentage compared to the control.
Wound Healing Assay
The wound healing assay was used to determine the migration ability of the cells. When the cell density reached 90%, MCF-7 and MDA-MB-231 cells were seeded at 7 × 105 cells/well in a 6-well plate and incubated for 24 hours. Cells were treated with varying concentrations of pinostrobin (10, 20, and 50 µM). The cells were scratched using a pipette tip and grown in a culture medium with 1% FBS for 24 hours. Images of the wounds were monitored and captured using a 4× objective lens on an inverted microscope (Nikon Eclipse Ts2R-FL/Ts2R) with NIS-Elements software. The percentage of the recovered areas was measured using ImageJ software (1.53t, NIH).
Transwell Chamber Assay
The transwell chamber assay was performed using a 24-well transwell plate (0.8 µm pore, pc membrane, Corning Life Science, USA). Cells were treated with pinostrobin and seeded at 2 × 104 cells in 200 µl per well with serum-free medium into the upper chamber. DMEM supplemented with 10% FBS was placed into the lower chamber, then incubated at 37°C and 5% CO2 for 24 hours. Cells on the top of the plates were removed by using a cotton swab. The cells at the bottom of the plate which represent migrated cells were fixed with 100% methanol for 10 minutes and then stained with 0.05% crystal violet for 10 minutes before PBS rinsing. Images of migrated cells were captured under a 4× objective lens on an inverted microscope (Nikon Eclipse Ts2R-FL/Ts2R) with NIS-Elements software. The stained cells were counted using ImageJ software. 14
Real-Time Polymerase Chain Reaction (RT-PCR)
To detect the level of mRNA expression, RT-PCR was performed. MCF-7 and MDA-MB-231 cells were plated at a density of 1.0 × 106 cells/60 mm dishes in DMEM for 24 h. After incubation, cells were treated with different concentrations of pinostrobin (250, 500, and 1000 µM). Total RNA was extracted from the cell line by TRIzol® reagent (Invitrogen, USA) following the manufacturer’s protocol. The amount of RNA was quantified by measuring with NanoDrop One (Thermo Scientific). The RNA was converted into cDNA using the iScript™ Reverse Transcription Supermix at 25°C for 5 minutes, 46°C for 20 minutes, and 95°C for 1 minute, respectively. The genes include TNF-α, IκBα, NF-κB (RelA), E-cadherin (CDH1), N-cadherin (CDH2), and Twist were performed on equal amounts of cDNA (50 ng/2 µl). Forward and reverse primers were designed using NCBI/Primer-Blast and the sequences are listed in Supplemental Table S1, and RT-PCR was detected using the iTaq™ Universal SYBR® Green Supermix (Bio-Rad, Hercules, CA, USA) at 95°C for 10 minutes, followed by 45 cycles of 95°C for 10 seconds and 60°C for 60 seconds (denaturing, annealing and extension, respectively) and analyzed by system and analytical software (Bio-Rad). Samples were analyzed in duplicate manner and the expression ratio was normalized with GAPDH, the housekeeping gene.
Molecular Docking
Molecular docking was performed using AutoDock Vina 1.2.5 15 to predict the binding mode and affinity of pinostrobin to the targeted proteins. The three-dimensional (3D) structures of the IKKβ/NEMO (NFκB essential modulator) association complex, NF-κB p65, and N-cadherin were retrieved from the Protein Data Bank (PDB, https://www.rcsb.org/) using the PDB accession codes 3BRV, 1NFI, and 2QVI, respectively. These proteins were selected based on their crucial roles in EMT regulation, where NF-κB activation promotes EMT progression, and N-cadherin plays a key role in cell-cell adhesion and migration. The selection aimed to determine whether pinostrobin could inhibit EMT by suppressing NF-κB signaling or directly interacting with N-cadherin to disrupt EMT-associated processes. The Simplified Molecular Input Line Entry System (SMILES) format of pinostrobin (PubChem CID: 6950539) was obtained from the PubChem database (https://pubchem.ncbi.nlm.nih.gov). Its SMILES string was then converted into the PDB file format using UCSF Chimera. 16 All protein and ligand structures were converted into PDBQT file format using the AutoDockFR 1.0 software suite 17 prior to performing molecular docking. The dimensions of the grid box were set to 20 Å × 20 Å × 20 Å, with the grid center coordinates (x, y, and z) for each protein provided in Supplemental Table S2. The best docked pose (ie, the lowest binding energy) for each complex structure was selected, and the protein–ligand interactions were analyzed using the Discovery Studio Visualizer (BIOVIA, San Diego, CA, USA). The 3D binding mode was visualized using UCSF ChimeraX. 18
Statistical Analysis
All data were presented as the mean ± standard error of the mean (SEM) of 3 independent experiments. Statistical differences between the control and treatment samples were determined by using a two-tailed unpaired T-test from GraphPad Prism 9.0.0 (GraphPad Software Inc., La Jolla, Ca, USA). *P-values ≤ .05 were considered statistically significant.
Results
Effect of Pinostrobin on the Cell Viability
The effect of pinostrobin on the cell viability of MCF-7 and MDA-MB-231 breast cancer cells was examined using an MTT assay. The results demonstrated that pinostrobin did not affect the cell viability of both cell lines at the low concentrations (6.25, 12.5, 25, 50, and 100 µM) over 24, 48, and 72 hours (Figure 2). However, higher concentrations (250, 500, 750, and 1000 µM) of pinostrobin showed a trend to decrease the cell viability of both cells. The MCF-7 cell viability with treatment of 1000 µM pinostrobin at 24 and 48 h was reduced to 67% and 71%, respectively (Figure 3A and B). The IC50 values of pinostrobin on MCF-7 cells at 24, 48, and 72 hours were 1503.17, 1502.39, and 2866.35 µM, respectively. For MDA-MB-231, pinostrobin significantly reduced the cell viability to 78% of the control at concentrations of 500 µM for 24 hours, and approximately 64% and 50% at concentration of 750 and 1000 µM for 48 and 72 hours (Figure 3D-F). The IC50 values of pinostrobin on MDA-MB-231 cell lines at 24, 48, and 72 hours were 1316.84, 1035.88, and 1157.43 µM, respectively. On the other hand, 30 µM of andrographolide was used as a positive control and showed significant results in inhibiting both MCF-7 and MDA-MB-231 at an all-time range (Figure 3A-F). Furthermore, these results revealed that 750 and 1000 µM pinostrobin were effective in reducing cell viability of MDA-MB-231 cells for all 3 periods of time, while only 1000 µM was effective in MCF-7 cells at 24 and 48 hours.

The effect of pinostrobin on cell viability of MCF-7 and MDA-MB-231 cells at various concentrations (6.25, 12.5, 25, and 100 µM) at 24, 48, and 72 hours post treatment. Cell viability was assessed by MTT assay. MCF-7 and MDA-MB-231 cells were treated with pinostrobin for (A and D) 24 hours, (B and E) 48 hours, and (C and F) 72 hours, respectively. There were no statistically significant differences between control and treatments of pinostrobin for all cell lines. Data are represented as means ± SEM compared to the DMSO control (n = 3).
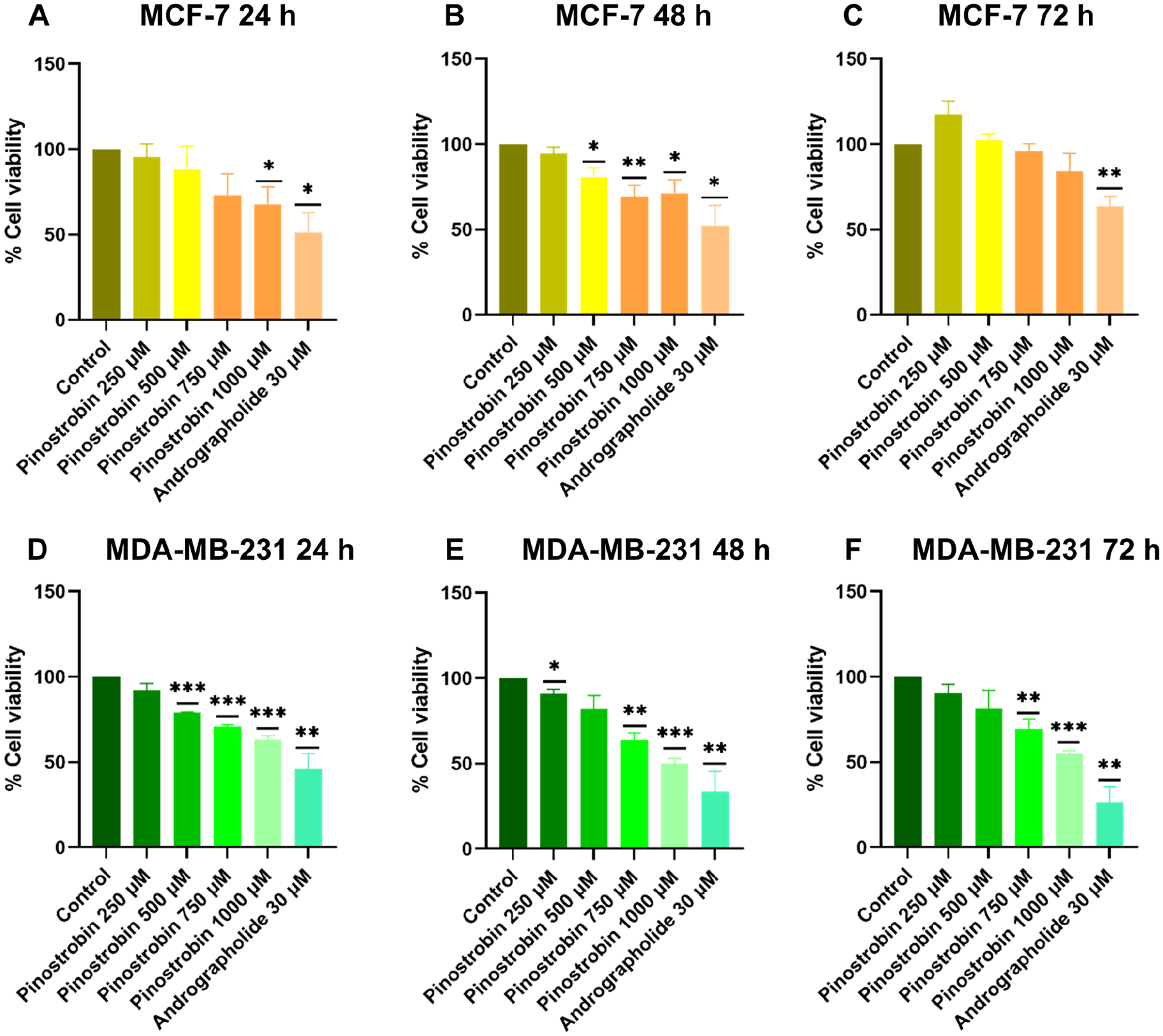
Pinostrobin decreased cell viability of MCF-7 and MDA-MB-231 cells at various concentration (250, 500, 750, and 1000 µM) for 24, 48, and 72 hours. Cell viability was assessed by MTT assay. MCF-7 and MDA-MB-231 cells were treated with different concentration of pinostrobin and 30 µM of andrographolide (positive control) for (A and D) 24 hours, (B and E) 48 hours, and (C and F) 72 hours, respectively. The results were statistically significant compared to DMSO control. Data are means ± SEM compared to DMSO control (n = 3).
Effect of Pinostrobin on Cell Migration
The effect of pinostrobin on migration was examined by using wound healing and transwell chamber assays. In wound healing assay, a wound was created on both cell lines and treated with 10, 20, and 50 µM pinostrobin for 24 hours. These concentrations were selected based on the MTT assay results, which showed that they did not significantly affect cell viability (Figure 2). This ensured that any observed reduction in migration was not due to cytotoxicity but rather a direct effect of pinostrobin on cell motility. The wound area was measured before and after treatment, and the difference in wound closure was calculated as a percentage compared to the untreated control. The results showed that pinostrobin decreased the percentage of wound healing in both cells, with a more pronounced effect observed in MCF-7 cells. Treatment with 20 µM of pinostrobin resulted in a 55% reduction in wound closure in MCF-7 cells, while it produced a 21% inhibition in MDA-MB-231 cells, as shown in Figure 4.

Pinostrobin suppressed cell migration on MCF-7 and MDA-MB-231 cells. Both cell lines were treated with 10, 20, and 50 µM pinostrobin for 24 hours. Pinostrobin produced significant inhibition on (A) MDA-MB-231 and (B) MCF-7 cell lines compared to untreated control. Data are presented as mean ± SEM from 3 independent experiments performed in triplicate. The statistical significance compared to non-treatment control.
In the transwell assay, MDA-MB-231 cells were cultured in transwell chambers with different pinostrobin concentrations (20 and 50 µM) selected based on the wound healing assay results and then allowed to migrate for 24 hours. As depicted in Figure 5A, pinostrobin treatment inhibited the migration of MDA-MB-231 cells in a dose-dependent manner. Treatment with 20 and 50 µM pinostrobin resulted in approximately 26% and 35% inhibition of migration in MDA-MB-231 cells, respectively. Conversely, MCF-7 cells were also cultured in transwell chambers with the same concentrations of pinostrobin, but only a small number of cells migrated, as shown in Figure 5B.
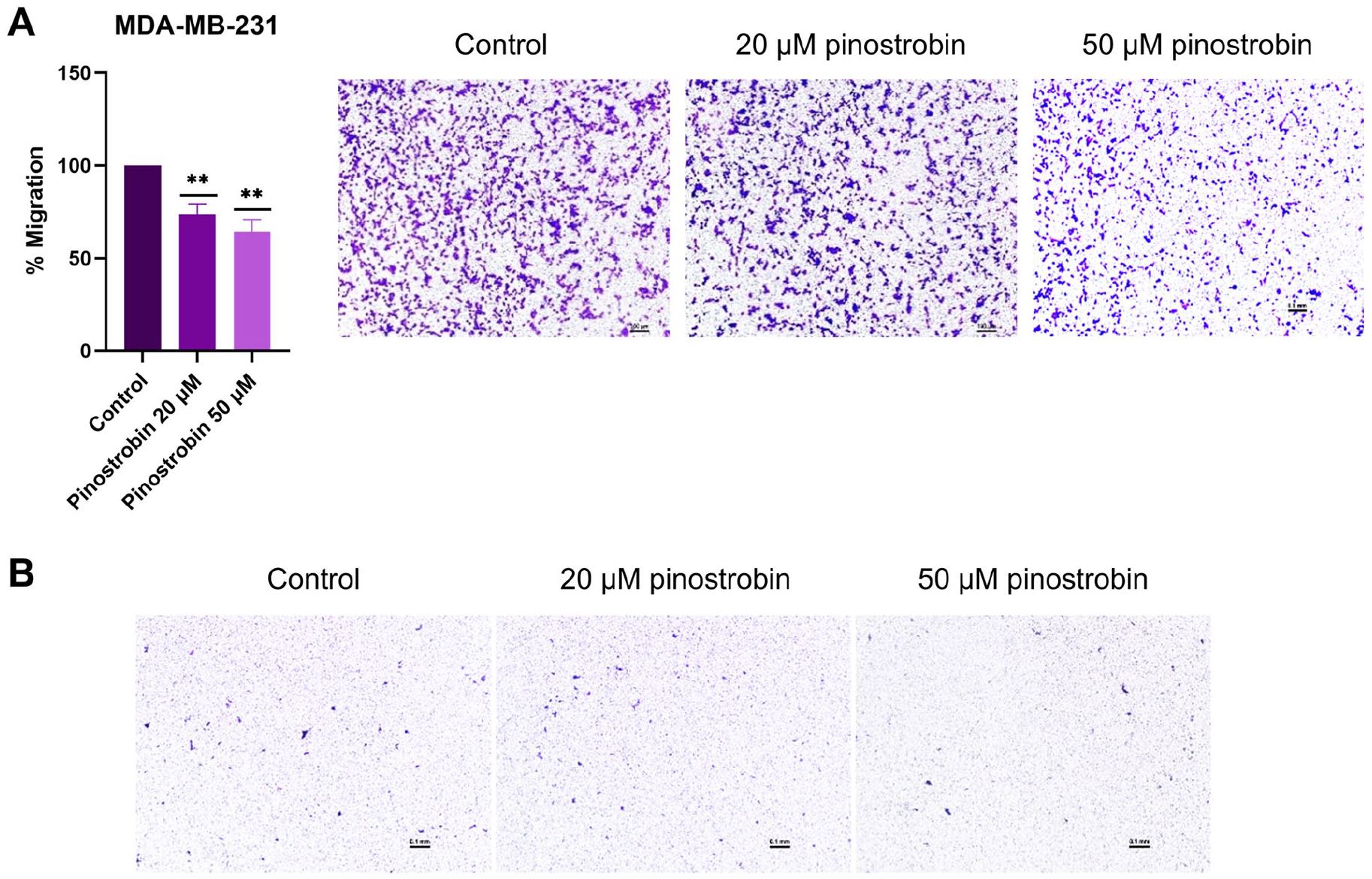
Pinostrobin inhibits migration of MDA-MB-231 cells by transwell chamber assay. (A) The percentage MDA-MB-231 cells migration after treatment with 20 and 50 µM of pinostrobin. (B) MCF-7 migrated cells. Data are means ± SEM compared to the untreated control from a minimum of 3 independent experiments performed in duplicate.
Effect of Pinostrobin on NF-κB Signaling Pathway
As the activation of the NF-κB signaling pathway is involved in promoting the EMT process, the mRNA expressions of TNF-α, IκBα, and NF-κB after treatment with varying concentrations of pinostrobin on MCF-7 and MDA-MB-231 cells were examined by RT-PCR. The concentrations of pinostrobin used ranged from 250 to 1000 µM and were selected according to the effect on cell viability from the MTT assay. The results showed that in MCF-7 cells, the mRNA level of TNF-α was significantly increased by 250 and 1000 µM pinostrobin. However, 500 µM pinostrobin produced a small effect on the cells. In MDA-MB-231 cells, only 1000 µM of pinostrobin had a minimal effect on the expression of the TNF-α gene (Figure 6A). The expression of gene IκBα was significantly increased by 250 µM pinostrobin in MCF-7 cells and by 500 µM in MDA-MB-231 cells (Figure 6B). On the other hand, pinostrobin had a minimal effect on NF-κB mRNA level in both cell lines, and there was no statistical significance, although it seemed to be reduced in MDA-MB-231 cells (Figure 6C) Interestingly, despite our initial hypothesis that pinostrobin would inhibit this pathway, our results did not show a significant reduction in NF-κB mRNA levels in either MCF-7 or MDA-MB-231 cells. This suggests that the NF-κB pathway may not be the primary target of pinostrobin in these cell lines and that its anticancer effects may be mediated through alternative mechanisms.

The effect of pinostrobin on the gene expression by RT-PCR. The mRNA expression of (A) TNF-α, (B) IκBα, and (C) NF-κB in both MCF-7 and MDA-MB-231 cells post treatment with varies concentration of pinostrobin (250, 500, and 1000 µM). Data are means ± SEM compared to the untreated control (n = 3).
Effect of Pinostrobin on the EMT Markers
To determine the effect of pinostrobin on EMT markers, the gene expression of EMT markers such as E-cadherin (CDH1), N-cadherin (CDH2), and twist were examined on MCF-7 and MDA-MB-231 cells after treatment with vary concentrations of pinostrobin (250, 500, and 1000 µM). The results showed that the levels of CDH1 and Twist genes were similar to the untreated control after being treated with pinostrobin (Figure 7A). There were no statistically significant differences observed between the control and the treatments of pinostrobin in both cell lines (Figure 7A and C). However, at 500 and 1000 µM, pinostrobin treatment downregulated CDH2 gene and N-cadherin expression in both cell lines, especially in MDA-MB-231 cells (Figure 7B). These results indicated that pinostrobin was able to suppress the expression of mesenchymal markers at mRNA levels.

The effect of pinostrobin on mRNA levels of EMT markers detected by RT-PCR. mRNA relative expression levels of the gene (A) CDH1 encode for E-cadherin, (B) CDH2 encode for N-cadherin, and (C) twist in MCF-7 and MDA-MB-231 cells untreated and treated with different concentration of pinostrobin (250, 500, and 1000 µM). Data are means ± SEM compared to the untreated control (n = 3).
Molecular Docking of Pinostrobin with Targeted Proteins
Molecular docking was performed to identify the potential binding modes and affinities of pinostrobin with key proteins involved in the signaling pathway. The docking results for pinostrobin against 3 target proteins—IKK complex, NF-κB, and N-cadherin—are summarized in Table 1. The rationale for selecting these proteins was based on their crucial roles in EMT and breast cancer progression. The IKK complex and NF-κB p65 were selected due to their involvement in NF-κB-mediated EMT activation, a key signaling pathway that promotes migration and metastasis. N-cadherin was chosen as a target due to its role in cell-cell adhesion and EMT regulation, where its upregulation is associated with increased cancer cell invasion and metastasis. Given the significance of these proteins in EMT, docking analysis was conducted to predict whether pinostrobin could inhibit EMT by disrupting NF-κB activation or directly interacting with N-cadherin. A more negative binding energy value indicates a higher binding affinity of the ligand to the protein. The docking energy of pinostrobin with the IKK complex is −5.822 kcal/mol. In this complex, pinostrobin interacts with Trp739 through 2 conventional hydrogen bonds (H-bond) and a pi-pi T-shaped interaction (Figure 8A). Additionally, non-classical H-bonds at Thr735 and Ser740, pi-alkyl interactions at Phe92 and a pi-anion interaction at Glu99 contribute to the binding of pinostrobin within the binding pocket of IKK complex. Whereas the binding energy of pinostrobin with NF-κB p65 is −5.719 kcal/mol, which is similar to its binding energy with the IKK complex. In the docked conformation, pinostrobin interacts with the residues of NF-κB p65, including Gln119 and Arg124 through H-bonds, Lys122 via an alkyl bond, and Lys37 via a pi-alkyl interaction (Figure 8B). Interestingly, the pinostrobin–N-cadherin complex exhibited the highest binding energy (−6.525 kcal/mol) among the 3 proteins, suggesting that pinostrobin has a preferential binding affinity for N-cadherin compared to the other 2 proteins. It was found that the residues Leu145, Arg146, and Glu167 of N-cadherin play a critical role in binding with pinostrobin through pi-alkyl, H-bond and amide-pi stacked interactions, respectively (Figure 8C).
Docking scores of pinostrobin against the IKK complex, NF-κB p65, and N-cadherin, along with the crucial binding residues of the targeted proteins interacting with pinostrobin.


The 2D-interaction diagrams of pinostrobin in complex with (A) IKK complex, (B) NF-κB p65, and (C) N-cadherin obtained from molecular docking.
Discussion
EMT is one of the important factors that promote cancer progression and aggressiveness in many types of cancers, including breast cancer. 4 Various regulators of EMT include signaling pathways, tumor microenvironment, EMT transcription factors, epigenetics, and post-translational modifications. 19 These upstream regulators can activate EMT and enhance the motility and migration ability of cancer cells. Targeting this process may have the potential to suppress migration in breast cancer, especially in the TNBC subtype.
Nowadays, many natural products from traditional plants have become more popular as treatments for cancer among patients due to their cheaper price and fewer side effects compared to the medication prescribed by the hospital. Boesenbergia rotunda (L.) Mansf. (Fingerroot) is one of the plants that has gained increasing attention. It contains several bioactive compounds including pinostrobin. According to a few studies, pinostrobin can induce apoptosis, promote cell cycle arrest, and inhibit the cancer cells proliferation, invasion, and migration including in breast cancer. 9
Pinostrobin has been reported to exert cytotoxicity against various cancer cell lines, including leukemia, melanoma, cervical, and breast cancer cells.9,10,20,21 In a previous study, low concentrations of pinostrobin (0.01-20 µM) did not inhibit the viability of T47D and MDA-MB-231 breast cancer cells and did not affect MCF10A, which is a normal human breast cell, 10 while increasing concentrations (100-1500 µM) of pinostrobin reduced viability of T47D cells in another study. 22 However, no previous studies have reported the inhibitory effect of pinostrobin at higher concentrations on the MCF-7 and MDA-MB-231 cells. MCF-7 cell is a common breast cancer cell line that has been used in several research. It represents ER- and PR-positive type of breast cancer and is known as luminal A. 23 MDA-MB-231 is a cell that represents TNBC. It has been chosen for various in vitro models because it lacks hormone receptors and has an aggressive phenotype.24,25 MDA-MB-231 cells also demonstrated typical EMT associated with cancer progression. 26 In the present study, we demonstrated that pinostrobin exerted an anticancer effect on both MCF-7 and MDA-MB-231 cells by suppressing cell viability at higher concentrations (Figure 3). Low concentrations of pinostrobin did not show any significant effect on the viability of either cell line, similar to a previous study. 10 Increasing concentrations of pinostrobin, decreased the viability of both cell lines, especially at 750 and 1000 µM, at 24 and 48 hours. However, it should be noted that the cells seemed to regrow after 48 hours. In addition, to assess the test validity, andrographolide was used as a positive control as it has an effective effect on breast cancer cell lines including inhibiting the cell viability, decreasing cell migration, suppressing several signaling molecules, and NF-κB-dependent matrix metalloproteinase-9 expression.13,27 -29
Migration is a major characteristic of aggressive types of cancer, including TNBC. It can be promoted by the EMT process resulting in metastasis and a higher mortality rate in patients. 30 To examine the migration ability of cancer cells, wound healing and transwell assays are commonly used in vitro. In a previous study, low concentrations of pinostrobin (5, 10, and 20 µM) inhibited the migration and invasion abilities of T47D and MDA-MB-231 cells but did not affect MCF10A normal breast cells by performing both methods. 10 In this study, we determined the migration ability of MCF-7 and MDA-MB-231 cells after treatment with pinostrobin (10, 20, and 50 µM) for 24 hours using wound healing and transwell assays. In the wound healing assay, the wound recovery of MCF-7 cells showed less healing activity than that of MDA-MB-231 cells (Figure 4). As MCF-7 cell lines represent ER-positive breast cancer, which is a less aggressive type and has a lower rate of migration, they showed less healing activity than the aggressive MDA-MB-231 cell line. Pinostrobin significantly inhibited the migration of both cell lines at low concentrations. The migration activity of MDA-MB-231 cells was also confirmed by transwell chamber assay. These results were similar to those of the wound healing assay. Pinostrobin significantly inhibited the cell migration of MDA-MB-231 breast cancer cells (Figure 5). In the example tests on MCF-7 treated with pinostrobin, only a small number of cells can be observed. This outcome is expected, as MCF-7 cells are a non-invasive breast cancer cell line with inherently low migratory capacity due to limited expression of key motility-related molecules. Even in the control assay, very few MCF-7 cells migrated, consistent with their known non-invasive nature. This finding aligns with the study by Gest et al, 31 which reported minimal migration in MCF-7 cells compared to the highly invasive MDA-MB-231 cells. These differences highlight the intrinsic variation in cell motility between the 2 breast cancer subtypes.
One of the upstream pathways that can activate the EMT program is NF-κB.5,32 The breast cancer tumor microenvironment is highly inflamed and composed of many molecules such as immune cells, growth factors, cytokines, and extracellular matrix molecules. These components contribute to promoting EMT. NF-κB is a major transcription factor that links cancer to inflammation. It promotes inflammation, proliferation, survival, and metastatic potential of cancer cells. NF-κB forms a complex with inhibitor subunits of IκB-α, IκB-β, IκB-γ as an inactive form located in the cytoplasm. This complex can be released by various activators that cause IκB phosphorylated by the IκB kinase (IKK) complex, which leads to IκB degradation and translocation of NF-κB into the nucleus. In the nucleus, NF-κB binds to DNA to promote transcriptional activity and induce inflammation, invasiveness, metastasis, and EMT-transcription factor.33 -35 Based on this knowledge, this study chose this pathway to examine the possible mechanism underlying the effects of pinostrobin. However, our result demonstrated that the NF-κB gene expression showed no significant changes in either cell line, with only a slight decrease observed in MDA-MB-231 cells (Figure 6C). This could be due to factors such as pinostrobin concentration, treatment duration, or experimental conditions that may not have been optimal for detecting significant effects on this pathway. Our findings suggest that anticancer activity of pinostrobin may be mediated through alternative molecular mechanisms. Previous studies have shown that pinostrobin targets the Notch signaling pathway in lung carcinoma 36 and promotes melanogenesis via the cAMP/PKA and p38 MAPK pathways. 21 These findings indicate that the effects of pinostrobin may vary depending on cell type and context, highlighting the need for further investigation into its precise molecular targets.
However, there are some reports on the biphasic effects of flavonoids at higher concentrations on the expression of genes that should be considered.37,38 As shown in Figure 6A, TNF-α expression exhibited a biphasic response to pinostrobin, increasing significantly at 250 µM, showing no significant change at 500 µM, and rising again at 1000 µM. This pattern aligns with the dual role of TNF-α, which can promote tumor progression at low concentrations but induce apoptosis at higher levels. 37 These findings suggest that pinostrobin may trigger an initial inflammatory response, followed by apoptotic effects at higher doses, warranting further investigation into its dose-dependent mechanisms. A similar biphasic effect has been observed in other flavonoids (eg, apigenin, quercetin, chrysin and diosmetin), as reported by Xi et al, 38 where low concentrations (~10 µM) promoted proliferation, while higher doses (>50 µM) induced cell death. In our study, TNF-α expression increased at 250 and 1000 µM but not at 500 µM, suggesting a comparable dose-dependent mechanism. This supports the idea that flavonoids, including pinostrobin, exert varied cellular effects based on concentration. We also provided evidence for the EMT marker genes after treatment with pinostrobin. The genes are CDH1 (E-cadherin), CDH2 (N-cadherin), and twist (EMT-TF). Twist is an EMT transcriptional factor that regulates the EMT. It is activated by signaling pathways including NF-κB pathway. E-cadherin represents an epithelial marker that is suppressed when the EMT process is activated, whereas N-cadherin, a mesenchymal marker, is highly expressed. 39 To reverse the EMT process, the treatment may have to be able to reverse these 2 markers. The results showed that pinostrobin does not affect the mRNA level of E-cadherin and twists in both MCF-7 and MDA-MB-231 cells. Interestingly, pinostrobin was able to downregulate the N-cadherin gene in MDA-MB-231 cells, and a small decreasing trend in MCF-7 cells. Therefore, the RT-PCR results indicated that 500 µM pinostrobin can upregulate IκBα, the inhibitor of NF-κB, it may suppress IKK phosphorylation or disrupt the ubiquitination of IκB. Increasing of IκB is essential to inhibit the translocation of NF-κB. Pinostrobin may provide an effective effect on migration activity due to its effect on wound healing and transwell assays. This effect was further supported by RT-PCR results showing the downregulation of CDH2 (Figure 7B), which encodes N-cadherin, a key mesenchymal marker associated with EMT. Inhibition of N-cadherin may improve the cell-cell junctions and decrease the mesenchymal phenotype, cell migration, proliferation, and survival of cancer cells. There are a few studies that have proposed the N-cadherin antagonists as cancer therapeutics. 40
Furthermore, we evaluated the binding affinity of pinostrobin to the targeted proteins through molecular docking to support our proposed mechanism, which were selected based on their crucial roles in EMT initiation and metastasis. Pinostrobin was docked into the structures of the IKK complex, NF-κB p65 and N-cadherin. Our results revealed that pinostrobin exhibited the highest binding energy with N-cadherin compared to the other target proteins (Table 1). Specifically, pinostrobin showed interactions with Leu145, Arg146, and Glu167 at the binding site of the N-cadherin structure, with a binding energy of −6.525 kcal/mol (Figure 8C). This suggests strong potential for pinostrobin to interfere with N-cadherin-mediated cell adhesion and migration, which are essential processes in EMT-driven metastasis. A previous docking study indicated that ADH-1 (a N-cadherin antagonist) bound in a similar region to pinostrobin and interacted with multiple residues of N-cadherin, particularly Glu167. Additionally, an in vitro study demonstrated that ADH-1 significantly inhibited colorectal cancer cell migration. 41 The similarity between pinostrobin and ADH-1 in their docking profiles further supports our hypothesis that pinostrobin may act as an N-cadherin inhibitor with therapeutic potential. The computational findings are consistent with our in vitro experiments, which demonstrated that pinostrobin downregulated CDH2 (N-cadherin) mRNA expression and significantly inhibited cancer cell migration in both wound healing and transwell assays. These results provide evidence that the anti-migratory effects of pinostrobin are primarily mediated through direct N-cadherin inhibition rather than NF-κB pathway suppression, as RT-PCR analysis showed no significant inhibition of NF-κB p65 expression. Thus, our study provides both computational and experimental validation that pinostrobin exerts its anticancer effects mainly by targeting N-cadherin. These findings support the potential therapeutic application of pinostrobin as an N-cadherin inhibitor to suppress EMT-driven metastasis in breast cancer.
There are several limitations to this study that should be acknowledged. Firstly, the use of higher concentrations of pinostrobin was restricted by the toxicity of DMSO. Exceeding the recommended DMSO concentration can lead to non-specific cytotoxic effects, which may confound the interpretation of results. Secondly, the solubility of pinostrobin in culture media is another important factor. At higher concentrations, the solubility of the compound may limit its bioavailability, potentially affecting its effective concentration in the cells and influencing the observed effects. Additionally, the regrowth of cells observed after 48 hours suggests that the anticancer effect of pinostrobin may not be sustained over longer periods. This regrowth may be due to a delayed or partial inhibition of cell proliferation, or it could indicate the activation of compensatory survival mechanisms within the cancer cells. Future studies employing different dosing schedules, extended treatment durations, or combination therapies could help clarify whether effects of pinostrobin can be prolonged and whether its efficacy can be enhanced.
Conclusion
In conclusion, this study revealed that pinostrobin, a natural compound commonly found in Krachai, exerts anticancer effects on breast cancer cells. It significantly reduces cell viability at high concentrations and inhibits cell migration by suppressing N-cadherin, a marker of EMT (Figure 9). Molecular docking also identified potential targets for pinostrobin, showing a stronger binding affinity toward N-cadherin compared to the IKK complex and NF-κB p65. We believe our findings will guide further studies on pinostrobin’s anticancer activity in both in vitro and in vivo models. These results provide valuable data on the effects of pinostrobin against breast cancer cells, supporting further research toward developing new anticancer drugs.

Summary diagram illustrating the anticancer effect of pinostrobin on breast cancer cells through suppression of N-cadherin, an EMT marker.
Supplemental Material
sj-docx-1-ict-10.1177_15347354251341438 – Supplemental material for Anticancer Effect of Pinostrobin on Human Breast Cancer Cells Through Regulation of Epithelial Mesenchymal Transition
Supplemental material, sj-docx-1-ict-10.1177_15347354251341438 for Anticancer Effect of Pinostrobin on Human Breast Cancer Cells Through Regulation of Epithelial Mesenchymal Transition by Pimrapat Jongjang, Sutharinee Likitnukul, Somrudee Reabroi, Supachoke Mangmool, Bodee Nutho and Darawan Pinthong in Integrative Cancer Therapies
Footnotes
Author Contributions
Experimental design, data analysis, writing and original draft preparation: PJ, SL, BN, DP; Conceptualization and supervision: DP, SM, SL; Experiments: PJ, SL, SR, BN; Resources: DP and SM; Funding acquisition, reviewing and editing: DP. All authors read and approved the final version of the manuscript.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was partially supported by the Department of Pharmacology, Faculty of Science, Mahidol University. Andrographolide was obtained from ARDA (PRP6605030320) through Assoc. Prof. Dr. Rungnapha Saeeng of the Department of Chemistry, Faculty of Science, Burapha University. This work was supported by the Central Instrument Facility (CIF) and Center of Nanoimaging (CNI), Faculty of Science, Mahidol University, for the use of research instruments and facilities.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.