
Abstract
The radioprotective property of 50 mg/kg body weight jamun (Syzygium cumini) extract was studied in the cultured splenocytes of mice exposed to 0, 0.5, 1, 2, 3, or 4 Gy of γ-radiation. The spleens of irradiated mice were removed aseptically and the splenocytes were extracted from the individual spleens and cultured. The micronuclei were prepared 72 hours after irradiation in binucleate splenocytes by blocking cytokinesis with cytochalasin-B. Irradiation of mice resulted in a dose-dependent elevation in the micronucleated splenocytes. The exposure of mice not only elevated splenocytes bearing one micronucleus but also cells bearing 2 and multiple (>2) micronuclei indicating induction of complex DNA damage after irradiation. Oral treatment of mice with 50 mg/kg body weight of jamun leaf extract protected against the radiation-induced micronuclei formation. Jamun extract also protected against the formation of 2 and multiple micronuclei indicating repair or inhibition of complex DNA damage. The assessment of lipid peroxidation in mice brain homogenate has indicated a concentration dependent inhibition of lipid peroxidation by jamun extract. Studies in a cell free system revealed that jamun extract inhibited the formation of OH, O2−, DPPH, and ABTS+ free radicals in a concentration dependent manner. Our study demonstrates that jamun extract protected mice against the radiation-induced DNA damage and inhibition of radiation-induced free radical formation may be one of the mechanisms of radioprotection.
Introduction
Human exposure to ionizing radiation has increased because of its use for medical diagnosis and therapy, industrial applications, nuclear power generation, atomic weapons testing, deployment of nuclear arsenals, atomic detonations for peaceful purposes, nuclear accidents, stellar radiations, and air and space travel. 1 Irradiation has been known to cause genomic instability and subsequently mutagenesis and carcinogenesis. 2 However, the potent mutagenic effect of ionizing radiation was first demonstrated more than 84 years ago by Muller. 3 Mutagens can induce genomic changes by targeting DNA directly or/and indirectly, by binding to proteins involved in the maintenance of genome integrity. Ionizing radiation—including X-rays, γ-rays, α− or β−particles, nuclear fission products, and radionucleides—induces various genetic damage and mutations, and is an established carcinogen. Ionizing radiation induces a variety of DNA lesions by direct and indirect interactions, including DNA base alterations, DNA–DNA and DNA–protein cross-links, single- and double-strand breaks and immunosupression, micronucleus formation, sister chromatid exchanges, chromosomal aberrations, and genomic instability.4-7 The induction of chromosomal aberrations depends on the stage of cell cycle during radiation exposure. Irradiation of cells in G1 and early S phase causes chromosome aberrations, whereas chromatid aberrations are produced during G2 or mid and late S phase. 8
The cytogenetic damage induced by physical or chemical agents has been traditionally evaluated using chromosome aberration analysis after experimental challenge until 1973, when Heddle and Carrano 9 devised a test to detect micronuclei in human peripheral blood lymphocytes as a marker for chromosomal damage. This was followed by the micronucleus test in erythrocytes in mouse bone marrow. 10 The micronucleus test has become very popular among investigators owing to its simplicity, speed, and sensitivity. In addition, it has an advantage over chromosome aberration analysis as a higher number of cells could be scored in relatively less time, which precluded the utilization of services of highly skilled personnel that is essential for chromosome aberration analysis. 11 Owing to easy application, the micronucleus assay immediately found its use in screening of environmental toxicants, identification of individuals more sensitive to a clastogenic exposure, 11 routine screening of radiation workers, 11 prediction of radiocurability of tumors, 12 and assessment of genomic damage during prenatal development. 13 The use of cytochalsin-B by Fenech and Morley 14 allowed screening of micronuclei in first division binucleate cells as the presence of cytochalsin-B inhibited actin polymerization and formation of cytoplasmic bridge in the daughter cells. 15 This test has been especially useful in cultured mammalian cells, including human peripheral blood lymphocytes. Ionizing radiation induces DNA damage that in turn is converted to chromosome damage and subsequently to micronuclei after one cell division. 16 Therefore, assessment of micronuclei is an appropriate indicator to evaluate the radioprotective activities of pharmacological agents in vitro as well as in vivo.
Drugs that can differentially protect surrounding normal tissue and/or sensitize hypoxic tumor cells are desirable to improve the therapeutic index during radiotherapy of tumors. However, a radioprotector will also be useful during nuclear accidents, inadvertent release of radioactivity, space and air travel, as well as nuclear terror attacks. 1 Several chemical agents have been screened for their radioprotective potential since Patt et al 17 first demonstrated that cysteine, an amino acid containing sulfydryl group, protected rats against the radiation-induced lethality. However, a majority of the sulfydryl-containing compounds provided protection at very high, toxic dose, and their application for human use has been always limited. 18 Therefore, it is necessary to screen new protectors in the hope to find a suitable agent that is cheap, nontoxic at protective dose in a wide dose range, orally administrable, rapidly absorbable, provides a higher degree of protection, possesses a reasonably good dose reduction factor, and can act through multiple mechanisms. Since plants are more biochemically similar to human tissue than synthetic chemicals, it is reasonable to screen plant or herbs for their radioprotective properties. 1
Jamun, Syzygium cumini (L.) Skeels (family Myrtaceae) is a medium sized to large tree, and it has been attributed to posses several medicinal properties in the folklore system of medicine. 19 The stem bark is astringent, sweet, refrigerant, carminative, diuretic, digestive, antihelminthic, febrifuge, constipating, stomachic, and antibacterial. The fruits and seeds are used to treat diabetes and splenopathy. Various studies have indicated that jamun has antioxidant, anti-inflammatory, antibacterial, antidiabetic, and gastroprotective properties.20-22 The leaves of the plant have been reported to reduce radiation-induced DNA damage in cultured human peripheral blood lymphocytes and to increase mouse survival.23,24 Similarly, the seed extract of jamun has been found to protect against radiation-induced lethality in mice. 25 The aim of the present study was to evaluate the radioprotective activity of leaf extract of S. cumini on the radiation-induced micronucleus formation in cultured mouse splenocytes exposed to different doses of γ-radiation.
Materials and Methods
Chemicals
Phytohemagglutinin (PHA) was procured from Difco Laboratories (Detroit, MI), whereas cytochalasin-B, RPMI, fetal calf serum (FCS),
Preparation of Extract
The fresh mature leaves of S. cumini (synonym Eugenia cumini Druce) were collected locally during the months of April and May and were identified by Dr G. K. Bhat (Department of Botany, Poorna Prajna College, Udupi, Karnataka, India). The leaves were cleaned, shade dried, and powdered in a mixer, and the extract was prepared as described earlier. 26 Briefly, the leaf powder was extracted in petroleum ether and chloroform and finally in 1:1 dichloromethane and methanol at 50°C to 60°C using a Soxhlet apparatus. The extract was cooled and concentrated by evaporating its liquid contents in vacuo and freeze dried. The extract was stored at −70°C until further use. Henceforth the extract of S. cumini will be called SCE.
Preparation of Drug and Other Solutions
SCE was dissolved in 1% w/v carboxymethyl cellulose (CMC) in sterile physiological saline. The animals were given 0.01 mL/g body weight of CMC or SCE through oral gavage (p.o.) consecutively for 5 days before irradiation. Cytochalasin B was dissolved in DMSO at a concentration of 10 mg/mL, stored at −80°C, and diluted with the RPMI immediately before use. Concanavalin-A was dissolved in phosphate buffered saline (PBS) and filter sterilized.
Animal Handling
Animal care and handling were performed according to guidelines issued by the World Health Organization (Geneva, Switzerland), the Indian National Science Academy, New Delhi, India and the “Guide for the Care and Use of Laboratory Animals” (NIH publication #86-23, revised in 1985). Ten- to 12-week old male Swiss albino mice weighing 30 to 36 g were selected from an inbred colony maintained under controlled conditions of temperature (23 ± 2°C), humidity (50% ± 5%), and light (12 hours of light and dark cycle). Animals had free access to sterile food and water. Four to 6 animals were housed in a polypropylene cage containing sterile paddy husk (procured locally) as bedding throughout the experiment. The Animal Ethical Committee at the authors’ institution approved the study.
Radioprotective Effect
The radioprotective effect of 50 mg/kg body weight SCE was taken and studied on the radiation-induced micronucleus formation in cultured splenocytes of mice exposed to different doses of γ-radiation, where the animals were divided into the following groups:
CMC + Irradiation: The animals of this group were treated with 0.01 mL/g body weight of CMC before exposure to different doses of γ-radiation.
SCE + irradiation: The animals of this group were administered 50 mg/kg body weight of SCE p.o. once daily for 5 consecutive days before exposure to different doses of γ-radiation.
Irradiation
One hour after the last administration of CMC or SCE, animals from both groups were exposed to 0, 0.5, 1, 2, 3, or 4 Gy 60 Co γ-radiation from a Tele cobalt therapy source (Theratron, Atomic Energy Agency, Ontario, Canada) at a dose rate of 1.66 Gy/min.
Splenocyte Culture
The animals were humanely killed by cervical dislocation 30 minutes post irradiation. The animals were thoroughly wiped with sterillium disinfectant (Bode Chemie, Hamburg, Germany), and their abdominal cavity was opened with the help of sterile scissors and forceps under aseptic conditions. The spleens of the animals were removed aseptically and washed twice in the sterile PBS. The splenocytes were extracted as described earlier 27 and cultured in RPMI-1640 medium containing 10% FCS and concanavalin-A as mitogen, at 37°C for 72 hours. Duplicate cultures were set from each animal for the each drug/radiation dose.
Micronucleus Assay
Micronuclei were prepared according to the method of Fenech and Morley 14 with minor modifications. Briefly, 40 hours after initiation of splenocyte culture, 5 µg/mL of cytochalasin-B was added to each culture and the cells were allowed to grow for the next 32 hours. The splenocytes were harvested at 72 hours after initiation of the cultures by centrifugation. The splenocytes were subjected to mild hypotonic treatment (0.7% ammonium oxalate) so as to retain the cytoplasm, fixed in Carnoy’s fixative (3:1 methanol: acetic acid), centrifuged again and resuspended in a small volume of fixative. The cell suspension was placed onto precleaned coded wet slides to avoid observer’s bias and the cells were spread by gentle blowing. Cells were stained with acridine orange (25 mg/50 mL; BDH, England, Gurr Cat. No. 34001 9704640E) in Sorensen’s buffer (pH 6.8). The slides were washed twice in Sorensen’s buffer, mounted and observed under a fluorescent microscope, equipped with 450 to 490 nm BP filter set with excitation at 453 nm (Photomocroscope III, Carl Zeiss, Oberkohn, Germany), using a 40× Neofluar objective. A minimum of 1000 binucleate splenocytes (BNCs) with well-preserved cytoplasm were scored from each culture and the frequency of micronucleated binucleate cells (MNBNCs) was determined. The micronuclei identification was done as described earlier. 16 A total of 48 animals were used for this experiment. The protection factor (PF) at each radiation exposure dose was calculated as follows 7 :

Free Radical Scavenging
The effect of SCE was studied on the scavenging of following free radicals in cell free system in vitro.
Hydroxyl radical
Scavenging of hydroxyl (•OH) free radical was measured by the method of Halliwell et al. 28 Briefly, the reaction mixture contained deoxyribose (2.8 mM), KH2PO4–NaOH buffer, pH 7.4 (0.05 M), FeCl3 (0.1 mM), EDTA (0.1 mM), H2O2 (1 mM), ascorbate (0.1 mM), and SCE (50-500 µg/mL) in a final volume of 2 mL. The reaction mixture was incubated for 30 minutes at ambient temperature, followed by the addition of 2 mL of TCA (2.8% w/v) and TBA. The reaction mixture was kept in a boiling water bath for another 30 minutes, cooled, and the absorbance was read at 532 nm in a UV–visible double beam spectrophotometer (UV-260; Shimadzu Corporation, Tokyo, Japan). All the analyses were done in triplicate and average values were taken. Inhibition (I) of deoxyribose degradation in percentage was calculated according to the equation

where A0 is the absorbance of the control reaction and A1 is the absorbance of the test compound.
Superoxide anion radical
Scavenging of superoxide (

where A0 is the absorbance of the control and A1 is the absorbance of the test compound.
DPPH Scavenging Activity
The ability of SCE to scavenge the stable free radical DPPH was measured as a decrease in absorbance at 517 nm. 30 The methanolic solution of DPPH (0.05 mM) was mixed with an equal volume of SCE (10-400 µg/mL), to make up the final volume up to 1.0 mL. An equal amount of methanol was added to the control. After 20 minutes, absorbance was recorded at 517 nm in a UV–visible double beam spectrophotometer. Lower absorbance of the reaction mixture indicates higher free radical scavenging activity.
Total Antioxidant Activity Assay
Total antioxidant potential was determined by the ABTS assay, as described earlier. 31 This technique measures the relative ability of antioxidant substances to scavenge the ABTS•+ cation radical generated in the aqueous phase. The reaction mixture contained ABTS (0.00017 M), SCE (10-100 µg/mL), and buffer in a total volume of 3.5 mL. The absorbance was measured at 734 nm in a UV–visible double beam spectrophotometer.
Preparation of Brain Homogenate
A separate experiment was undertaken to study the effect of SCE on FeCl3-induced lipid peroxidation in mouse brain homogenate where the animals were anesthetized with diethyl ether, decapitated, and perfused transcardially with ice cold normal saline to prevent contamination of brain tissue with blood. The brain was removed and weighed, homogenized (10% w/v) in 0.15 M potassium chloride, and centrifuged at 13 000 rpm for 10 minutes. The supernatant was immediately used for the estimation of in vitro lipid peroxidation. 32
Inhibition of Lipid Peroxidation by SCE
The reaction mixture contained 0.5 mL of brain homogenate, 100 µL of 150 mM potassium chloride, and ethanol (10 µL) or various concentrations of SCE, and 100 µM FeCl3 (to initiate lipid peroxidation) in a final volume of 1 mL incubation mixture. The reaction mixture without SCE was considered as control. The above mixture was incubated for 20 minutes at 37°C, and the reaction was stopped by adding 2 mL ice cold 0.25 N hydrochloric acid containing 15% trichloroacetic acid, 0.38% thiobarbituric acid, and 0.05% BHT. Finally, the mixture was heated at 80°C for 15 minutes, cooled, centrifuged at 1000g for 10 minutes, and the absorbance of the supernatant was measured at 532 nm. The amount of lipid peroxidation was determined using the molar extinction coefficient of 1.56 × 10 5 M− 1 cm− 1 and expressed as TBARS. 33 Percentage inhibition of TBARS formed was calculated by comparing control samples. All solutions were freshly prepared in distilled water immediately before use. Since most buffers trap hydroxyl radical or interfere with iron, 33 the reactions were unbuffered and carried out in 0.15 M KCl. Results are means of triplicate experiments.
Statistical Analysis
The statistical significance was determined by one-way analysis of variance with Bonferroni’s post hoc test used for multiple comparisons. Student’s t test was used for biochemical studies. Solo 4 (BMDP Statistical Software, Los Angeles, CA) was used for statistical analysis.
Results
The results are expressed as frequency of MNBNCs per 1000 ± SEM (standard error of the mean), and percentage inhibition of TBARS and various free radicals is presented in the figures and tables latter in the text.
Radioprotective Effect
The baseline MNBNC frequency in the CMC + sham-irradiated mice was 14.33 ± 1.45 and administration of 50 mg/kg SCE orally once daily for 5 consecutive days before sham-irradiation did not alter the baseline frequency of MNBNC (13.30 ± 1.73) significantly (Table 1). The frequency of MNBNC increased in a dose-dependent manner in CMC + irradiation group and the dose–response relationship was linear (Figure 1). Pretreatment of animals with SCE before exposure to different doses of γ-radiation resulted in a significant decline in the frequency of MNBNC when compared with CMC + irradiation group and the greatest protection factor was obtained for 0.5 Gy (2.6), which declined with increase in radiation dose and the lowest protection factor of 1.6 was observed for 4 Gy (Table 2). The dose effect relationship was linear (Table 1).

Effect of SCE on the induction of MNBNC (total) in cultured splenocytes exposed to different doses of γ-irradiation: ■ CMC + irradiation; • SCE + irradiation
Alteration of Radiation-Induced MN Formation by 50 mg/kg SCE in Cultured Splenocytes Exposed to Various Doses of γ-Radiation

Abbreviations: MN, micronuclei; SCE, Syzygium cumini extract; BNC, binucleated cells; SEM, standard error of the mean; CMC, carboxymethyl cellulose; IR, irradiation.
P < .05, b P < .01, c p < .001 versus control.
P < .05, y P < .01, z P < .001 versus 0.5 Gy.
P < .05, ♠ P < .01, # P < .001 versus 1 Gy.
P <.05, π P < .01, ϒ P < .001 versus 2Gy.
P < .01, Ω P < .001 versus 3 Gy.
P < .001 versus 4 Gy.
Protection Factor of SCE
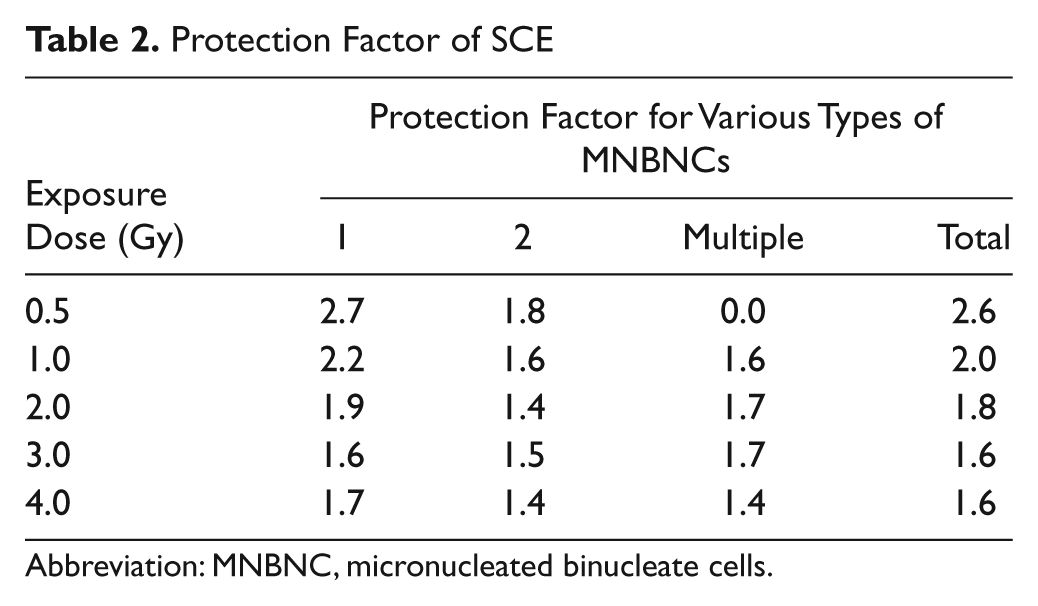
Abbreviation: MNBNC, micronucleated binucleate cells.
The frequencies of BNC with 1, 2, and multiple MN are represented separately in Figures 2, 3, and 4, respectively. The frequency of BNC with 1 MN was elevated in a dose-dependent manner in the CMC + irradiation group and a maximum number of MNBNC were scored for 4 Gy, the last irradiation dose studied (Figure 2). Administration of animals with 50 mg/kg body weight of SCE before exposure to different doses of γ-radiation resulted in a significant inhibition of MNBNC bearing 1 MN when compared with the CMC + irradiation group. The protection factor for BNC with 1 MN was 2.7 for 0.5 Gy, which declined with increasing dose of radiation (Table 2). The curve fitting of data revealed a linear dose–response relationship for both groups (Figure 1 and Table 1).

Effect of SCE on the induction of BNC with 1 MN in cultured splenocytes exposed to different doses of γ-irradiation: ■ CMC + irradiation; •SCE + irradiation

Effect of SCE on the induction of BNC with 2 MN in cultured splenocytes exposed to different doses of γ-irradiation: ■ CMC + irradiation; • SCE + irradiation

Effect of SCE on the induction of BNC bearing multiple MN in cultured splenocytes exposed to different doses of γ-irradiation: ■ CMC + irradiation; • SCE + irradiation
The frequency of BNC with 2 MN showed a dose-dependent rise in the CMC + irradiation group (Figure 3). Oral administration of SCE before irradiation resulted in a significant reduction in the frequency of BNC with two MN when compared with the CMC + irradiation group (Table 1). The highest frequency of MNBNC with 2 MN was observed at 4 Gy irradiation. A highest protection factor of 1.8 was obtained for 0.5 Gy, which declined steadily with increasing dose of radiation (Table 2). Both CMC and SCE + irradiation group showed a linear dose–response relationship (Table 1).
The BNCs with multiple (≥3) MN were conspicuous by their absence for CMC + sham-irradiation (Figure 4) and with further increase in the irradiation dose, the frequency of BNC with multiple MN increased in a dose-dependent fashion, and the maximum number of multiple MNBNCs was observed for 4 Gy. Oral treatment of animals with SCE before irradiation resulted in a significant decline in the frequency of multiple MNBNCs at all the exposure doses and protection factor of approximately 1.6 was observed for 1 to 3 Gy, except 4 Gy where it was 1.4 (Table 2). The dose response was linear for CMC + irradiation and SCE + irradiation groups.
Hydroxyl Radical Scavenging
This assay shows the ability of SCE to inhibit hydroxyl radical–mediated deoxyribose degradation in a reaction mixture with EDTA. The relative extents of inhibition of deoxyribose degradation will give an indication of •OH scavenging activity. SCE inhibited the generation of hydroxyl radical in a dose-dependent manner, and a maximum inhibition of OH free radicals was observed at 350 µg/mL (51%); thereafter a steady state was attained (Figure 5).

Effect of various concentrations of Syzygium cumini extract (SCE) on the scavenging of hydroxyl free radicals
Superoxide Anion Radical Scavenging
Superoxide anions are a precursor to active free radicals that have the potential of reacting with biological macromolecules and thereby inducing tissue damage. SCE inhibited the generation of
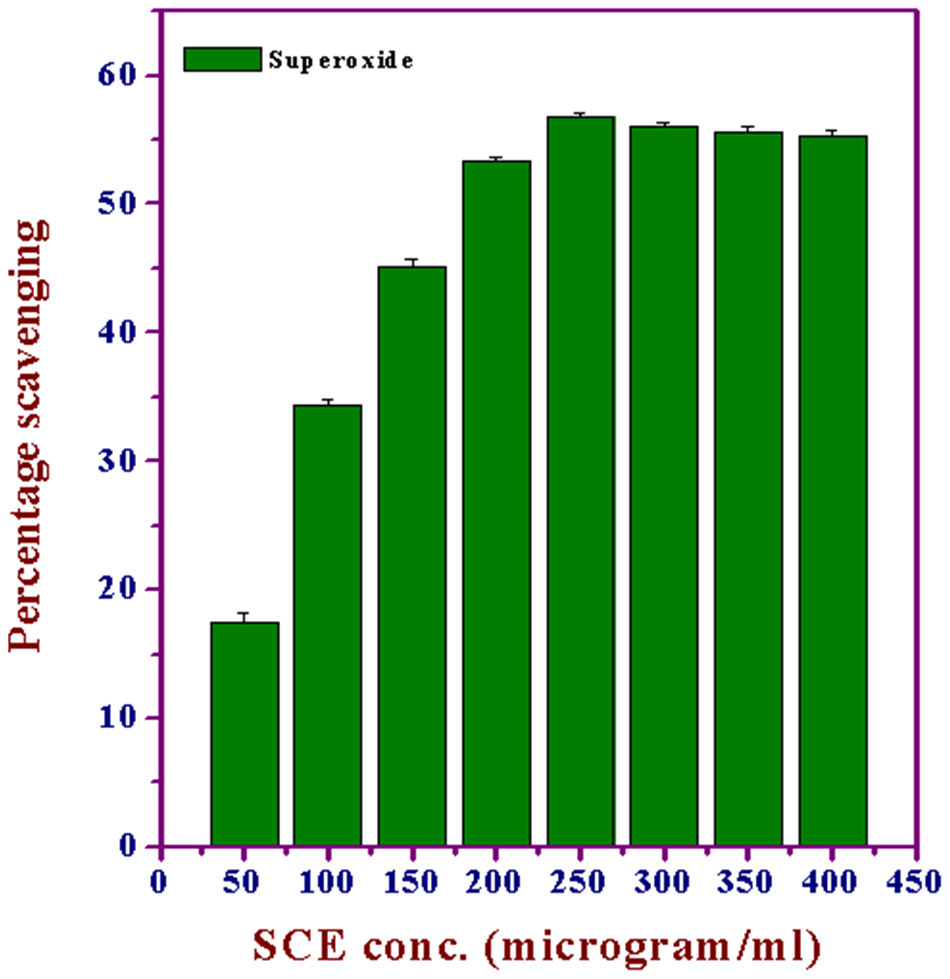
Effect of various concentrations of Syzygium cumini extract (SCE) on the scavenging of superoxide anion free radicals
DPPH Free Radical Scavenging
DPPH has been widely used to evaluate the free radical scavenging effects of various antioxidant substances. 34 In the DPPH assay, the antioxidants were able to reduce the stable radical DPPH to the yellow colored diphenyl picrylhydrazine. The method is based on the reduction of alcoholic DPPH solution in the presence of a hydrogen-donating antioxidant because of the formation of the nonradical form DPPH-H by the reaction. With this method it is possible to determine the antiradical power of an antioxidant by measuring of a decrease in the absorbance of DPPH at 517 nm. Because of a resulting color change from purple to yellow, the absorbance decreased when the DPPH• was scavenged by an antioxidant through donation of hydrogen to form a stable DPPH molecule. In the radical form, this molecule had an absorbance at 517 nm, which disappears after acceptance of an electron or hydrogen radical from an antioxidant compound to become a stable diamagnetic molecule. 35 Figure 7 illustrates a decrease in the concentration of DPPH radical because of the scavenging ability of SCE in a concentration-dependent manner, and a maximum inhibition was observed at 80 µg/mL (72.6%); thereafter, a steady state was attained.
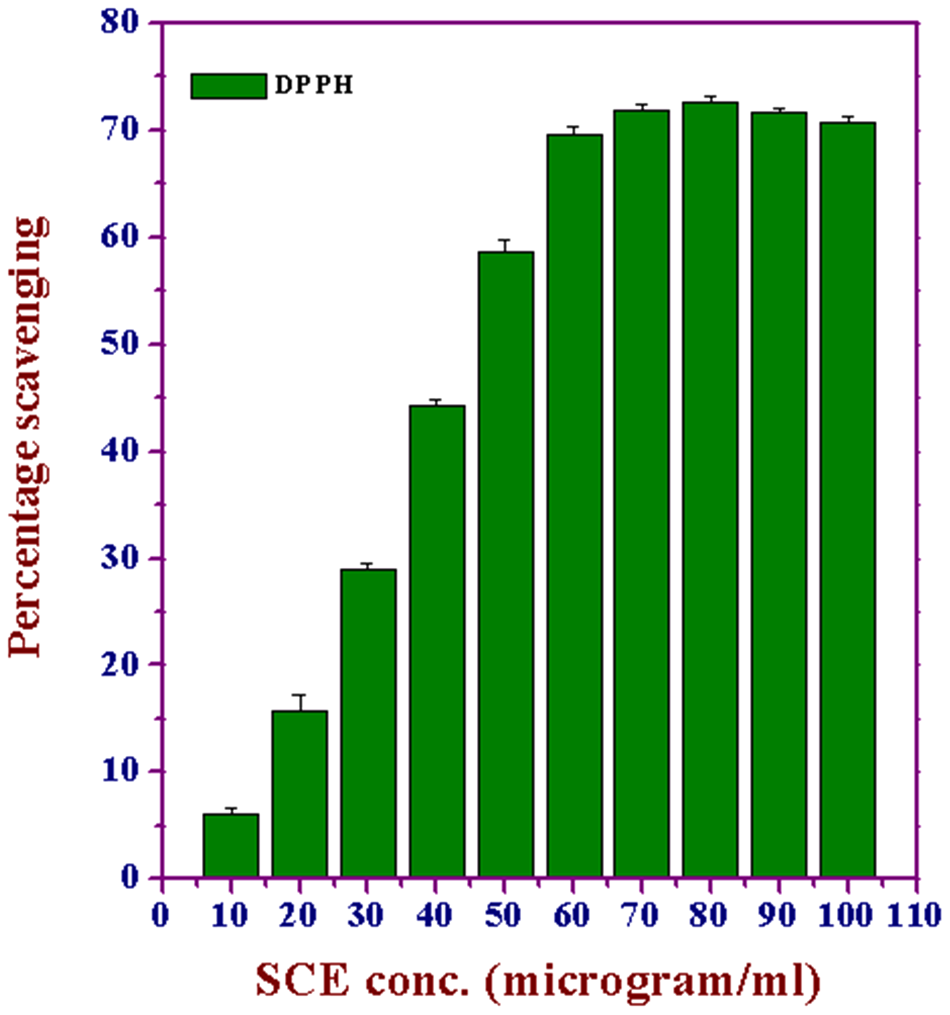
Effect of various concentrations of Syzygium cumini extract (SCE) on the scavenging of 2,2-diphenyl-1-picrylhydrazyl (DPPH) free radicals
ABTS Radical Scavenging
Generation of the ABTS radical cation forms the basis of one of the spectrophotometric methods that have been applied for the measurement of the total antioxidant activity of solutions of pure substances, aqueous mixtures, and beverages. 31 A more appropriate format for the assay is a decolorization technique where the radicals are generated directly in a stable form prior to reaction with putative antioxidants. The improved technique for the generation of ABTS•+ described here involves the direct production of the blue/green ABTS•+ chromophore through the reaction between ABTS and potassium persulfate. SCE inhibited the generation ABTS•+ radical in a concentration-dependent manner (10-100 µg/mL), and a maximum inhibition was observed at 80 µg/mL (72.6%), which declined steadily thereafter (Figure 8).

Effect of various concentrations of Syzygium cumini extract (SCE) on the scavenging of 2,2-azinobis(3-ethylbenzothiazoline-6-sulfonic acid) diammonium salt (ABTS+) cation free radicals
Inhibition of Lipid Peroxidation by SCE
Lipid peroxidation is an important pathophysiological event in a variety of diseases, drug toxicities, and traumatic or ischemic injuries. In the central nervous system, for example, the role of oxygen free radicals and lipid peroxidation results in the irreversible loss of neuronal tissue following brain injury 36 and degenerative neurological disorders. 37 SCE inhibited the formation of TBARS in a dose-dependent manner (50-500 µg/mL) in brain homogenates, and the maximum inhibition was observed at 350 µg/mL (65.34%), which declined steadily thereafter (Figure 9).
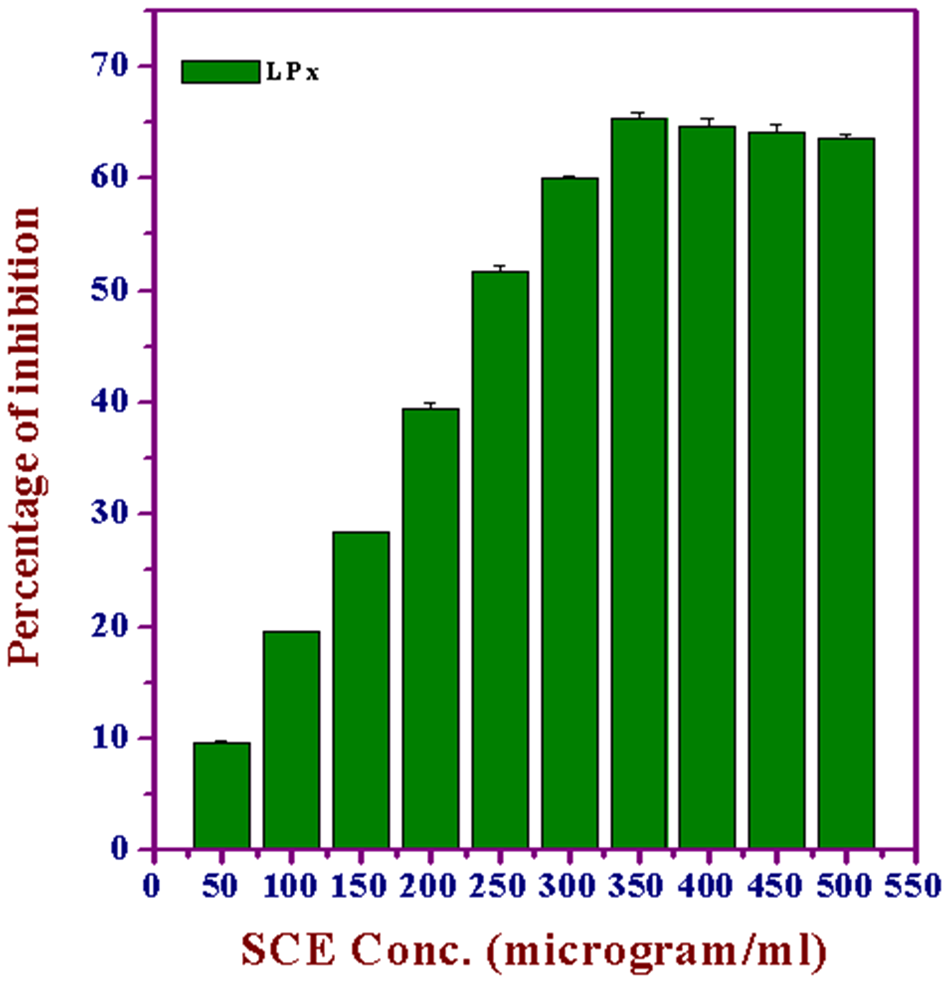
Effect of various concentrations of Syzygium cumini extract (SCE) on lipid peroxidation
Discussion
The DNA damages, especially the double-strand breaks produced by ionizing radiation, are converted into chromosome aberrations, which are subsequently expressed as micronuclei after one cell division.16,38,39 Micronuclei can be assessed in dividing cells easily and are a measure of DNA damage. 40 The micronucleus assay has been used to screen genotoxic agents and can serve as a short-term test to screening radioprotective ability of any pharmacological agent.1,41-43 Therefore, present study was undertaken to investigate the radioprotective potential of SCE in cultured splenocytes of mice exposed to different doses of γ-radiation.
The acentric chromosome or chromatid fragments or whole chromosomes or chromatids that lag behind in anaphase of cell division because of defective kinetochore or spindle fiber/s are not included in the daughter nuclei during telophase and are converted into small rounded or spherical extranuclear bodies that can be observed in the cytoplasm of daughter cell/s and are known as micronuclei. 9 The assessment of micronuclei gives an indirect indication of DNA damage as micronuclei harboring chromosomal fragments may arise from direct double-strand DNA breakage, conversion of single-strand breaks to double-strand breaks after cell replication, or inhibition of DNA synthesis. Exposure of animals to different doses of γ-radiation resulted in a dose-dependent elevation in MNBNCs and the dose–response relationship was linear. A similar effect has been observed earlier.5,44-47 However, a linear quadratic response has also been reported.23,48-50 Irradiation not only increased the frequency of BNC with 1 MN but also with 2 or multiple MN, indicating that induction of complex multiple sites of DNA damage and production of these lesions was radiation dose dependent. Irradiation has been reported to increase the frequency of cells bearing 1, 2, or multiple MN in lymphocytes and various cultured cell lines.5,23,43,46,51-53
Treatment of animals with 50 mg/kg body weight of SCE significantly reduced the frequency of MNBNCs when compared with CMC + irradiation group. An earlier study from this laboratory has reported that leaf extract of S. cumini reduced the MNBNC frequency in HPBLs; however, no attempt was made to study the effect of SCE after different doses of radiation. 23 Likewise, plant polyphenols such as naringin and mangiferin have been reported to protect mice and HPBLs against radiation-induced MN formation.43,47 Certain other plant flavonoids, such as luteolin, quercetin, kaempferol, morin, rutin, fisetin, myricetin, epicatechin, genistein, eridictyol, phloretin, naringin, and hesperidin have been found to protect mice bone marrow cells against the radiation-induced micronuclei.47,54,55 SCE not only protected against the formation of BNC with 1 MN but also with 2 or multiple MNs in the cultured splenocytes after exposure to different doses of γ-radiation, indicating that SCE could efficiently protect complex multiple sites of DNA damage. An identical effect has been observed earlier with Agele marmelos extract and mangiferin in HPBLs.5,43 Likewise, a reduction in polychromatic erythorcytes bearing 2 or more MN has been observed in mice bone marrow treated with (E)4-(4-N, N-dimethyl aminophenyl) but-3-en-2-one, 44 naringin, a citrus bioflavonoid, 47 the extracts of abana, 45 , S. cumini, 24 and A. marmelos, 5 and also mangiferin. 43
The whole berry extract of Hippophae rhamnoides, labeled RH-3, has been found to render antioxidant-mediated radioprotection, by preventing cellular and mitochondrial free radical generation. 56 Leaf extract of Aloe vera has also been demonstrated to have damage-resistant properties against radiation-induced biochemical alterations in Swiss albino mice. 57 The radioprotective property of Podophyllum hexandrum fraction (REC-2006) in p53-carrying hepatoma cell line, perhaps mediated by cell cycle regulatory proteins 58 and free radical scavenging activity and radioprotection of Caesalpinia digyna extracts 59 has been reported. More recently, the radioprotective property of a seed extract of Silymarin, with INM-7035 as its bioactive constituent 60 and the radiation mitigating effect of hydro-alcoholic extract of Saraca indica (SIE) against mice exposed to whole body γ-radiation have also been reported. 61
The exact mechanism by which SCE protects against the genotoxic effects of radiation is not known. There may be several putative mechanisms by which SCE may have protected mouse splenocytes against radiation-induced micronuclei formation. Ionizing radiation is a potent inducer of free radicals and oxidative stress. Superoxide anion, arising either through metabolic processes or following oxygen “activation” by physical irradiation, is considered the “primary” reactive oxygen species, whereas the hydroxyl radical, •OH, has a high reactivity, making it a very dangerous radical with a very short in vivo half life.
62
These free radicals elicit a cascade of events that eventually interact with molecular DNA.
63
Hence scavenging of these radiation-induced free radicals by SCE may be one of the important mechanisms to reduce radiation-induced DNA damage. This contention is supported by the in vitro results in this study, where SCE scavenged •OH,
The present study demonstrates that oral administration of jamun extract for 5 consecutive days before exposure to various doses of γ-radiation reduced the radiation-induced micronuclei formation and also protected splenocytes against the complex multiple sites of DNA damage. The radioprotective activity of jamun may not be because of single mechanism but may be because of interplay of several mechanisms. It may protect the cells against radiation damage by scavenging of free radicals and blocking radiation-induced lipid peroxidation. It may also upregulate antioxidant mechanisms of cells, thereby reducing DNA damage. Jamun may have also inhibited the activation of NF-κB and COX-II mRNA. It may have also upregulated DNA polymerase and efficiently repaired the lesions induced by radiation in the cellular genome.
It has recently been suggested that data from Asian botanicals (from both Chinese and Ayurvedic medicine) are promising enough to be considered for clinical trials. 76 Additionally, if a plant preparation shows in vitro and in vivo efficacy against radiation, then in-depth exploration should be initiated instead of looking for further screening of more flora. In this context, the need of high throughput assays in understanding the complex signaling pathways involved in radioprotection cannot be underestimated. 77 Most important, it is also vital to evaluate them for efficacy, tumor specificity, and safety profiles. 78 Furthermore, knowledge about the mechanism of action, chemical nature, shelf life, ease of administration, absorption, and distribution characteristics are some of the prime requisites before clinical trials can be taken up in this regard. 77 To conclude, we are optimistic and support the view that a definite and systematic path toward the development of herbal radioprotector will certainly deliver a highly potential and safe drug for human use.
Footnotes
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
The authors received no financial support for the research, authorship, and/or publication of this article.