
Abstract
Isolinderanolide B (IOB), a butanolide extracted from the stems of Cinnamomum subavenium, was investigated for its antiproliferative activity in T24 human bladder cancer cells. To identity the anticancer mechanism of IOB, its effect on apoptosis, cell cycle distribution, and levels of p53, p21 Waf1/Cip1, Fas/APO-1 receptor, and Fas ligand was assayed. Enzyme-linked immunosorbent assay showed that the G0/G1 phase arrest is because of increase in the expression of p21 Waf1/Cip1. An enhancement in Fas/APO-1 and membrane-bound Fas ligand (mFasL) might be responsible for the apoptotic effect induced by IOB. This study reports the novel finding that the induction of p21 Waf1/Cip1 and activity of the Fas/mFas ligand apoptotic system may participate in the antiproliferative activity of IOB in T24 cells.
Introduction
Bladder cancer is the fourth most common cancer in humans, and survival of high-risk bladder cancer remains problematic when first-line curative measures have failed. 1 Although most bladder cancer cases are superficial at the time of diagnosis, repeated transurethral resections and intravesical chemo/immunotherapy may be required to treat recurrences. Each of these interventions, particularly intravesical therapy, is accompanied by side effects.2,3 There is an obvious urgent need for novel and effective therapies against bladder cancer.
Apoptosis plays an important role in homeostasis and development of tissue in the organism. 4 Imbalance between cell proliferation and apoptotic cell death will result in serious disease such as cancer. Many studies have demonstrated that cancer treatment by chemotherapy and γ-irradiation kills target cells primarily by the induction of apoptosis.4,5 Several previous publications have reported that anticancer agents may induce apoptosis via the Fas/FasL system.6-11 Fas is a cell surface receptor comprising a type I integral membrane protein that expresses a cytoplasmic death domain and belongs to the tumor necrosis factor receptor superfamily. 12 Activation of Fas by its ligand (FasL) results in the oligomerization of its intracellular death domain and the recruitment of the intracellular adaptator FADD (Fas-associated death domain). Once bound, FADD is able to activate procaspase-8 and procaspase-10 in a death inducing signaling complex. In turn, caspase-8 and caspase-10 activate downstream caspases, resulting in apoptotic cell death. 13
Cinnamomum subavenium Miq. (Lauraceae) is a medium-sized evergreen tree, found in central to southern mainland China, Burma, Cambodia, Taiwan, Malaysia, and Indonesia.14,15 Isolinderanolide B (IOB; Figure 1) is a butanolide constituent isolated from the stems of C subavenium, and its properties as an antitumor agent have not yet been fully described.14,16 This study is the first to determine the cell growth inhibition activity of IOB and examine its effect on cell cycle distribution and apoptosis in T24 human bladder cancer cells.
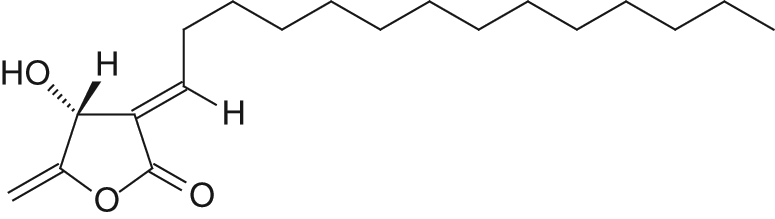
Chemical structure of isolinderanolide B from Cinnamomum subavenium.
Materials and Methods
Chemicals and Reagents
Fetal bovine serum (FBS), penicillin G, streptomycin, amphotericin B, and Dulbecco’s modified Eagle’s medium (DMEM) were obtained from GIBCO BRL (Gaithersburg, MD). Dimethyl sulfoxide (DMSO), ribonuclease (RNase), and propidium iodide (PI) were purchased from Sigma Chemical (St Louis, MO). XTT enzyme-linked immunosorbent assay (ELISA) kit was obtained from Roche Diagnostics GmbH (Mannheim, Germany). p53 ELISA, Fas Ligand ELISA, and caspase-8 assay kits, and caspase-8 inhibitor, benzyloxy-carbonyl-Val-Ala-Asp-fluoromethylketone (Z-IETD-FMK) were purchased from Calbiochem (Cambridge, MA). p21 Waf1/Cip 1 and Fas/APO-1 ELISA kits were purchased from Invitrogen (Camarillo, CA). Anti-Fas Ab (ZB4) was obtained from Upstate Biotechnology, Inc (Lake Placid, NY).
Test Compound
Isolinderanolide B was isolated from the stems of C subavenium as described previously. 14 Briefly, the air-dried stems were extracted with methanol (MeOH) at room temperature and the MeOH extract was obtained on concentration under reduced pressure. The MeOH extract, suspended in H2O, was partitioned with CHCl3 to give fractions soluble in CHCl3 and H2O. The CHCl3-soluble fraction was chromatographed over silica gel using n-hexane/EtOAc/MeOH mixtures as eluents to produce 5 fractions. Part of fraction 1 was subjected to silica gel chromatography by eluting with n-hexane–EtOAc (30:1), enriched with EtOAc to furnish 10 fractions (1-1 to 1-10). Fraction 1-3 was subjected to silica gel chromatography, eluting with n-hexane–EtOAc (40:1) and enriched gradually with EtOAc, to obtain 3 fractions (1-3-1 to 1-3-3). Fraction 1-3-2, eluted with n-hexane–EtOAc (40:1), was further separated using silica gel column chromatography and preparative thin layer chromatography (n-hexane–EtOAc, 30:1) and gave isolinderanolide B.
Cell Culture
Bladder cancer cell line T24 (ATCC HTB-4) was obtained from the American Type Cell Culture Collection (ATCC, Rockville, MD). It was maintained in monolayer culture at 37°C and 5% CO2 in DMEM supplemented with 10% FBS, 100 units/mL of penicillin G, 100 µg/mL of streptomycin, and 0.25 µg/mL of amphotericin B.
Cell Proliferation Assay
Inhibition of cell proliferation by IOB was measured by XTT {sodium 3′-[1-(phenylamino-carbonyl)-3,4-tetrazolium]-bis(4-methoxy-6-nitro) benzene-sulfonic acid hydrate} assay. Briefly, cells were plated in 96-well culture plates (1 × 104 cells/well). After 24-hour incubation, the cells were treated with vehicle alone (0.1% DMSO) and IOB (1.5, 3, 6, and 12 µM) for 24 and 48 hours. A 50-µL XTT test solution, which was prepared by mixing 5 mL of XTT-labeling reagent with 100 µL of electron coupling reagent, was then added to each well. After 6-hour incubation, the absorbance was measured on an ELISA reader (Multiskan EX, Labsystems, Thermo Electron Corporation, Milford, MA) at a test wavelength of 492 nm and a reference wavelength of 690 nm.
Cell Cycle Analysis
To determine cell cycle distribution, 5 × 105 cells were plated in 60-mm dishes and treated with vehicle alone (0.1% DMSO) and IOB (3 and 6 µM) for 24 hours. After treatment, the cells were collected by trypsinization, fixed in 70% ethanol, washed in phosphate-buffered saline (PBS), resuspended in 1 mL of PBS containing 1 mg/mL RNase and 50 µg/mL propidium iodide, incubated in the dark for 30 minutes at room temperature, and analyzed by EPICS flow cytometer. The data were analyzed using the Multicycle software (Phoenix Flow Systems, San Diego, CA).
The Analysis of Apoptosis
Apoptotic cells were quantitatively determined by the terminal deoxynucleotidyl transferase-mediated deoxyuridine triphosphate nick endlabeling (TUNEL) method, which examines DNA-strand breaks during apoptosis by using BD ApoAlert DNA Fragmentation Assay kit. Briefly, cells were incubated with vehicle alone (0.1% DMSO) and IOB (3 and 6 µM) for the indicated times. The cells were trypsinized, fixed with 4% paraformaldehyde, and permeabilized with 0.1% Triton X-100 in 0.1% sodium citrate. After being washed, the cells were incubated with the reaction mixture for 60 minutes at 37°C. The stained cells were then analyzed with an EPICS flow cytometer and a fluorescence microscope at 20× magnification.
Assaying the Levels of p53, p21, Fas/APO-1, and Fas Ligand (mFasL and sFasL)
p53 ELISA, p21 Waf1/Cip1, Fas/APO-1 ELISA, and Fas Ligand ELISA kits were used to detect p53, p21 Waf1/Cip1, Fas/APO-1 receptor, and soluble (sFasL)/membrane-bound Fas ligand (mFasL). Briefly, T24 cells were treated with vehicle alone (0.1% DMSO) and IOB (3 and 6 µM) for 6, 12, 24, and 48 hours. The samples of cell lysate were placed in 96-well microtiter plates that were coated with monoclonal detective antibodies, and incubated at room temperature. Each sample was assessed in triplicate. It was necessary to determine the soluble Fas ligand in cell culture supernatant by using Fas Ligand ELISA kit. After removing the unbound material by washing with washing buffer (50 mM Tris, 200 mM NaCl, and 0.2% Tween 20), the detector antibody that is bound by horseradish peroxidase, conjugated streptavidin, was added to bind to the antibodies. Horseradish peroxidase catalyzed the conversion of a chromogenic substrate (tetramethylbenzidine) to a colored solution with color intensity proportional to the amount of protein present in the sample. The absorbance of each well was measured at 450 nm, and concentrations of p53, p21 Waf1/Cip1, Fas/APO-1, and FasL were determined by interpolating from standard curves obtained with known concentrations of standard proteins.
Assay for Caspase-8 Activity
The assay is based on the ability of the active enzyme to cleave the chromophore from the enzyme substrate, Ac-IETD-pNA. The cell lysates were incubated with peptide substrate in assay buffer (100 mM NaCl, 50 mM HEPES, 10 mM dithiothreitol, 1mM EDTA, 10% glycerol, 0.1% CHAPS, pH 7.4) for 3 hours at 37°C. Each sample was assayed in triplicate. The release of p-nitroaniline was monitored at 405 nm. Results are represented as the percentage change of the activity compared with the untreated control.
Statistical Analysis
Data were expressed as means ± standard deviation (mean ± SD). Statistical comparisons of the results were made using analysis of variance. Significant differences (P < .05) between the means of control and IOB-treated cells were analyzed by Dunnett’s test.
Results
Effect of IOB on T24 Cell Proliferation
We first tested the antiproliferative effect of IOB in T24 human bladder cancer cells. As shown in Figure 2, the proliferative inhibitory effect of IOB was observed in a dose-dependent manner. At 48 hours, the maximal effect on proliferation inhibition was observed with 12 µM IOB, which inhibited proliferation in 90.7% of T24 cells. The IC50 value was 3.5 µM.

The antiproliferative effect of isolinderanolide B (IOB) in T24 cells
IOB-Induced Cell Cycle Arrest and Apoptosis in T24 Cells
The results on the effects of IOB on cell cycle progression of T24 are shown in Figure 3. Compared with the control, 3 µM IOB increased the population of G0/G1 phase from 39.7% to 53.6% in T24 cells. This effect was enhanced when T24 cells were treated by 6 µM IOB (67.3% cell population in G0/G1 phase of T24).
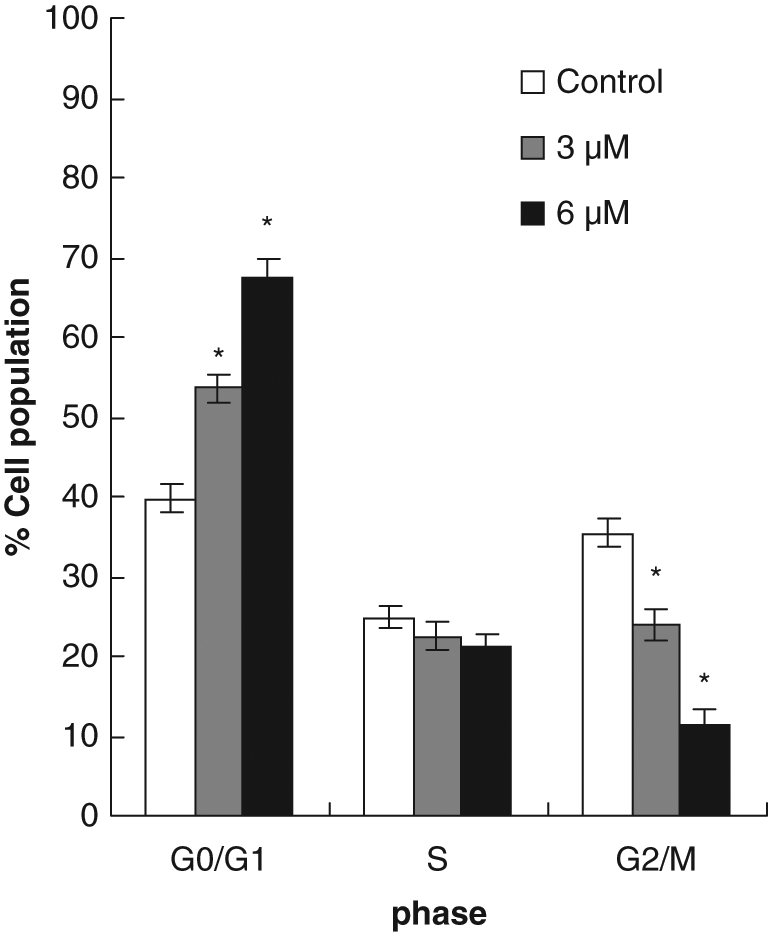
Effects of isolinderanolide B (IOB) on cell cycle distribution in T24 cells
Next, we assessed the effect of IOB on the induction of apoptosis in T24 cells. A quantitative evaluation was made using TUNEL to detect DNA-strand breaks. Compared to vehicle-treated cells, IOB induced 34.23% of apoptotic cells in T24 cells at concentrations of 6 µM at 48 hours (Figure 4).

Induction of apoptosis in T24 cells by isolinderanolide B (IOB)
IOB-Mediated Cell Cycle Arrest Operates Through the Induction of p21 Waf1/Cip1 Protein in T24 Cells
To determine whether tumor suppression factor p53 and its downstream molecule p21 Waf1/Cip1 are involved in the IOB-mediated antiproliferative effects of T24 cells, the levels of these proteins were assayed by ELISA kits. T24 cells were treated with 3 and 6 µM IOB for 6, 12, 24, and 48 hours. Treatment of IOB for up to 6 µM at 48 hours did not affect the protein expression of p53 (Figure 5A). The p21 Waf1/Cip1 protein is first characterized as a downstream target of p53 and is thought to be responsible for G0/G1 cell cycle arrest. 17 Surprisingly, Figure 5B shows that an increase in p21 Waf1/Cip1 protein was apparent at 6 hours and was observed in a time-dependent manner. Moreover, the induction of p21 Waf1/Cip1 was observed to be in a dose-dependent manner. Our results indicated that treatment of T24 cells with IOB was not observed to exert any changes on the expression of p53. Furthermore, IOB-mediated cell cycle arrest operates through the induction of p21 Waf1/Cip1 protein in T24 cells.

Effects of isolinderanolide B (IOB) on protein expression of p53 and p21 Waf1/Cip1
Fas/mFasL Apoptotic System Might Be a Possible Pathway of IOB-Mediated Apoptosis
One study demonstrated immunohistochemistry Fas and Fas-ligand expression in tumor tissue of patients with bladder cancer. 18 By using Fas/APO-1 ELISA and Fas Ligand ELISA kits, we found that IOB increased expression of Fas/APO-1 receptor and membrane-bound Fas ligand in T24 cells as early as 6 hours after treatment in a dose- and time-dependent manner (Figures 6A and 6B). We detected very low concentrations of the soluble-bound Fas ligand in culture supernatants of T24 cells (Figure 6C). This result was similar to a previous report. 19 The time relationship between the expression of Fas/mFasL at 6 hours of treatment and the occurrence of DNA fragmentation at 12 hours of treatment could support the idea that the Fas/mFasL system might mediate IOB-induced apoptosis of T24 cells.
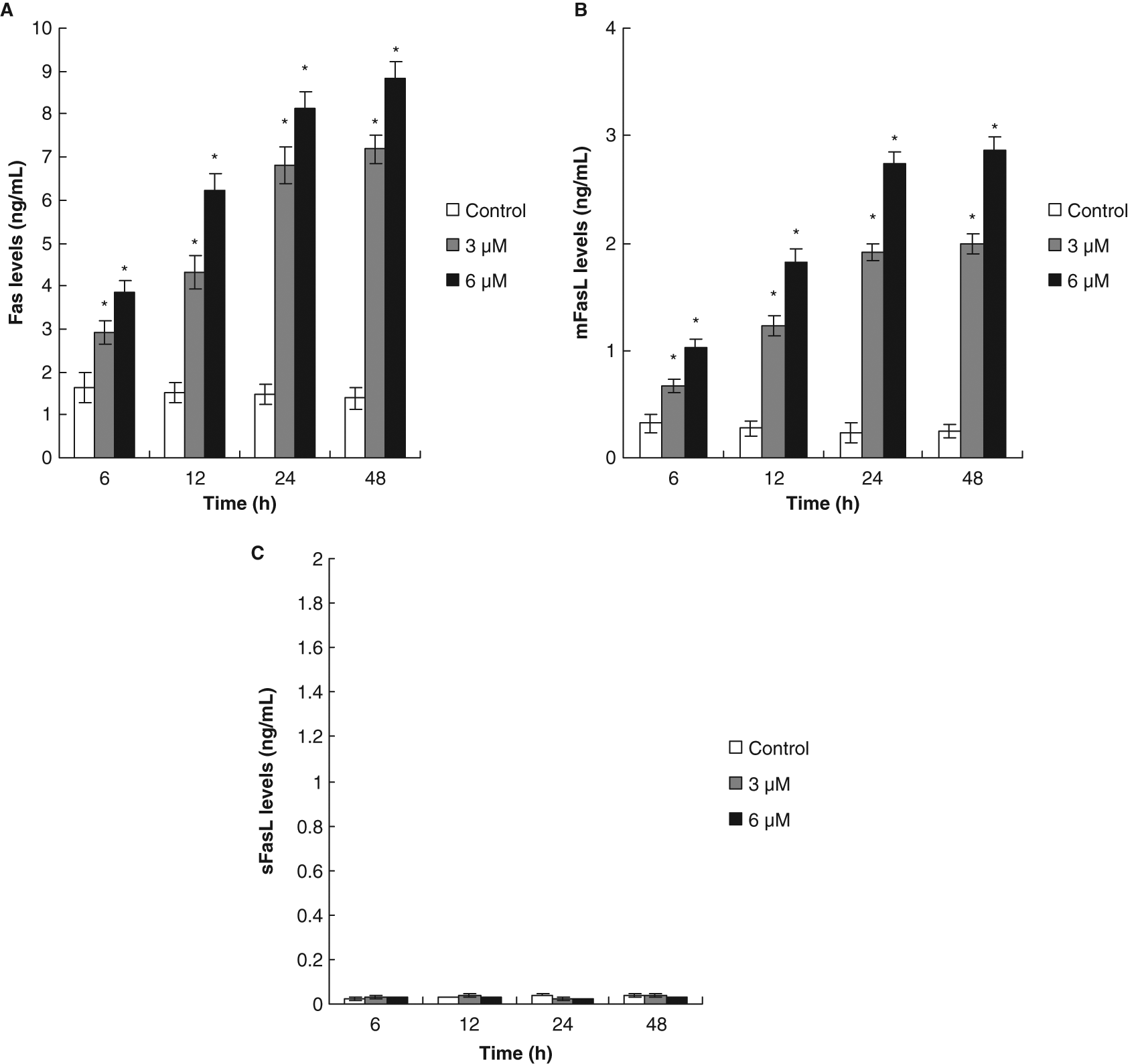
Fas/FasL apoptotic system was involved in isolinderanolide B (IOB)-mediated apoptosis
When T24 cells were pretreated with an antagonistic anti-Fas antibody, ZB4, the antiproliferative and proapoptotic effects of IOB were effectively inhibited. At 6 µM IOB, cell proliferation inhibition decreased from 71.2% to 16.5% in T24 cells (Figure 7A). Compared with the control, the induction of apoptosis induced by 6 µM IOB decreased from about 34.23% to 11.49% at 48 hours in ZB4-pretreated T24 cells (Figure 7B).
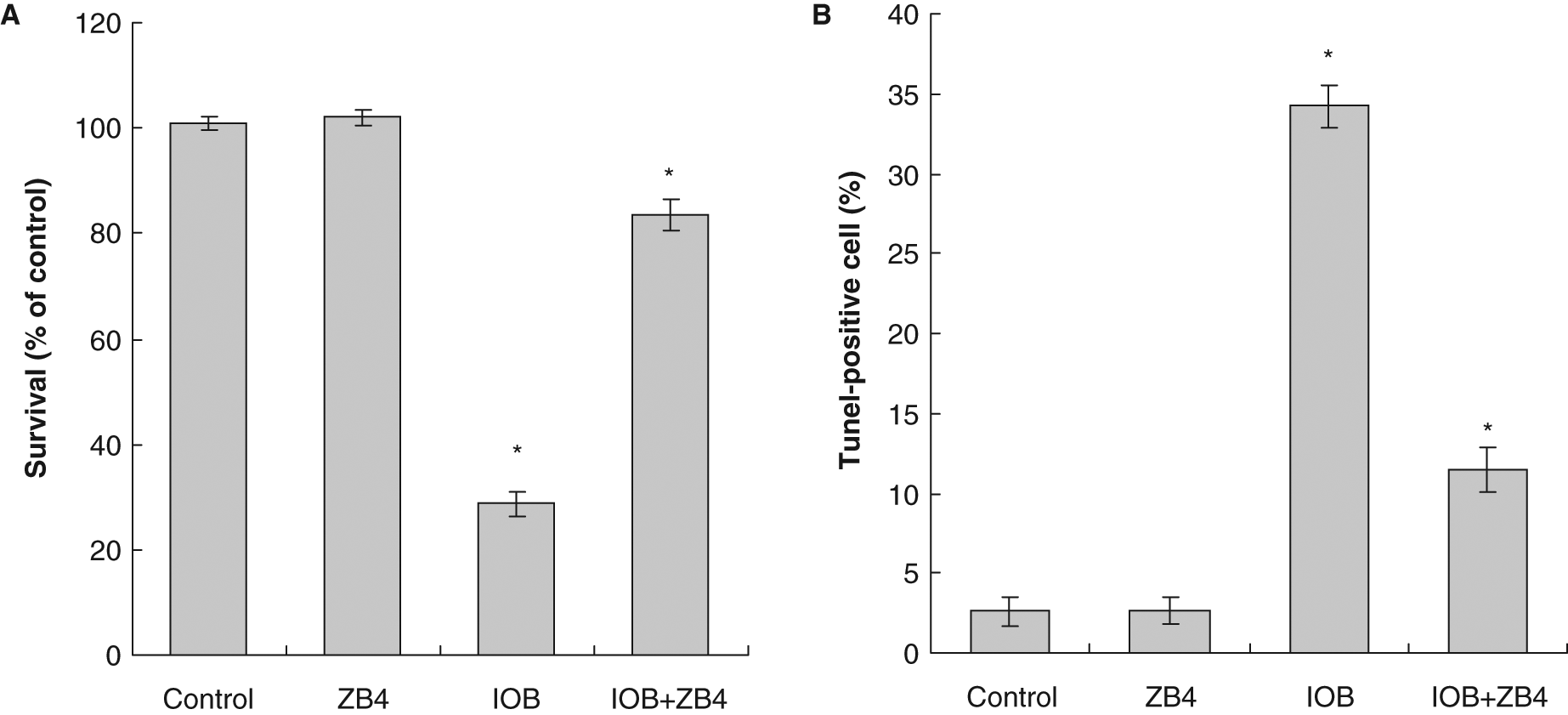
Effect of antagonistic anti-Fas antibody (ZB4) on isolinderanolide B (IOB) in T24 cells
Next, we measured the downstream caspase of Fas/FasL system. The results showed that caspase-8 activity increased at 10 hours and reached maximum induction at 48 hours in 6 µM IOB-treated T24 cells (Figure 8A). The activation of caspase-8 (at 10 hours) was before the production of DNA fragmentation (at 12 hours) showing caspase-8 activation was required in IOB-induced apoptosis. To further probe this hypothesis, we assessed the effect of caspase-8 inhibitor (Z-IETD-FMK) on the IOB-mediated antiproliferation and apoptosis. Our results showed that inhibition of caspase-8 not only decreased the IOB’s antiproliferative activity but also completely abolished induction of apoptosis in T24 cells (Figures 8B and 8C).
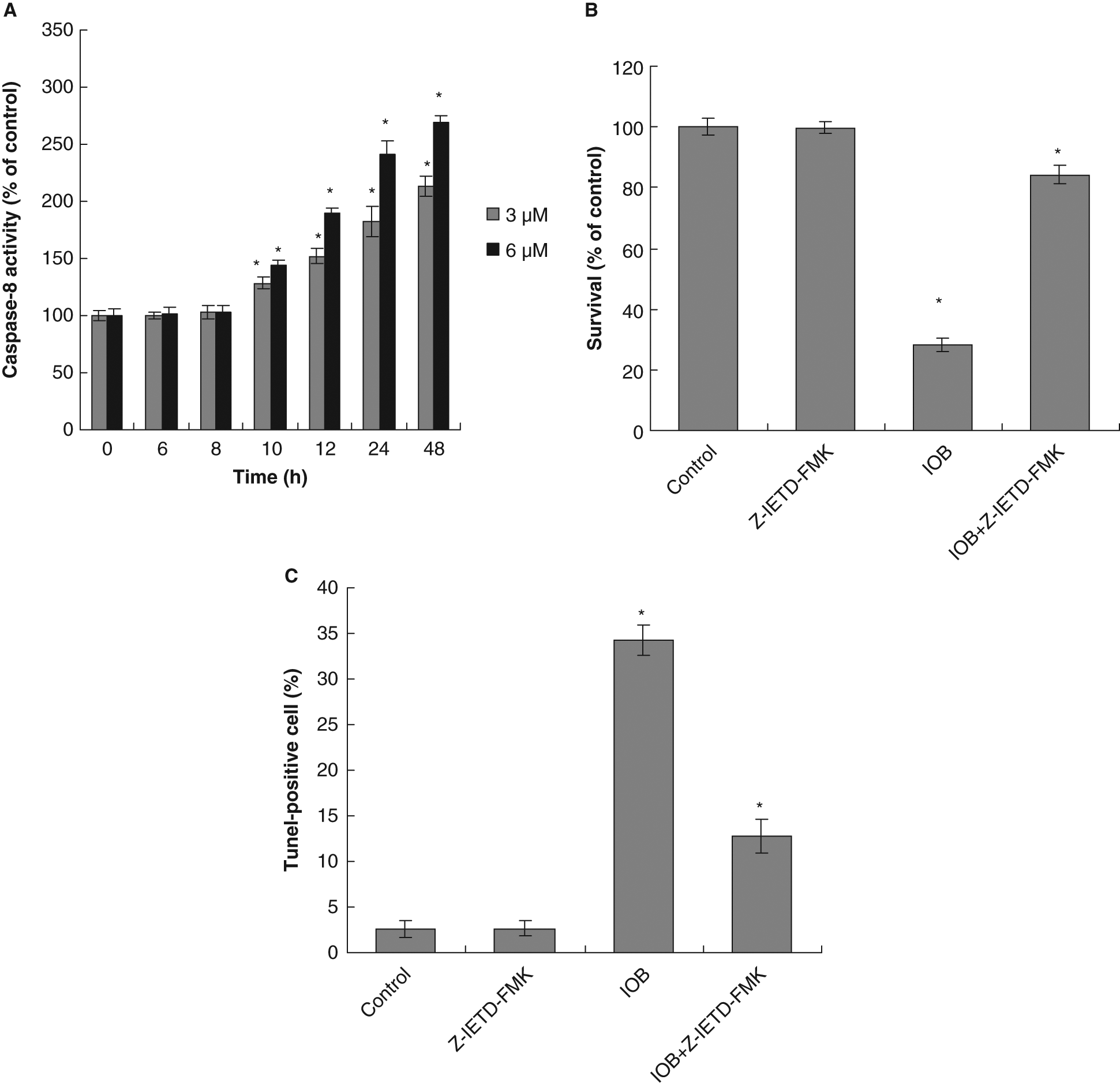
(A) The activation of caspase-8 in T24 cells by isolinderanolide B (IOB); (B) effect of caspase-8 inhibitor on IOB-mediated antiproliferation; and (C) effect of caspase-8 inhibitor on IOB-induced apoptosis
Discussion
The normal p53 gene is well known to play a crucial role in inducing apoptosis and acting as a cell cycle checkpoint in human and murine cells following DNA damage. 17 p21 Waf1/Cip1 protein blocks the activities of various cyclin-dependent kinases,20-22 and inhibits the phosphorylation of retinoblastoma (RB) protein, thereby preventing the G1-S phase transition.21-23 Previous studies have shown that p21 Waf1/Cip1 is transcriptionally regulated by p53-dependent and -independent pathways.24-26 Our results show that treatment of T24 cells with IOB did not result in any changes on the expression of p53. In contrast, the amount of p21 Waf1/Cip1 was increased by IOB treatment. Furthermore, flow cytometric analysis indicated that IOB could arrest T24 cells in the G0/G1 phase. Therefore, we suggest that the blockade of cell cycle progression was attributed to the amount of enhancement of p21 Waf1/Cip1 protein.
Fas/FasL system is a key signaling transduction pathway of apoptosis in cells and tissues. 12 Ligation of Fas by agonistic antibody or by its natural ligand induces receptor oligomerization and formation of death-inducing signaling complex, followed by activation of caspase-8, then further activating a series of caspase cascades resulting in cell apoptotic death.12,27 FasL is a tumor necrosis factor–related type II membrane protein. 28 Cleavage of membrane-bound Fas ligand (mFasL) by a metalloprotease-like enzyme results in the formation of soluble Fas ligand (sFasL). 29 Investigation by Mizutani and coworkers30,31 revealed that the established bladder cancer cells were resistant to apoptosis following treatment with the agonistic Fas antibody CH-11. However, bladder cancer cells were sensitized to Fas-mediated cytotoxicity by CH-11 following cotreatment with either doxorubicin or adriamycin.30,31 Our study indicated that mFasL increased in IOB-treated T24 cells. Moreover, the levels of Fas/APO-1 and the activity of caspase-8 were simultaneously enhanced in mFasL-upregulating T24 cells following by apoptotic cell death (DNA fragmentation). Furthermore, when the Fas/mFas ligand system was blocked by ZB4, a decrease in cell growth inhibition and the proapoptotic effect of IOB was noted. Similarly, cell growth inhibition and apoptotic induction of IOB decreased in T24 cells treated with caspase-8 inhibitor.
Conclusion
In summary, our study has clearly demonstrated that IOB exerts a cytotoxic activity on the T24 human bladder cancer cells. These findings are the first to show that the Fas/mFasL system plays an important role in IOB-mediated T24 cellular apoptosis. Therefore, the potential of this substance for the treatment of bladder cancer should be further investigated.
Footnotes
Chung-Yi Chen and Ya-Ling Hsu have contributed equally to the article. Dr. Hsu can be contacted at
The author(s) declared no potential conflicts of interests with respect to the authorship and/or publication of this article.
The author(s) disclosed receipt of the following financial support for the research and/or authorship of this article:
This study was supported by grants from the Chi-Mei Medical Center and Kaohsiung Medical University Research Foundation (99CM-KMU-05), and Excellence for Cancer Research Center Grant, Department of Health, Executive Yuan, Taipei, Taiwan (DOH99-TD-C-111-002).