
Abstract
Background and Objective:
We have previously reported that BRCA2 N372 H i.a.1342A>C heterozygous variation presented in platinum-resistant patients. This study aimed to further investigate the mechanism of BRCA2 N372 H mutation in the development of platinum resistance in ovarian cancer.
Methods:
The BRCA2 N372 H i.a.1342A>C was synthesized and used to exchange 1 wildtype allele followed by sequencing to confirm the mutant allele sequence. Plasmids were constructed and transfected into the OVCAR-3 cells after lentiviral packaging. BRCA2 N372 H mRNA was detected by qPCR. BRCA2 protein was assessed by immunoblotting. Binding of the BRCA2 to Rad51 was detected by immunofluorescence staining. Sensitivity of the cells to cisplatin treatment was assessed with CCK-8 assay.
Results:
It was found that expression of BRCA2 protein in ovarian cancer cells transfected with BRCA2 N372 H i.a.1342A>C gene (2.177 ± 0.003) was significantly increased compared to that of the cells transfected with lenti-EGFP only (1.227 ± 0.003, P < 0.001). Binding of the BRCA2 and Rad51 proteins was significantly increased in the cells with BRCA2 N372 H i.a.1342A>C mutation (3.542 ± 0.24) than that in the cells transfected with lenti-EGFP (1.29 ± 0.32) or empty cells (1.363 ± 0.32, P < 0.001). Cell viability significantly increased in the cells transfected with BRCA2 N372 H mutant gene. The IC50 value was significantly higher in the cells transfected with BRCA2 N372 H mutant gene (1.963 ± 0.04) than that of the cells transfected with lenti-EGFP (0.955 ± 0.03, P < 0.01) or empty cells (1.043 ± 0.007, P < 0.01).
Conclusion:
Over expression of mRNA and protein of BRCA2 was detected in the cells with BRCA2 N372 H i.a.1342A>C mutation but not in the lentivirus negative control (lenti-EGFP) or the cells without transfection (empty cells), which may lead to resistance to platinum-based drugs in ovarian cancer cells through homologous recombination repair pathway.
Introduction
Ovarian cancer is a malignant tumor that poses a serious threat to all women. Platinum-based chemotherapy is the main adjuvant therapy for advanced ovarian cancer. However, even as the most commonly used first-line chemotherapy drug, primary or secondary resistance to platinum therapy remains a major obstacle in the treatment of ovarian cancer, and it is one of the main reasons that contribute to the less than 30% of the five-year survival rate in the patients with advanced ovarian cancer. 1 Therefore, early identification and intervention may be an effective way to improve the 5-year survival rate of ovarian cancer patients. The most commonly used platinum-based drugs in clinical practice are carboplatin and cisplatin. Both carboplatin and cisplatin are cell-cycle nonspecific drugs, mainly acting on the N7 and O6 atoms of guanine, causing DNA interchange and intra-strand crosslinking, preventing helix melting, and interfering with DNA synthesis resulting in cytotoxicity.
The BRCA2 gene is a tumor suppressor gene located at 13q12-13 and plays an important role in the development, progression, and prognosis of ovarian cancer and breast cancer. BRCA2 and RAD51 homologous protein physically interact with each other during homologous recombination (HR), and jointly participate in DNA damage repair and recombination. 2 In this regard, when DNA double strands are damaged, DNA damage signal activates kinases such as ATM (ataxia-telangiectasia, mutated) and ATR (ATM and Rad3-related) to catalyze the phosphorylation of the BRCA2/RAD51 protein complex, 3 thus converting it from an inactive form to an active form. Double strand breaks of DNA can be repaired by a HR-based error-free mechanism, which is carried out by homology search and strand invasion to complete the repair. 4,5 RAD51 is the key protein required for the initial strand invasion step, and this invasion step is dependent on several proteins such as BRCA1 and BRCA2. 4,6 The BRCA2 protein carries the RAD51 protein to the damage site of the double-stranded DNA and participates in the repair process. 7 There are 3 described genotypes in BRCA2 N372 H variant, that is, wild type (AA), heterozygous (AC) and homozygous (CC). In our previous studies, the BRCA2 N372 H i.a.1342A>C gene heterozygous variation was specifically found in the plasma of the patients with drug-resistant recurrence of ovarian cancer compared with platinum-sensitive patients. 8 Therefore, we hypothesized that BRCA2 N372 H gene mutation may be associated with platinum resistance in ovarian cancer. The current study was, therefore, designed to explore the effect of BRCA2 N372 H mutation gene on platinum-resistance in the ovarian cancer cells. Since OVCAR-3 cell line is a known and widely used ovarian cancer cell line, it was used for the investigation of BRCA2 gene mutation effect on cell survival and function in the current study.
Materials and Methods
Experiment Design
We first synthesized the BRCA2 N372 H i.a.1342A>C (rs144848) heterozygous mutant gene using gene synthesis technology and sequenced the BRCA2 N372 H gene fragment. Then constructed OVCAR-3 ovarian cancer cells with BRCA2 N372 H i.a.1342A>C mutation, and cell clones carrying the target gene were screened out by monoclonal culture. Identification of the mutant gene was performed by digital PCR and BRCA2 protein expression was determined by immunoblotting. The OVCAR-3 cells without transfection were used as positive cells that constitutively expressed BRCA2. Finally, the binding of the RAD51 protein to BRCA2 protein in the target cells was detected by immunofluorescence. Commercially available CCK-8 assay kit, which uses a water-soluble tetrazolium salt to quantify the number of live cells, was used to detect the resistance of BRCA2 N372 H mutant cells to cisplatin, and the mechanism by which BRCA2 N372 H mutation affecting the sensitivity of cells to platinum-based drugs was determined.
Cell Strain and Culture
The human OVCAR-3 cell line was purchased from a commercial source (Shanghai Suer Biotech, Shanghai, China). Cells were cultured with RPMI-1640 (GIBCO, Invitrogen Corporation, Shanghai, China) containing 10% fetal bovine serum (FBS) in 5% CO2, 95% humidified air, and 37°C incubator.
BRCA2 N372H Mutant Gene Synthesis and Sequencing
The BRCA2 N372 H mutant gene was obtained by gene synthesis and amplification (Generay, Shanghai, China) using PCR specific for the BRCA2n372 h gene fragment. Sequence of BRCA2 has been provided as supplemental material. The BRCA2 N372H-F1 primer sequence was GCCAAATGTCCTAGAAGATGAAG and the BRCA2 N372H-R1 primer sequence was CTGTTTCCTCATTTAATGGCTTC. Genomic DNA was extracted from samples using a genomic DNA extraction kit (AxyPrep Mini DNA extraction kit, Baisai Biology Technology, Shanghai, China) and subjected to electrophoretic identification. Using genomic DNA as a template, the target fragment was amplified with a high-fidelity enzyme. The PCR cycle was as following: 95°C 5 min, 1 cycle, 95°C 30 sec, 58°C 30 sec, 72°C 45 sec, 30 cycles, 72°C 10 min, 1 cycle, after which PCR products were recovered (AxyGen PCR purification kit, Baisai Biology Technology, Shanghai, China) and sequenced.
Construction of BRCA2 N372 H Mutant Gene Vector
Primers were designed based on the sequence of the target gene, and the restriction sites Asc1 and Pme1 were added upstream and downstream of the target fragment, respectively, with BRCA2 N372H-F primer sequence TCAAGTTCTTTAGCTACACCACCC and BRCA2 N372H-R primer sequence TTCTTTTTTAAAGTTTGGATCAGTCA. The gene was used as a template to amplify the target fragment by PCR. As shown in the Supplement Figure 1, appropriate restriction sites were added to both ends of the target fragment. The PCR cycle was as following: 95°C 3 min, 1 cycle, 94°C 30 sec, 58°C 30 sec, 68°C 60 sec, 30 cycles, 68°C 10 min, 1 cycle. The PCR product was separated by agarose gel electrophoresis, and the target fragment was recovered using the AxyGen Gel Recovery Kit. The target gene PCR product and the target vector were digested with Asc1 and Pme1, respectively. After being digested at 37° C for 4-5 h, the fragments were electrophoretically separated and the target fragment was excised and recovered from the gel. The T4 DNA ligase ligated the digested PCR product and target vector.

Infection of OVCAR-3 Cells With Lentivirus Carrying BRCA2 N372 H Mutant Gene
The bacteria solution containing target gene plasmids was used for streak culture. Plasmids were extracted using the AxyGen Plasmid Extraction Kit (Baisai Biology Technology, Shanghai, China) and were packaged in lentivirus. OVCAR-3 cells at the logarithmic growth phase were trypsinized, plated in 6-well cell culture plates, and cultured at 37°C with 5% CO2 until cell fusion reached 40%-50%. Then, 55 µl of the target virus solution (lenti-BRCA2-EGFP, titration of 2.54 x 107 TU/mL) or 25 µl of negative control virus solution (lenti-EGFP, titration of 108 TU/mL), and Polybrene was added at a final concentration of 8 µg/ml for transfection. The cells were incubated with the viral solution for 48 h.
Detection of BRCA2 N372 H Mutant Gene With RT-qPCR in OVCAR-3 Cells Following Transfection
Quantification of BRCA2 N372 H mutant gene in the OVCAR-3 cells was performed following the MIQE guidelines. 9 A total of 0.5 ml Trizol was added to each well. Total RNA was extracted and reverse-transcribed into cDNA after treatment with DNase I. Quantitative PCR (qPCR) was used to detect expression of BRCA2 gene, with BRAC2 N372H-F2 primer sequence TGGAGCCCAGATGGAGAAA and BRAC2 N372H-R2 primer sequence AGTTCCAGATATTGCCTGCTTT. ACTB was used as reference gene and used for data normalization with the following primer sets: hACTB-2F: TCCTTCCTGGGCATGGAGT and hACTB-2 R: CAGGAGGAGCAATGATCTTGAT. The PCR condition was as following: 95°C, 2 sec, 1 cycle (1X); 95°C, 15 sec, 60°C, 2 sec, 72°C, 2 sec, 2 cycles (40X), 60°C-95° C, 3 sec, 3 cycles (39X).
Identification of the BRCA2 N372 H i.a.1342 A>C Gene Mutation in OVCAR-3 Ovarian Cancer Cell Strains
The BRCA2 N372 H mutant gene was obtained by gene synthesis and the 1342 “A” to “C” mutation (N372 H, rs144848) was confirmed by sequencing (Figure 1A), which was compared with variant type of BRCA2 N372 H gene locus in in the wild type cells (Figure 1B, Illumina HiSeq 4000 high-throughput sequencing platform), cells transfected with negative virus control (lenti-EGFP, Figure 1C), or cells without transfection (empty cells, Figure 1D).
Immunoblotting
After washing once, boiled protein loading buffer (120 µl) was added to each well to lyse the cells. The sample was boiled at 100°C for 10 min, centrifuged at 12000 rpm (14 cm diameter of the centrifuge) for 1 min, and stored at −20°C after dispensing. Proteins were loaded and electrophoresis was performed at constant pressure 80 V for 30 min followed by constant pressure 120 V for 1 h. Distance between the 2 electrodes was 12 cm. The protein was then transferred to the PVDF membrane and blocked with the blocking solution (containing 5% skim milk powder in TBST) for 1 h at room temperature. The membrane was then incubated with the primary antibody against BRCA2 (1:200, Abcam, Cat#: ab27976) at 4°C overnight. Next day, the secondary antibody (Anti-mouse second antibody, Jackson, Cat #: 315-035-003; or anti rabbit secondary antibody, Jackson, Cat#: 111-035-008) was applied at room temperature for 1 h. After washing, protein band was visualized, and densitometry analysis was performed.
Detection of RAD51 Protein by Confocal Microscopy
The cells were fixed with 4% paraformaldehyde for 15 min followed by rinsing twice with PBS. Cells were then permeabilized with 0.5% Triton-100 in PBS for 10 min followed by blocking with 4% BSA in PBS at room temperature for 1 h. Primary anti-RAD51-antibody (1:100, Abcam, Cat#: ab133534) was allowed to react at 4°C overnight. After washing 5 times with PBS, goat anti-rabbit Alexa-594 secondary antibody (Abcam, Cat#: ab150080) were applied for 1 h at room temperature. After washing with PBS, nuclei were stained with DAPI, and the slides were sealed and observed using a confocal microscope (ZEISS, LSM710). Fluorescence intensity (IntDen value) was obtained using the Image J software following the manufacturer’s instruction (NIH, Public Domain, USA).
Assessment of Cellular Sensitivity to Cisplatin by CCK-8 Assay Kit
Sensitivity of the cells to cisplatin was assessed using CCK-8 kit following the manufacturer’s instruction (Dojindo, Cat#: CK 04). The transfected cells and non-transfected cells were trypsinized after 72 h culture. The cell suspension was seeded in 96-well culture plates (8000 cells/well) and incubated at 37°C with 5% CO2. Cisplatin was added to the cell culture plates at concentrations of 0, 0.05, 0.1, 0.5, 1, 2, 4, and 8 µg/ml. After 24 h and 48 h treatment, respectively, 10 µl of CCK-8 was added to each well and incubated for 2 h followed by measuring the absorbance at 450 nm with a microplate reader (Thermo, MuLTiSKAN MK3).
Statistical Analysis
Image J software was used to analyze the immunofluorescence intensity for the binding of RAD51 protein expression. An unpaired t test in the Prism 7.0 software package was used to compare the differences of mRNA expression and semi-quantification of protein levels. SPSS23.0 was used to analyze the IC50 of each group in the CCK-8 experiment.
Results
Quantification of BRCA2 N372 H Mutant Gene mRNA in the 3 Strains of OVCAR-3 Cells
Following the co-transfection of BRAC2 N372 H mutant gene expression vectors and the packaged plasmids into the OVCAR-3 cells, expression of BRCA2 N372 H mRNA in OVCAR-3 cells was quantified by RT-qPCR. As shown in the Figure 2, mRNA expression of BRAC2 N372 H mutant gene was 7000 fold higher in the cells transfected with BRAC2 N372 H mutant gene (7625.8 ± 30.5) compared to that in the cells either non-transfected (empty cells, 1.00 ± 0.12, P < 0.0001) or cells transfected with Lenti-EGFP (7.16 ± 0.15, P < 0.0001).

Comparison of BRCA2 Protein Expression Following Transfection of BRCA2 N372 H Mutant Genes in the OVCAR-3 Cells
Protein level of BRCA2 was detected by immunoblotting in the cells transfected with the BRCA2 N372 H mutant gene, the lentivirus negative control (Lenti-EGFP), or the OVCAR-3 cells without any transfection (empty cells). As shown in Figure 3, the OVCAR-3 cells transfected with the BRCA2 N372 H mutant gene showed significantly increased BRCA2 protein expression (Ratio of BRCA2/tubulin: 2.177 ± 0.003) compared to that of the cells transfected with Lenti-EGFP (Ratio of BRCA2/tubulin: 1.227 ± 0.003, P < 0.001).

Comparison of RAD51 Protein Expression
In order to assess the effect of BRAC2 N372 H gene mutation on the physical binding of BRCA2 protein and RAD51 protein in OVCAR-3 cells, expression of RAD51 protein in the OVCAR-3 cells transfected with virus expressing the BRAC2 N372 H mutant gene and Lenti-EGFP was examined by immunofluorescence staining. As shown in the Figure 4, the immunofluorescent intensity of RAS51 protein in OVCAR-3 cells transfected with expressing the BRAC2 N372 H mutant gene (3.542 ± 0.24) was significantly stronger than that in the cells either transfected with Lenti-EGFP (1.29 ± 0.32) or the cells without transfection (empty cell) groups (1.363 ± 0.32, P < 0.001).
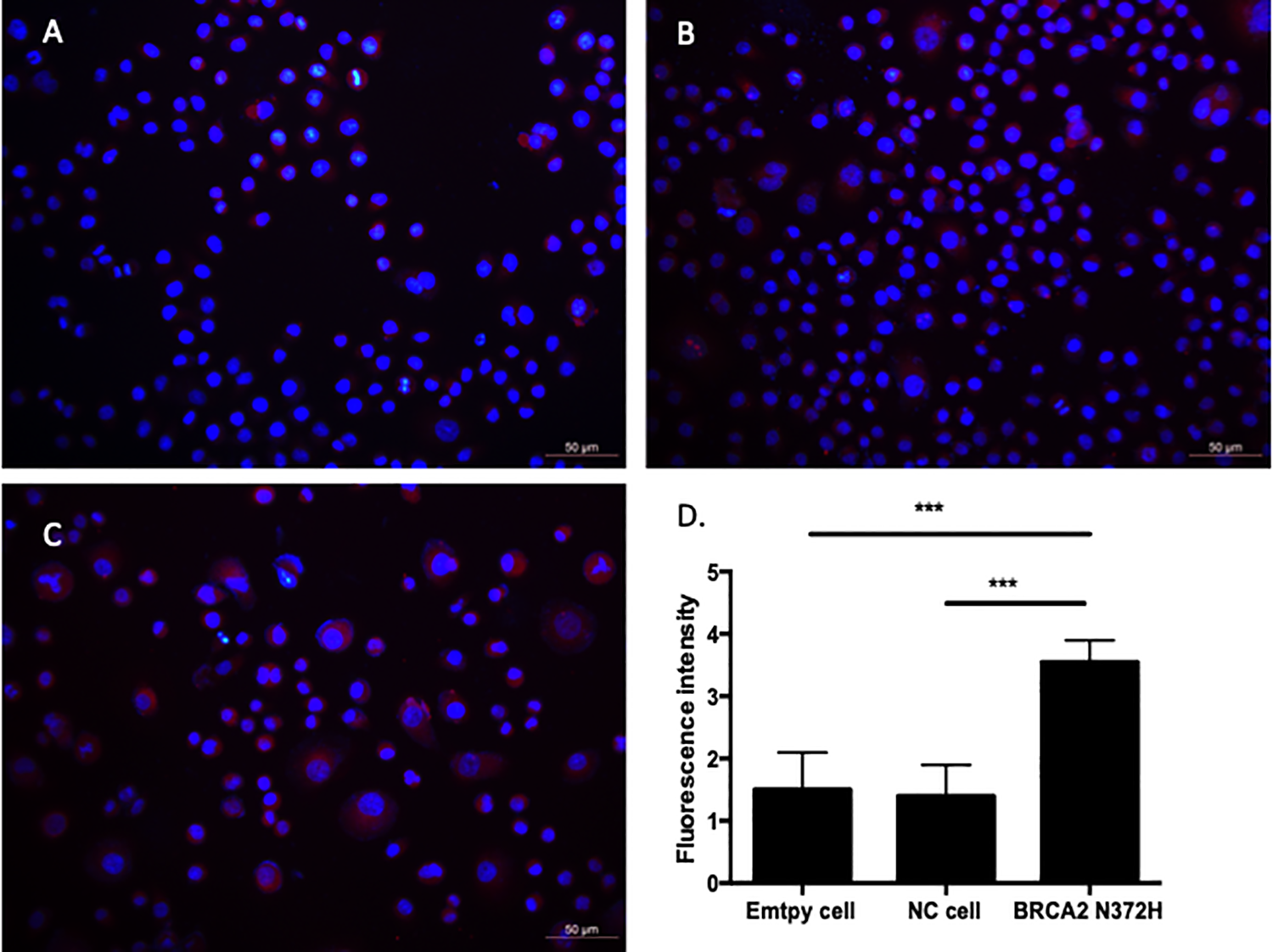
Cell Viability in Response to Cisplatin and IC50 in 3 Cell Strains
Following transfection of BRCA2 N372 H mutant gene and negative control Lenti-EGFP vectors, the OVCAR-3 cells were treated with different concentrations of cisplatin (0.05, 0.1, 0.5, 1, 2, 4, and 8 µg/ml) for 24 and 48 h, respectively. Cell viability and the IC50 of the cells in response to cisplatin were assessed by CCK-8 kit. As shown in Figure 5, cell viability was significantly higher in the cells that transfected with BRCA2 N372 H mutant gene compared to that of the cells transfected with Lenti-EGFP or empty cell groups when they were treated with different concentrations of cisplatin at 24 h (Figure 5A) or 48 h (Figure 5B). After 24 h treatment, the difference was especially significant at cisplatin concentrations of 2, 4, and 8 μg/ml, and after 48 h, the difference was significant at concentrations of 1, 2, 4, and 8 μg/ml. The IC50 value was significantly higher in the cells transfected with BRCA2 N372 H mutant gene (1.963 ± 0.04 at 24 h and 1.299 ± 0.06 at 48 h) than that of the cells transfected with Lenti-EGFP (0.955 ± 0.03 at 24 h and 0.575 ± 0.02 at 48 h, P < 0.01) or non-transfected control (empty) cells (1.043 ± 0.007 at 24 h and 0.593 ± 0.01 at 48 h, P < 0.01, Figure 5C).

Discussion and Conclusion
In the TCGA database, rs144848 (GG) homozygotes and (GT) heterozygotes cause a significantly increased risk of ovarian cancer, breast cancer, and prostate cancer. For this special single nucleotide polymorphism (SNP
We used gene synthesis technology to synthesize the BRCA2 N372 H i.a.1342 A>C mutant gene, which was confirmed by sequencing. Then, BRCA2 N372 H mutant gene vectors were constructed and packaged with lentivirus to transfect OVCAR-3 cells. Monoclonal culture of BRCA2 N372 H i.a.1342 A>C mutant OVCAR-3 cell lines was performed and the expression of large amounts of BRCA2 N372 H mRNA was detected using the RT-qPCR method, demonstrating the BRCA2 N372 H i.a.1342 A>C mutant OVCAR-3 cell line was successfully constructed. In addition, expression of BRCA2 protein in OVCAR-3 cells with BRCA2 N372 H i.a.1342 A>C mutation was significantly higher than that in the cells transfected with Lenti-EGFP or the cells without transfection, indicating that BRCA2 N372 H i.a.1342 A>C mutation can increase the expression of BRCA2 protein.
Homologous recombination (HR) is one of the major mechanisms in DNA double-strand break repair pathways. The crucial step of HR is homologous strand exchange directed by the protein RAD51, which is recruited by BRCA2. 5,13 BRCA2 regulates both the intracellular localization and DNA binding ability of RAD51, and loss of these controls following BRCA2 inactivation may be a key event leading to genomic instability and tumorigenesis. 14 In the current study, therefore, expression of RAD51 was examined by immunofluorescence staining in the 3 cell strains. The purpose of this design was to test the function of the mutant BRCA2 protein in OVCAR-3 cells and verify whether these BRCA2 proteins can effectively bind to RAD51 protein and exert DNA damage repair function. The results showed that the fluorescence intensity of RAD51 protein in OVCAR-3 cells with BRCA2 N372 H i.a.1342 A>C mutation was higher than that in the cells transfected with Lenti-EGFP or the cells without transfection (empty cells), demonstrating that the mutated BRCA2 protein could bind to the RAD51 protein.
Since recent studies also indicated that the BRCA2 N372 H mutation lead to drug resistance in breast cancer cell strains, 3,15 survival of the 3 cell strains in response to cisplatin was examined by CCK8 assay. It was found that cell viability of the OVCAR-3 cells transfected with the BRCA2 mutant was significantly higher than that of the cells transfected with Lenti-EGFP or the cells without transfection (empty cells) at 24 or 48 h, with a more pronounced difference as the concentration of cisplatin was increased. Furthermore, in the OVCAR-3 cells with BRCA2 N372 H i.a.1342 A>C mutation, the IC50 of cisplatin was significantly higher than that of the cells transfected with Lenti-EGFP or the cells without transfection (empty cells). These findings demonstrated that BRCA2 N372 H i.a.1342A>C mutation in the OVCAR-3 cells resulted in significant increase of resistance to cisplatin.
Consistent with our findings, many researchers suggested that secondary mutation of BRCA2 may be an important mechanism for platinum resistance in BRCA2 mutations. 16 -19 This interaction occurs at the BRCA2 exon 11 binding site or the so-called BRCA2 DBD (DNA/DSS1-binding domain) domain on the carboxy terminus of BRCA2. 20 In BRCA1 and BRCA2 ovarian cancers that carry frameshift mutations, acquired platinum resistance is generated in the gene by secondary mutations, by which the open reading frames (ORF) of mutated BRCA1/2 are corrected, and the wild-type BRCA1/2 expression and DNA homologous recombination repair function are recovered. In the pancreatic or ovarian cancer cell lines with BRCA2 mutations, secondary mutations in BRCA2 are induced by cisplatin or poly ADP-ribose polymerase (PARP) inhibitors. 19 As a result of these secondary mutations, the truncated BRCA2 protein is reconnected and carries out the function of homologous recombination. 18 In addition to the regulation of homologous recombination repair genes, it has also been reported that the development of platinum resistance may be attributed to other mechanisms since BRCA2 mRNA expression is not detected in 13% of ovarian cancer cases. 21 In this regard, we found no truncating mutations in the BRCA2 gene in the first tumor specimen of 4 patients who had drug resistant ovarian cancer recurrence and the BRCA2 N372 H i.a.1342 A>C mutation. This discrepancy with results reported by other researchers suggested that the N372 H i.a.1342 A>C mutation may increase the resistance to platinum-based drugs in tumor cells through different pathways.
Therefore, we have further explored the potential mechanisms and found that increased physical binding of BRCA2 protein and RAD51 protein may contribute to the regulation of BRCA2 N372 H i.a.1342 A>C mutation on platinum sensitivity in the cells. Specifically, binding of the aforementioned 2 proteins may increase the homologous recombination repair function of the cells, and lead to the development of platinum resistance in ovarian cancer cells, which was confirmed by the CCK-8 experiment.
In addition, studies have also suggested that expression of ATP-binding cassette (ABC) transporters in cancer cells can confer resistance to cytotoxic and targeted chemotherapy. 22 -24 ABC transporters regulate cellular transportation of variety types of molecules including hormones, lipids, ions, and other small molecules across cell membranes. On one hand, loss of function of a particular ABC transporter is associated with a number of heritable diseases such as cystic fibrosis. 25 On the other hand, ABC transporters are also associated with drug resistance in variety kinds of cancers including ovarian cancer. 23 In this regard, whole-genome sequencing of 92 patients with high-grade serous ovarian carcinoma with primary and matched resistant disease showed that reversion of BRCA1 and/or BRCA2 mutations were potential mechanisms of drug resistance. 15 Whether expression of ABC transporters is altered in the cells with BRCA2 N372 H i.a.1342 A>C mutation remains to be determined.
One of the limitations of this study was that a cell line with overexpressing wild type BRCA2 was not applied as control for comparison with BRCA2 N372 H mutation although the “empty cells” OVCAR3 ovarian cell that contains wild type BRCA2 gene was used. However, it has been reported that exogenous expression of wild type BRCA2 in a human pancreatic cancer line (Capan-1 cells) conferred resistance to methyl methanesulfonate treatment, 26 and that complex of BRCA2 and RAD51 in breast cancer MCF-7 cells was diminished upon conditional expression of a wild type, but not a mutated, BRC4-repeat. 27
The BRCA2 N372 H i.a.1342 A>C mutation increased the expression of BRCA2 protein in ovarian cancer cells. This mutation-associated BRCA2 protein up-regulation resulted in enhanced binding of BRCA2 and RAD51 proteins and augmented DNA homologous recombination repair function of ovarian cancer cells, and by which mechanism, it led to the development of resistance to platinum-based drugs. While these in vitro experimental results need to be further confirmed in vivo with large population screening of the patients with drug-resistant recurrence of ovarian cancer, findings of this study may provide the theoretical basis for drug development and targeted therapy of ovarian cancer.
Conclusion
Taken together, the current study revealed that the BRCA2 N372 H i.a.1342 A>C mutation could regulate the sensitivity of ovarian cancer cells to platinum-based drugs.
Supplemental Material
Supplemental Material, Supplement_material - The BRCA2 p.N372 H i.a.1342A>C Could Regulate the Sensitivity of Ovarian Cancer Cells to Platinum-Based Drugs
Supplemental Material, Supplement_material for The BRCA2 p.N372 H i.a.1342A>C Could Regulate the Sensitivity of Ovarian Cancer Cells to Platinum-Based Drugs by Zhen-Hua Du, Yu Xia, Qing Yang and Song Gao in Technology in Cancer Research & Treatment
Supplemental Material
Supplemental Material, SupplmentFig - The BRCA2 p.N372 H i.a.1342A>C Could Regulate the Sensitivity of Ovarian Cancer Cells to Platinum-Based Drugs
Supplemental Material, SupplmentFig for The BRCA2 p.N372 H i.a.1342A>C Could Regulate the Sensitivity of Ovarian Cancer Cells to Platinum-Based Drugs by Zhen-Hua Du, Yu Xia, Qing Yang and Song Gao in Technology in Cancer Research & Treatment
Footnotes
List of Abbreviations
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) declared the following potential conflicts of interest with respect to the research, authorship, and/or publication of this article: Experimental study on the relationship between BRCA2 N372H mutation and platinum resistance in ovarian cancer, Science and Technology Department of Liaoning Province, No: JC2019018.
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.