
Abstract
This review provides an evaluative overview of five concepts specific to developmental and neurobiological psychology that are found to be largely overlooked in current textbooks. A sample of 19 introductory psychology texts was surveyed to develop a list, including glial cell signaling, grandmother cells, memory reconsolidation, brain plasticity, and moral judgements by infants. These topics are relatively new, have proven to be of high impact in their respective fields, but are rarely discussed in psychological textbooks or by instructors in the classroom. In addition to a brief, but detailed background on each of the concepts, potential textbook chapters and classroom topics that would benefit from a discussion of these concepts are identified. Finally, this review briefly addresses possible ways for textbook authors to incorporate these new topics in future editions of texts without drastically increasing the overall length of the text.
Keywords
Introduction
In 1875, William James brought the field of psychology to America. Within a few years, James contracted to write a textbook on the subject of experimental psychology, which would later be published as The Principles of Psychology. Had James completed this textbook in a timely manner, it would have been the first text of its kind. However, other universities were adopting the teaching of psychology, and so the race to publish psychology textbooks began. Today, there are nearly 1.6 million psychology students per year in the market for introductory psychology texts (Steuer & Hamm, 2008) and approximately 98% of instructors use textbooks in their introductory psychology courses (Griggs, 2014). With scores of textbook options to choose from, a fair amount of diversity among the texts and the information provided within them would be expected.
Textbooks Examined
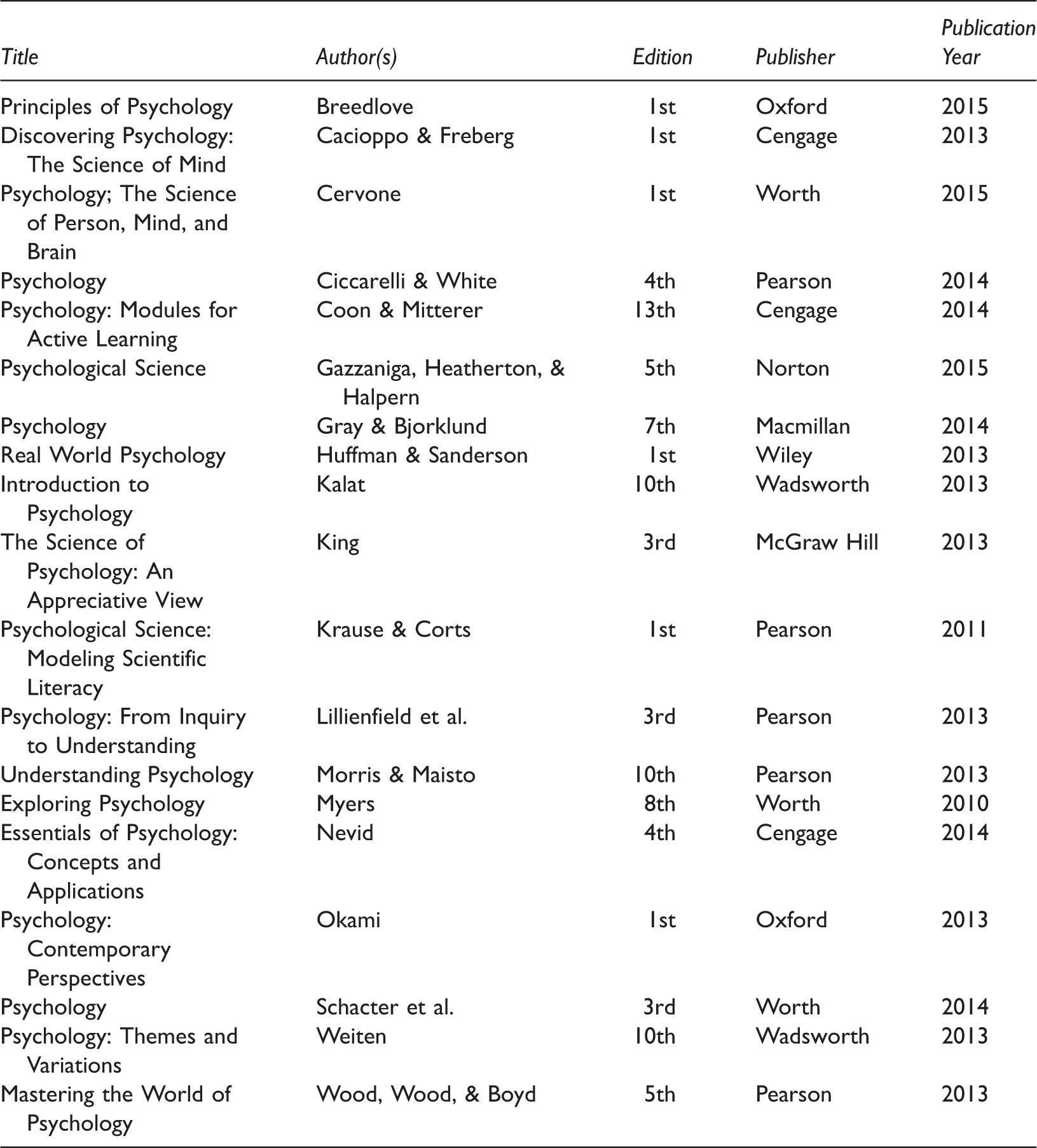
Based on our experience in research and instruction, we developed a list of concepts in the neurobiological and developmental areas that we considered to be relatively new, have proven to be of high impact in their respective fields, but are rarely discussed in textbooks or instructors in the classroom. This list, which is not intended to be comprehensive, included in the neurobiological domain glial cell signaling, mirror neurons, epigenetics, grandmother cells, brain plasticity, and memory reconsolidation. The developmental concepts included moral judgements by infants, punishment’s effect on children’s motivation, theory of mind, epigenetics, and brain plasticity. Brain plasticity and epigenetics are included in both groups owing to their importance to both fields.
Following review of the 19 texts, mirror neurons and epigenetics were removed from the neurobiological list and punishment’s effects on children’s motivations, epigenetics, and theory of mind were removed from the developmental list because they were found to be fairly well represented in most, although not all, of the reviewed texts; the remaining concepts—glial cell signaling, grandmother cells, brain plasticity, memory reconsolidation, and infant moral judgment—were not. Given the popularity of these topics in not only emerging literature, but also literature dating back up to 50 years, it is surprising that these concepts are rarely included in introductory textbooks. For example, since the original introduction of glial cell signaling in psychological journals in 1991 (publications not specific to the role of glial cells in neural signaling date back as far as 1922), there have been over 300 publications on the topic. Of those 300, 251 were published in the last decade, and 34 in the last year, indicating substantial growth of this topic in the field.
The term “grandmother cells” did not yield more than 30 results in psychology-related journals. However, the early seminal paper by Gross, Bender, and Rocha-Miranda (1969) (which discusses grandmother cells under the term visual receptive fields) has been cited over 1,300 times in psychological journals, of which 627 occurred in the last 10 years and 93 in the last year. Memory reconsolidation, a topic not introduced in psychological journals until 1993 (Rodriquez, Phillips, Rodriquez, & Martinez, 1993) yielded 780 results, 732 of which were published in the last 10 years and 179 in the last year.
The concept of moral judgement by children, first developed by Piaget (1932) and more thoroughly by Kohlberg and Kramer (1969), is outlined in every textbook reviewed for this paper. However, Hamlin and colleagues (2007, 2010, 2011) have focused on moral behaviors in infants for the last decade and have published a plethora of data regarding infant moral capabilities. Their initial findings (2007) have been cited in 823 different psychological publications. To provide a comparison, the original paper that outlines Kohlberg’s theory (1969) has been cited by only 741 different psychological publications.
Finally, brain plasticity as a general topic yields more than 3,300 results and specific divisions of structural and functional plasticity generate 940 and 960 results, respectively. Of the 960 functional plasticity results, 172 were published between the initial publication year of 1969 and 2006, with 788 appearing in the last decade and 181 in the last year. This means that more literature on this topic was published in the last year than was published in the first 36 years following the initial discussion of the topic. This trend is also paralleled by structural plasticity publications.
To provide a comparison between these identified topics and one of the most well-known topics in psychology, classical conditioning yields 16,000 results. Of those, 4,081 were published in the last decade and 429 in the last year. Although these are substantial numbers (and definitely influential enough to justify maintaining this topic’s place in introductory texts), there is not an increasing trend to indicate popularity growth as has been found in our suggested topics. Below, each of the concepts that we found to be largely overlooked is discussed in order to highlight their importance to the field of psychology and promote awareness of topics that may not be discussed in introductory psychology courses.
Glial Cell Signaling
Virtually all introductory psychology textbooks discuss glial cells. This discussion is invariably about the role that glial cells play as the insulating myelin sheath that helps speed up neural transmission. Few texts go beyond this, and if they do, the janitorial duties of glial cells are typically what are mentioned. The term glia itself encapsulates this diminutive concept because it means “glue” in Greek.
Douglas Fields, Chief of the Section on Nervous System Development and Plasticity at the National Institute of Child Health and Human Development, is determined to rehabilitate the reputation of glial cells. Fields has founded a journal, Neuron Glia Biology, and published a book titled The Other Brain (2009) in an attempt to disseminate research findings from the past two decades that have shown that glial cells are more than “mental bubble wrap” (p. 7).
Fields begins his book with an intriguing story about how Marian Diamond, the renowned neuroanatomist from the University of California – Berkeley, examined four slices of cerebral cortex from the brain of Albert Einstein. Diamond compared Einstein’s brain to those of 11 other men, finding that the average number of neurons was the same for Einstein’s cortex as it was for the others. Where Einstein’s brain differed was in the number of glial cells: Einstein’s cortex had twice as many as the other brains. Although this discovery does little to prove that glial cells formed the basis of Einstein’s genius, Fields uses this story to introduce what he feels is one of nature’s best-kept secrets.
Perhaps the most important point that Fields (2009) makes is that glial cells, especially astrocytes, contribute significantly to neural signaling. The old model of the synapse included only neurons: On the transmitting side was the presynaptic membrane from which neurotransmitters such as glutamate were released. On the receiving side was the postsynaptic membrane in which were receptors to bind the glutamate, thus causing depolarization in that neuron and the subsequent transmission of information between neurons. However, recent research has shown that each synapse typically involves, in addition to neurons, a type of glial cell called an astrocyte, which also has receptors for glutamate as well as the ability to release glutamate into the synapse (Volterra & Meldolesi, 2005). These processes allow the astrocyte to modulate neural transmission in synapses. Consequently, the old model of the synapse that considers only neurons has been replaced by one which includes astrocytes. Hence, the term “tripartite synapses” has been coined to represent this new concept (Pannasch & Rouach, 2013).
Fields (2009) and others (Pannasch & Rouach, 2013; Volterra & Meldolesi, 2005) have also noted other recently discovered functions for glial cells, including calcium signaling, regulation of blood flow, and their roles in chronic pain and neurodegenerative diseases. Fields predicts that as understanding of glial cells advances, understanding of the brain and its functions will also radically change. We agree and feel that students of psychology should begin learning about this new brain from their earliest introduction to our discipline.
Glial cells are almost exclusively referred to only in the neurobiological chapter. A second occasion to present this concept might be in the sensation and perception chapter for their contribution to pain regulation and the treatment of chronic pain. Fields (2009) also notes that learning is influenced by the way that glial cells help to increase the formation of new synapses. Memory, too, is contributed to by release of the neurotransmitter glutamate by glial cells in the hippocampus, a crucial structure for the encoding of new experiences. Fields’ statement that Einstein’s cortex was found to have twice as many glial cells dovetails with animal studies that have found that the brains of rats raised in enriched environments have up to 33% more glial cells than those raised in impoverished environments. Although exactly how this abundance of glia might improve intelligence is not yet known, it likely has to do with the way that these cells help to integrate neuronal functioning within and between large areas of the cerebral cortex.
Grandmother Cells
Grandmother cells, yellow-Volkswagen detectors (Weisstein, 1973), or Halle Berry neurons as they have been referred to in the popular media (Sanders, 2009), are neurons in the cerebral cortex thought to respond to highly specific stimuli (Gross, 2002). Early discovery of grandmother cells can be credited to Gross and colleagues (1969), who found that a macaque monkey’s inferior temporal cortex contained visual neurons that selectively responded to hands and faces. Named grandmother cells because these cells appear to become active only when an individual is presented with a specific, complex stimulus, their role in the neurobiology of perception has often been interpreted to imply that a single cell in the brain can distinguish and react to the image of one’s grandmother or Halle Berry, even though one has never met Ms. Berry in person (Weisstein, 1973). One alternative explanation for these findings is that a given grandmother cell in and of itself has no specific features to which it responds, but does so only as part of a large neural ensemble, made up of perhaps thousands of neurons. This ensemble serves as the neural representation of one’s grandmother; recording from just one cell in that ensemble might make it appear that this individual neuron alone is responding to a unique, complex stimulus but that neuron is a small part of a group of cells that fire in a particular pattern in response to a particular face.
There is a middle ground, of course. The Nobel-prize-winning work of Hubel and Wiesel (1959)—research that is well represented in introductory psychology textbooks—has shown that neurons in the mammalian visual system do respond preferentially to specific features, although these tend to be elementary features such as line segments at certain orientations, rather than stimuli as complex as a face. Thus, the middle-ground hypothesis is that complex stimuli (specific faces) are represented in the brain by neural ensembles, but each neuron contributes a specific feature to that ensemble.
After several decades of debate, Freiwald, Tsao, and Livingstone (2009) carried out an investigation that attempted to settle the matter, albeit with non-human primates. Using functional magnetic resonance imaging (fMRI), Freiwald et al. (2009) first identified an area in the temporal lobe of macaque monkeys that responded to faces; they then verified that cartoon faces activated this area nearly as often as real faces. Cartoon faces allowed the experimental control of seven facial features, including face outline, eye outline, eyes, mouth, hair, nose, and eyebrows. Freiwald et al. then searched for individual cells that responded to at least one of these seven varied features. Perhaps their most important finding was that no neuron responded to all seven of the facial features, i.e., no grandmother cells were found. Although it is impossible to prove a negative without recording from every neuron in the brain, nevertheless there was no evidence in support of the grandmother cell theory. On the other hand, many neurons responded systematically to variations in at least one of the seven facial features and several neurons responded to up to four such features. This rules out the model of face perception that relies solely on neural ensembles without feature detectors; the data from this study support the middle ground. As the authors conclude, “[t]he mechanism for distinguishing faces appears to rely on a division of labor among cells tuned to different subsets of facial features” (Freiwald et al., 2009, p. 1194). In other words, a face is represented by a group of neurons, many of which have fairly high specificity for important facial features.
Perhaps one reason why this topic has been left out of introductory psychology texts is that it is quite complex. The concept of grandmother cells in and of itself is not difficult, but the idea of neural ensembles is. However, using the Freiwald et al. (2009) study as the basis, this topic can be presented as an evolving scientific controversy that has a rich history and an interesting outcome. It may also help to convey one of the most difficult concepts of all: How the wet matter of the brain converts neural signals into perceptual experiences. (Note: For an excellent summary of the methods and findings of Freiwald et al., 2009, see Jagadeesh, 2009.)
Grandmother cells (and neural ensembles) can be easily integrated into the discussion of perception in that their primary function deals with the organization, identification, and interpretation of faces. Furthermore, after being introduced in the chapter covering perception, this theory can also support discussion of infant-maternal bonds and social referencing in the emotion and development chapters as it provides neural evidence of the ability to identify specific faces, a component important to social referencing behaviors, which may aid in infant-mother pair bonding.
Memory Reconsolidation
Consolidation, the stabilization of memories in the brain, is a process that involves changes to receptor sites, synapse sensitivity, and the proteins contained in the neuron (McGaugh, 2000). This process works to transfer and stabilize short-term memories into long-term storage (Tronson & Taylor, 2007). According to the standard model of consolidation, prior to the transfer of memories into long-term storage, encoding can be easily disrupted. However, once a memory is consolidated in the brain, a process that may vary in duration depending on the complexity of the memory being consolidated (Dudai, 2004), it is stable and highly resistant to disruption or change (Nader & Hardt, 2009).
Memory reconsolidation, as the name implies, is the process of storing a memory after it has been retrieved. According to Nader (2003), “[r]eactivation of a consolidated memory can return it to a labile, sensitive state—in which it can be modified, strengthened, changed or even erased” (p. 226). It other words, it was previously thought that once a memory was consolidated, it could no longer be altered. However, recent research suggests that once a memory is reactivated, it can potentially be altered and reconsolidated into long-term memory (Dudai, 2012; Nader & Hardt, 2009). A good analogy here is that of a computer text file. Once closed and saved, that file is stored safely on our computer. However, once individuals reopen, i.e., reactivate, that file, it is vulnerable to alterations, which then get stored when they again save and close the file.
A number of studies carried out with rodents in the 1960s and ’70s demonstrated how the reactivation of a memory makes it vulnerable to modification (e.g., Misanin, Miller, & Lewis, 1968). Schiller and colleagues (2010) more recently demonstrated this in human participants by classically conditioning a fear response (measured by skin conductance). Here, the conditioned stimulus (CS) was a colored square and the unconditioned stimulus (UCS) used to evoke the fear response was a shock to the wrist. After conditioning, Schiller et al. reactivated the memory by giving one presentation of the CS and then a number of extinction trials. Schiller et al. were attempting to reactivate the memory, destroy the conditioned emotional aspect of it, and reconsolidate that memory. They found that the extinction group showed no spontaneous recovery either one day or one year following reconsolidation. Control groups showed spontaneous recovery on both occasions.
We need to mention here that the work of Loftus and her colleagues (e.g., Loftus, Miller, & Burns, 1978) has shown, in the behavioral domain, that such changes to stored memories are easily made, even when unintentional. This research is fairly well-represented in introductory psychology texts. However, the research on reconsolidation comes from neurobiological fields and therefore focuses on physiological dependent variables and the neural processes and structures involved (e.g., the amygdala). Another subtle but important difference is that reconsolidation research tends to address emotional aspects of memory rather than the episodic details emphasized in the work of Loftus. Therefore, what is important to note about memory reconsolidation is not only that it is a crucial part of memory encoding and storage processes that is being overlooked in curricula, but it is also an adaptive mechanism that has important clinical implications. Schiller et al. (2010) showed that extinction prior to reconsolidation prevents spontaneous recovery of an extinguished fear response. This knowledge, used in a clinical setting, could provide a way to erase, or at least diminish, negative emotional memories, specifically those that trigger posttraumatic stress disorder.
Memory reconsolidation is best suited for the discussion of the encoding processes in the memory chapter. Additionally, the Schiller et al. (2010) findings demonstrating the relationship between reconsolidation and fear depletion make this topic appropriate for treatment chapters. Finally, since reconsolidation studies utilize classical conditioning techniques, it can also be tied to the learning chapter.
Brain Plasticity
Neuronal plasticity includes the ability of the central nervous system to reorganize networks in the brain in response to the environment and recover after injury to the central nervous system (Johnston, 2003; Pascual-Leone et al., 2011). Brain plasticity occurs in two primary ways. Structural plasticity takes place during normal brain development when the brain undergoes physical changes in structure as a result of learning, memory, and developmental processes (Johansen-Berg, 2007). Functional plasticity shifts functions from one area of the brain to another following injury (Brain Plasticity, 2009). In contrast to structural plasticity, functional plasticity is an adaptive mechanism acting to maximize remaining functions in the brain and compensate for functions lost to injury (Anderson, Catroppa, Morse, Haritou, & Rosenfeld, 2005). Early knowledge of functional brain plasticity stems from rodent studies in which an area of the brain was lesioned and the surrounding cells began to undergo changes that allowed them to take over lost functions of the damaged area. Similar changes take place in human brains following injury and surgical procedures (Diamond, Krech, & Rosenzweig, 1964), perhaps the most drastic of which is hemispherectomy, a rare treatment for seizures in which an entire hemisphere is removed (Collins & Mullins, 2002; Duchowny, 2004). Studies have found that hemispherectomy is a highly successful procedure (Carreno et al., 2001; Jonas et al., 2004). For example, a recent study (Moosa et al., 2013) assessed the outcomes of 170 hemispherectomy patients, finding that 112 (66%) of the patients no longer experienced seizures and the remaining patients either experienced remission of seizures or showed a reduction in their frequency by over 90%. The success of this invasive procedure is due largely to the brain’s plasticity during the “critical period” of childhood. The critical period, a widely discussed concept of developmental psychology, is a period during which neural networks can be reshaped, resulting in structural and functional changes. As individuals age, the reshaping process becomes more difficult. Therefore, hemispherectomy is predominantly reserved for infants and toddlers because pediatric surgical candidates are able to couple the adaptiveness of functional plasticity with the structural plasticity that is most prevalent at this stage of life. Interestingly, memory and personality are shown to undergo normal development in these candidates following the procedure.
The developmental status of functions other than personality is largely dependent on which hemisphere is removed and which remains intact and fully functioning (Vining et al., 1997). Removal of the left hemisphere typically results in minor deficits in language comprehension. Interestingly, patients that underwent a left hemispherectomy had few differences in the grammatical characteristics of their speech compared to typically developing individuals and experienced close to normal grammatical development, making only minor errors (Curtis & Schaeffer, 2005; Kennison, 2014). However, the removal of the right hemisphere resulted in visuospatial deficits and increased difficulty completing complex tasks (Fournier, Calverley, Wagner, Poock, & Crossley, 2008). Some of the most significant differences between the removal of the right hemisphere versus the left can be seen in the patient’s ability to identify and make inferences about the emotions that another person is exhibiting. A study comparing two patients that underwent anatomical right and left hemispherectomies found that the removal of the right hemisphere resulted in severe deficits in the identification of negative emotional expressions. In contrast, the removal of the left hemisphere left interpersonal skills intact, with only minor difficulties in identifying negative emotional expressions, specifically anger and disgust (Fournier et al., 2008).
Brain plasticity is addressed in nearly every textbook, but is usually included as a sub-point to other topics. In some texts, it is briefly described in the neurobiological chapter under the discussion of changes in somatosensory inputs (Schacter, Gilger, Wegner, & Nock, 2014) or as an initial introduction to stem cells (Kalat, 2014). Although functional plasticity is best suited for the neurobiological chapter, it and structural plasticity should be discussed in detail to provide a platform for integration of the topic in the learning, language, intelligence, and development chapters, allowing for a well-rounded understanding of how it applies to the entire field of psychology.
Infant Moral Judgement
As previously mentioned, most introductory psychology texts discuss theory of mind (ToM) to some extent. ToM is the ability for one to understand that others have thoughts, feelings, and perceptions that may differ from one’s own (Premack & Woodruff, 1978) and is thought to develop at approximately four years of age. Thus, it is typically addressed in developmental chapters. Infants are thought to be unable to engage in ToM processes owing to the cognitive complexity required (Dunn, 1988). For this and other reasons, it has been long assumed that infants are unable to make judgements about the “goodness” or “badness” of the behaviors of others. However, recent research has discredited this assumption, finding that infants may make judgements regarding the behavior of agents, which appear to be moral judgements (Hamlin et al., 2007; Hamlin & Wynn, 2011).
In the prototypical experiment, infants are presented with a scenario that includes both a “helper” and a “hinderer” (characters that aid or prevent a protagonist from completing its goal). In one study (Hamlin, et al., 2007), infants 6 and 10 months of age observed the protagonist—a geometric shape with eyes –trying to reach the top of a hill. One condition showed the “helper” pushing the protagonist up the hill. In another, the “hinderer” pushed the protagonist down the hill it was attempting to climb. Infants were given a choice between the “helper” and “hinderer” leading to 26 of the 28 infants choosing the “helper” more that the “hinderer”, suggesting that these infants were basing their preferences on their evaluation of the behavior of the “helper” and “hinderer.” Replications and extensions of this study with infants of various ages (Hamlin et al., 2010; Hamlin & Wynn, 2011) have found similar results: Infants preferred the character that acted prosocially compared to the character that acted antisocially. The young age of the infant participants in these studies suggests not only that infants have an ability to make moral judgements, but that perhaps there is an innate capacity to distinguish “good” from “bad” behavior.
Further evidence suggesting moral judgements in infancy can be found in resource distribution research. Geraci and Surian (2011) examined whether infants can evaluate the subtler types of moral dilemmas involved with the inequity of resource distribution. They found that 16-month-old infants demonstrated a preference (measured through manual choice) for characters that distributed resources equally compared to those that distributed resources unequally. This indicates that even in subtle tasks, infants appear to be able to evaluate the fairness of the behavior of others.
The research on infant moral judgement is remarkable in that it identifies skills possessed by infants that have not only been unrealized, but the existence of which is denied by previous theories (e.g., Piagetian). Moreover, theory of mind can be a difficult concept for any layperson to understand, but can be particularly difficult to portray to a class of freshman students. These studies of infant moral judgement allow for a clear, real-world example of both theory of mind and empathy development in infancy.
Aside from Kohlberg’s stage theory (Kohlberg & Kramer, 1969), moral judgement and empathy are rarely mentioned in introductory texts, and virtually never with infants. This is surprising,
Owing to the number of fields and topics in psychology with which these concepts could be paired to aide in student’s understanding of more complex topics. For example, it could be useful in explaining the importance of mirror neurons in the neurobiological chapter, the implications of social behaviors and the development of social-emotional expressions in both the emotion and social chapters, and the development of theory of mind in the development chapter. Owing to the diversity of the topics with which infant moral judgement can be associated, and the implications this research has on theory of mind, a fundamental concept of child development, it warrants greater recognition within psychology texts than it is currently receiving.
Textbook Consolidation
Brief versions of introductory texts are a growing trend, with 6 of the 15 top-selling texts in the 2011–2012 academic year being brief versions containing 3–4 fewer chapters than their full-length companion texts (Griggs, 2014). Our review provides five topics that are currently under-represented in introductory textbooks, but the inclusion of new concepts increases the length of the textbook because outdated information is rarely removed. The question is, which old concepts should be removed to make room for new ones, and why? Identifying a list of specific topics to remove from future textbooks is beyond the scope of this paper. However, we can suggest one criterion that should be considered, namely, replicability, one of the defining features of scientific research. An example of a popular concept that has recently been put to the replication test is infant imitation of adult facial expressions, which is frequently introduced in texts through a discussion of Meltzoff and Moore’s (1983) research. A recent longitudinal study (Oostenbroek et al., 2016) failed to replicate those findings in infants between 1 and 9 weeks of age, reporting no evidence of the occurrence of these imitative behaviors. We do not mean to say that one failure to replicate should result in removing a concept from a textbook but we do propose that replicability is a necessary criterion for this selection process.
Conclusion
The goal of the presented work was to identify important emerging concepts in the fields of developmental and neurobiological psychology that are largely under-represented in introductory psychology textbooks. This was not intended to be a thorough search of every introductory text available, but a survey of widely used psychology texts that allowed us to gauge if and how these important concepts are taught in introductory psychology.
Psychology as a whole, and the individual areas within it, have continued to increase in both depth and breadth. However, the texts that introduce students to our broad and diverse field have not kept pace with these changes. Within both brief and full-length texts, the popular studies of Pavlov, Piaget, and other psychological pioneers are still given their earned fame and regard, but largely at the neglect of more recent studies that have expanded our knowledge of psychological processes. This is not to say that more recent studies should be included in place of such influential work as classical and operant conditioning, or developmental stage theories, but rather that these earlier works should provide a foundation upon which this new and exciting research has been built and from which it has evolved.
It is also important to remember that the topics addressed in this paper focus only on those concepts we feel are lacking in the neurobiological and developmental areas of psychology and further analysis of the available texts by specialists in the other areas of psychology is required in order to obtain a comprehensive list of topics throughout the field of psychology that are not being shown their due credit. It is not our intention to criticize the available psychology texts, but to inform authors and instructors of new, and in some cases, forgotten, research that is being overlooked. The field of psychology is growing and the texts need to reflect that growth by including the ongoing, new, and exciting research.
Footnotes
Acknowledgements
The impetus for this paper came from a request by Dr. Jeffrey Holmes to present at the 27th annual convention of the Association for Psychological Science. The inspiration for the topic came from a comment by Dr. Annette Taylor. Owing to the nature of the topic and the importance of it to the teaching psychology, Dr. Shelia Kennison of Oklahoma State University requested that we give the presentation at the 9th annual meeting of the Oklahoma Network for the Teaching of Psychology. We would like to thank both Dr. Holmes and Dr. Kennison for their interest in our topic and the support that they provided throughout the development of this project.
Declaration of Conflicting Interest
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.