
Abstract
Previous research has yielded mixed findings on the relationship between facial width-to-height ratio (fWHR), an androgen-dependent feature, and risk-taking propensity. We argue that mixed findings might result from overlooked variables. Given that risk-taking propensity might be ultimately linked to a search for mating opportunities, we analyze if reproductive strategies moderate the relationship between fWHR and risk-taking propensity. Our results, obtained from a sample of 434 male participants, show a positive association between fWHR and recreational and social risk-taking only for men who are more motivated to focus on mating effort over offspring survival. This finding aligns with research arguing that risk-taking may be a mating strategy since being social and recreational risk-prone might illustrate physical and psychological qualities and improve one's ability to attract mates. Our results support the notion that risk-taking might be a domain-specific construct. Overall, our research is in line with recent findings suggesting that the impact of testosterone exposure on risk-taking propensity is best understood when considering the role of contextual variables. Consequently, we add to previous research that studies related to risk-taking propensity should account for reproductive strategies.
Introduction
Testosterone exposure during the pubertal phase affects the body, the brain, and future behavior (Schultz et al., 2009). In particular, higher testosterone exposure is associated with facial width (Lefevre et al., 2013) and a greater risk-taking propensity among adolescent boys (Braams et al., 2015). Previous studies suggest that pubertal testosterone exposure might influence facial structures among men. For instance, experiments show that administering a low dose of testosterone to boys with delayed puberty accelerates the growth of facial structures (Verdonck et al., 1999). Likewise, research argues that testosterone exposure might influence facial structures (Roosenboom et al., 2018) and explain the variability in facial width-to-height (fWHR), suggesting a positive association between fWHR and pubertal testosterone exposure after controlling for age (Arnocky et al., 2018; Welker et al., 2016). Building on this rationale, we use the fWHR – an androgen-dependent feature - as an indicator of exposure to testosterone during the pubertal period.
Our research investigates the association between fWHR and risk-taking. Risk-taking propensity is an individual orientation toward avoiding (risk aversion) or engaging (risk-prone) situations with greater variability of consequences/ results (March & Shapira, 1992). During puberty, the rise in testosterone production in men (Duke et al., 2014) “shapes” brain areas underlying risky behavior (Peper & Dahl, 2013), which, in turn, might motivate adolescents to engage in more risky behaviors than older peers (Tymula et al., 2012). Though testosterone exposure during the pubertal period has been linked to risk-taking and fWHR, previous studies on the relationship between fWHR and risk-taking have yielded mixed findings. While some authors found an association between facial features and behaviors related to risk-taking (Arnocky et al., 2018; He et al., 2019; Kamiya et al., 2019), others found that there is an association between fWHR and risk-taking only when perceived status and experimentally manipulated status are low (Welker et al., 2015). Still, other research did not find an association between fWHR and behaviors related to risk-taking (Kosinski, 2017). These mixed findings may occur because overlooked variables might influence the relationship between pubertal androgen-dependent features, such as fWHR, and risk-taking.
Evolutionary scholars argue that risk-taking propensity might result from reproductive potential and parental investment (e.g., Baker & Baker, 2008; Buss & Schmitt, 2016; Saad, 2007). Trivers (1972) proposed a theoretical model based on an evolutionary lens, which has been used to explain mating behavior, the parental investment model (Vyncke, 2007). According to Trivers (1972), the sex with a lower parental investment demonstrates ways of rewarding its lower investment. For example, across the human species, while women gestate, breastfeed, and wait months until they can be pregnant again, men offer less vital parental investment. Because parental investment is greater in women than in men, women are more selective with whom they might mate.
In line with this, Price and Johnson (2011) argue that if males are the lesser parental-investing sex, they will engage in greater risk-taking to achieve higher status and better mating opportunities. For example, in hunter-gather societies, research shows a positive association between hunting skills, social status, and reproductive success (Price & Johnson, 2011). That might happen because hunting skills might provide food for the group, which translates to higher status and mating opportunities for the hunter (Saad, 2007). For instance, scholars argue that engaging in risky hunting might increase the reputation and attract mates (Bird et al., 2001; Hawkes & Bird, 2002). According to the human-mating literature, one possible explanation is that women have a mating preference for men who have a greater ability to obtain, keep, and share resources (Saad, 2007). Consequently, male hunting - a risky activity - may be a cue of mate quality, such as strength and agility (Bird et al., 2001), leading men to a higher social status and better chances of mating success. In an evolutionary sense, that probably happens because women's ancestors who mated to a higher-status men had better conditions to care for their offspring (Saad, 2007). Indeed, research argues that women prefer to mate with high-status and resourceful men (Buss, 1994; Geary et al., 2004; Rommel, 2002).
While hunting skills are seldom a signal of status and resources in today's societies, one's risk-taking propensity may lead men to be more successful in obtaining status resources and improving mating success. Some researchers argue that risk-taking might be a helpful signal in the mating market (Hawkes & Bird, 2002; Kelly & Dunbar, 2001; Wilke et al., 2006). Still, others say that mating motives might influence risk-taking propensity in men (Baker & Baker, 2008). Indeed, some studies report support for that evolutionary assumption. For example, Wilson and Daly (2004) show that when men view attractive women's faces, they give greater value to immediately available resources over the ones that would be available only in the future. In line with this, research shows that men are more prone to take risks in the presence of a woman (Pawlowski et al., 2008).
Considering that risk-taking propensity might be ultimately linked to status-seeking and a search for mating opportunities under an evolutionary lens, reproductive strategies might moderate the relationship between pubertal androgen-dependent features, such as fWHR, and risk-taking propensity. Reproductive strategies are personal resources or characteristics (psychosocial traits) related to enhancing the chances of mating or infant survival (Figueredo et al., 2006). According to evolutionary scholars, reproductive strategies vary from mating to parental investment. When there is a tendency for mating investment, the individual is more prone to short-term relationships. In contrast, individuals focused on parental investment tend to be selective regarding mating and focus on long-term relationships (Figueredo et al., 2005; Rushton, 1985).
Given that individuals more motivated to mating investment seek short-term and non-committed relationships (Figueredo et al., 2006), one could argue that they would be more prone to take risks that might improve their chances of mating. Studies show that women value risk propensity as an attractive characteristic when considering short-term relationships (e.g., Apalkova et al., 2018; Kelly & Dunbar, 2001; Sylwester & Pawlowski, 2011). For instance, research shows that women rate bravery (a risk-taking male personality trait) as a great influential attractiveness for short-term sexual partners (Kelly & Dunbar, 2001). Conversely, individuals prone to parental investment focus on long-term relationships and offspring survival. Thus, they might be less inclined to take risks fearing the consequences for their relationship and offspring. For instance, research shows that single men score higher on risk-taking propensity than those in a committed relationship or married (Wilke et al., 2014). Consequently, if risk-taking behavior is a mating strategy, some risk domains might illustrate characteristics that women desire in men, while others may have the opposite effect. For instance, research shows that women consider ethical and health risk domains unattractive (Wilke et al., 2006). In addition, financial risk is perceived as unattractive (when related to gambling) or neutral (when associated with investment) (Wilke et al., 2006). Therefore, such risk-taking propensities might not function as a way to signal male quality. Interestingly, women report recreational and social risk-taking as attractive (Wilke et al., 2006). Therefore, we adopt a risk-specific approach following previous research (e.g., Blais & Weber, 2006; Hanoch et al., 2006; Weber et al., 2002; Wilke et al., 2006). Under this approach, the risk-taking propensity depends on its domain, i.e., a person can show a tendency to a particular risk domain and an aversion to another.
We follow Blais and Weber (2006), and we analyze five risk domains: financial (proclivity of betting or investing money in risky choices); recreational (propensity to engage in extreme sports); social (tendency to risk one's social status, whether related to friendship, family, or job); ethical (inclination to engage in situations or activities considered illegal or against social norms); and health risk domains (tendency to engage in situations that put one's health at risk). This paper investigates the moderating influence of reproductive strategies on the association between pubertal testosterone exposure (as measured via the fWHR) and risk-taking propensity (across different risk domains).
With this research, we provide three contributions to the literature. First, our result shows that fWHR is positively associated with recreational and social risk-taking propensity, but only for men prone to short-term mating. Consequently, we highlight the importance of including other relevant moderators to understand risk-taking propensity. Second, we found that the association between fWHR and risk-taking propensity varies according to the risk-taking domain. Third, we bridge two literatures related to risk-taking: those who research the relationship between fWHR and risk-taking and those who accept risk-taking propensity as a way of getting advantages in the mating market.
Literature Review
Testosterone and Facial Width-to-Height Ratio (fWHR)
Testosterone is a gonadal steroid hormone that affects the nervous system in two ways, activational and organizational. The activational effect is transient and depends on the current level of the hormone to influence behavior in specific physiological or social contexts during adulthood (Sisk et al., 2013). The organizational effect happens in developmental periods such as late prenatal/ early postnatal life and puberty, and its effect on the nervous system persists beyond exposure to the hormone (Sisk et al., 2013). Consequently, it is often irreversible in programming sensitivity and responsiveness of the nervous system to hormones over the entire life span. Therefore, the hormonal activation during later periods, such as adulthood, depends on the organizational effects of earlier exposure to hormones on the nervous system, i.e., the long-term organizational effects (Archer, 2006; Sisk et al., 2013).
Saliva samples indicate testosterone's activational effects (Apicella et al., 2008). Some androgen-dependent features that indicate organizational influences of testosterone exposure are digit (i.e., fingers’ length) and facial ratios. The digit ratio is used as an indicator of exposure to testosterone during the prenatal phase because previous research shows that exposure to testosterone during prenatal life impacts the fingers’ length (e.g., Lutchmaya et al., 2004; Manning et al., 1998). The fWHR indicates exposure to testosterone during the pubertal phase. That happens because previous research argues that facial growth is associated with testosterone levels during puberty (e.g., Verdonck et al., 1999), such as the higher the exposure to testosterone during puberty, the wider the face (Lefevre et al., 2013). In line with this, previous research shows that pubertal testosterone might explain the variability in fWHR (Welker et al., 2016).
Research shows no correlation between digit and facial ratios (e.g., Apicella et al., 2008). Consequently, different indicators of testosterone exposure might signal distinct effects of testosterone exposure. Most previous research on the organizational effects has concentrated on the correlation between digit ratio and risk-taking (e.g., Chicaiza-Becerra & García-Molina, 2017; Lima de Miranda et al., 2018; Stenstrom et al., 2011). We focus on the association between fWHR and risk-taking propensity.
Studies have linked men's androgen-dependent features, such as the fWHR, to various behavioral characteristics (e.g. Arnocky et al., 2018). However, the relationship between fWHR and risk-taking propensity remains unclear. Previous research investigating the link between fWHR and risk-taking propensity has focused mainly on a specific risk such as firm risk (e.g., Kamiya et al., 2019), ethical-related risk (e.g., Arnocky et al., 2018), or health-related risk-taking (Kosinski, 2017). However, there are different types of risk: financial, recreational, social, ethical, and health (Blais & Weber, 2006; Weber et al., 2002).
Puberal Testosterone and Risk-Taking
The literature investigating the association between facial features and risk-taking is mixed. While some authors found an association between facial features and risk-taking, others did not. For example, after analyzing a sample of 1,200 Chief Executive Officers (CEOs) and correlating it to the firm's risk, authors found that firms that have CEO with more masculine faces (i.e., bigger fWHR) are more acquisitive and have higher financial leverage (Kamiya et al., 2019). Similarly, research suggests that banks led by CEOs with more masculine facial features have more volatile stock returns and higher levels of idiosyncratic risk (He et al., 2019). Likewise, other research found that fWHR predicts intended infidelity, an example of ethical risk (Arnocky et al., 2018). Also, measuring fWHR, researchers found a positive association between fWHR and risk-taking only when perceived status and experimentally manipulated status are low (Welker et al., 2015). However, other research has failed to replicate the positive association between fWHR and antisocial behaviors or interest in drug use (e.g., Kosinski, 2017). Notably, antisocial behaviors and interest in drugs overlap with some risk domains developed by Blais and Weber (2006).
These contradicting findings may occur because previous studies are related to different risk domains and because there may be moderators overlooked by previous research, such as status and reproductive strategies. Indeed, studies have reported that low-status individuals are more prone to health risks, such as risky sexual behaviors (e.g., Capaldi et al., 2002) and smoking behaviors (Finkelstein et al., 2006). Other research argues that men are more likely to engage in risk-taking when exposed to images of attractive women (Baker & Baker, 2008). Moreover, risk-taking might be associated with what degree men are perceived as attractive by women (Apalkova et al., 2018). Consequently, risk-taking might be related to one's effort dedicated to mating opportunities or offspring survival (Buss, 2003; Ellis et al., 2012; Miller, 2001). In this sense, one could argue that reproductive strategies might influence the relationship between fWHR and risk-taking.
Testosterone is a hormone thought to modulate facial dominance/masculinity (e.g., Penton-Voak & Chen, 2004) during specific periods of human development (Arnocky et al., 2018). During puberty, testosterone exposure influences the growth of facial structures (Verdonck et al., 1999). Interestingly, men's bodily dominance is related to status-seeking behaviors (Kordsmeyer et al., 2019); and according to evolutionary theories, risk-taking may have evolved as a behavioral strategy for mating (Miller, 2001; Saad, 2007). Therefore, in an evolutionary sense, men may take risks to acquire social status and show reproductive advantages. For example, previous research showed that male skateboarders engage in higher risks when a nubile woman is around (Ronay & von Hippel, 2010). Hence, risk-taking can advertise male qualities, and it might signal important cues in human mate choice (Wilke et al., 2006). Given that people differ in their qualities and that women view some risk domains as unattractive (Wilke et al., 2006), risk-taking can become a strategy that depends on particular social conditions (Ermer et al., 2008).
Of note, while the literature reviewed above provides evidence that high pubertal testosterone exposure is appropriate for predicting risk-taking propensity in men, the evidence in women is less clear. Testosterone plays a role in mating behavior in men, such that pubertal testosterone exposure is related to sexual desire (Arnocky et al., 2018) in men. In women, sexual desire has been shown to be associated with fWHR in some studies (e.g., Arnocky et al., 2018) but not in others (e.g., Zhang et al., 2018). Furthermore, while there is evidence that fWHR is associated with testosterone levels in men (Lefevre et al., 2013), we have not seen such evidence in women. Consequently, the relationship between pubertal testosterone and risk-taking might be more appropriately investigated in men.
Reproductive Strategies
Scholars rooted in evolutionary psychology describe reproductive strategies based on a tradeoff between quantity and quality of offspring, balancing mating and parenting effort. Some evolutionary scholars posit that individuals’ differences depend on environmental affordances (Volk, 2021; Heath & Hadley, 1998) and are a result of psychosocial traits that cluster together in a continuum that goes from “low-K” to “high-K” (Figueredo et al., 2006; Giosan, 2006) (Rushton, 1985). The “K” is derived from standard ecological algebra and means the carrying capacity of a local environment (Hui, 2006; Pianka, 1970), and “K” individuals are those that have relatively small numbers of offspring. Though human beings are “K” animals, the level of being a “K strategist” can vary among individuals (Figueredo et al., 2005, 2006; Rushton, 1985).
Researchers argue that the more high-K a person is, the more a person seeks long-term relationships, the greater the parental investment and the obedience to social rules (Figueredo et al., 2006). Other differences between a low-K and a high-K individual are the qualities and traits that enhance reproductive potential and the capacity to provide better conditions for oneself and the prole, which are related to environmental affordance. In modern society, one can recognize a high-K individual by the following characteristics: maintenance or improvement of health for self, descendants, and kin; better access to healthcare, educational and career opportunities for descendants; access to social capital and extended family (receiving help from others); living or working in safe places and having better living conditions (Giosan, 2006).
Evolutionary Roots of Risk-Taking
Some behavioral traits, such as risk-taking propensity, might be connected with evolutionary theories. For example, previous studies argue that risk-taking propensity might function as a mating strategy that allows men to out-compete other males by increasing status and generating advantages in mating opportunities (Ellis et al., 2012). Given that women view social and recreational risk-taking as attractive (Wilke et al., 2006), and considering that men also believe that social and recreational risks are perceived as attractive by women, we expect that men will be inclined to take such risks for mating purposes.
Regarding recreational risk, physical traits such as strength, fitness, and athleticism indicate survival value and phenotypic quality and thus might be attractive to the opposite sex (Wilke et al., 2006). Given that engaging in risky activities can show these traits (Sylwester & Pawlowski, 2011), one could argue that recreational risk might be a way of gaining advantages in the mating market. However, high-K individuals might be more worried about their children's future than low-K people (Figueredo et al., 2006). Hence, one can assume that high-K men might refrain from engaging in recreational risk, fearing any physical injury that might negatively impact their future and, consequently, their children's future.
Regarding social risk, previous research argues that high-K individuals are more predisposed to obey social rules and are more worried about their children's future than low-K people (e.g., Figueredo et al., 2006; Giosan, 2006). Moreover, the social risk is related to behaviors that can result in social penalization or a decreased reputation (Sylwester & Pawlowski, 2011). Consequently, one could expect that high-K individuals would not be prone to social risk, fearing losing social status. Conversely, low-K individuals may have lower social status and may be more motivated to engage in risk-taking situations to gain and maintain status. Therefore, we test if:
H1: The association between fWHR and social risk is positive for low-K individuals.
H2: The association between fWHR and recreational risk is positive for low-K individuals.
Ethical risk-taking propensity denotes a likelihood of having an affair with a married person or leaving the kids alone at home to run some errands (Blais & Weber, 2006). Consequently, women may not see it as a positive characteristic, and therefore, it might not function as a reproductive strategy for men. Indeed, research shows that women report ethical risk as an unattractive cue (e.g., Wilke et al., 2006).
Men's inclination to invest resources in the family (e. g., partner or offspring) is a sensitive point for women (Buss & Schmitt, 1993; Buss, 2003). Hence, it might be an important cue for mating purposes. Financial risk-taking propensity indicates a tendency to betting money or gamble (Blais & Weber, 2006; Weber et al., 2002), which might not benefit the partner or the offspring, so it may not be a good reproductive strategy. Indeed, research shows that women rate gambling as an unattractive characteristic in men and investing as a neutral cue (e.g., Wilke et al., 2006). In line with this, Farthing (2005) reports that women see financial risk-taking as an unattractive characteristic in men.
Similarly, health risk-taking propensity might not function as a positive trait. It implies attitudes that negatively impact health and well-being, such as drinking heavily or having unprotected sex (Blais & Weber, 2006). Consequently, being prone to this risk might not function as a reproductive strategy. For example, Farthing (2005) shows that drugs or alcohol consumption - health-related risk-taking - is a disapproved characteristic of mates. Likewise, Wilke et al. (2006) show that women see health-risk-taking propensities unattractive. Moreover, research has failed to replicate a positive association between fWHR and interest in drug use (a health-related risk) (Kosinski, 2017). Consequently, health risk-taking propensity might not function as a reproductive strategy.
In conclusion, we expect that reproductive strategies moderate the association between pubertal testosterone exposure (as measured via the fWHR) and risk-taking propensity only in risk domains that generate an advantage in the mating market (i.e., social and recreational). Research argues that financial, ethical, and health risks might be unattractive characteristics for mating purposes (e.g., Wilke et al., 2006). Consequently, we do not make any hypotheses for these three risk domains.
Method
The fWHR is the ratio between the width (distance between the zygomatic bones, determinants of the lateral end of the face) and face height (distance between the upper lip and the midpoint of the eyebrows) (Carré & McCormick, 2008; Lefevre et al., 2013). Given the link between fWHR and some behaviors related to risk-taking (e.g. Arnocky et al., 2018), we test if the fWHR is associated with social and recreational risk-taking propensity and whether reproductive strategies moderate this association.
We did not pre-register the hypotheses, but we obtained approval from the Brazilian Research Ethics Council before collecting the data. Most research on the association between fWHR and risk-taking propensity, or risk-taking related behaviors, is from WEIRD societies (Western, Educated, Industrialized, Rich, and Democratic societies) (e.g., He et al., 2019; Kamiya et al., 2019). However, according to the American Economic Class System (Snider & Kerr, 2021), our sample comprised males from a poor region of Northeastern Brazil, in which 81% of the respondents correspond to people from the low-income class.
We followed Bentler and Chou (1987) and Hair Jr. et al. (2009) to calculate the sample size. They suggest that the sample should be between 5 and 10 respondents per the questionnaire scales’ assertation. The questionnaire has 56 assertations (30 related to the risk-taking propensity scale and 26 to the Reproductive Strategy); therefore, the minimum sample would be 280 participants. Four hundred and eighty-three men participated in the initial sample. We removed 49 participants because it was not possible to accurately calculate their fWHR ratios with the photos taken (n = 7) and because participants had not fully responded to the questionnaire (n = 42). Therefore, the final sample comprised 434 participants (age M = 20.8 [18;35]; SD = 3.07). We did not recruit any participants under 18.
We use age as a covariate in the analyses for two reasons. First, when controlling for age, there is a positive association between fWHR and exposure to pubertal testosterone (Welker et al., 2016). Second, circulating testosterone levels change with age (Feldman et al., 2002; Vermeulen et al., 1999). Given that circulating testosterone is shown to influence risk-taking propensities (Herbert, 2018), we opted to collect data with participants of a similar age group to avoid the indirect effect of circulating testosterone (due to age) in our results. In addition to age, we included sexual orientation as a covariate because the signaling of mating qualities to potential mates might differ according to men's sexual orientation (Kuhnreich et al., 2018). Consequently, it might influence how risk-taking propensities might be used or perceived in the mating market for non-heterosexual men.
Researchers took five frontal facial pictures of each participant using a Canon EF-S 18-S5III Rebel T100 camera. From those five pictures, we chose the best one according to the research pattern: (i) photos in a neutral position, without smiling or any other muscle facial movement; (ii) participants without glasses, headband, beard, or tilting the head, and against a neutral backdrop; (iii) using a 55 mm focal length and under standardized lighting conditions; iv) constant distance between the participant and the tripod (1.8 meters). The chosen pictures were sent to a computer to calculate the fWHR ratio. Following Wilson et al. (2014), we calculate the fWHR ratio by dividing the width (horizontal facial distance from the left facial edge to the right facial edge) and the upper-face height (the distance from the center of the upper lip to the mid-point of the eyebrows). The fWHR was measured using the fWHR calculator (Ties de Kok, 2018), an open-source software.
Scales
We used the domain-specific risk attitude scale described in Blais and Weber (2006) to measure risk-taking propensity. Each of the five domains contained seven-point Likert-type items (1 to 7) assessing one's propensity to engage in risky situations. The items include issues related to: investing a percentage of the annual income in a new business venture (financial risk, α = .613); practicing bungee jumping (recreational risk, α = .839); having unprotected sex (health risk, α = .693); choosing an enjoyable career over a secure one (social risk, α = .600); having a relationship with a married person (ethical risk, α = .641).
We use the High-K scale (Giosan, 2006) to measure reproductive strategies (K-Factor). According to the K-Factor continuum, reproductive strategies can vary from “low-K” to “high-K” (Figueredo et al., 2006) (Rushton, 1985). Individuals prone to the “low-K” end tend to focus on mating effort over offspring survival. Therefore, they might be more inclined to have numerous mates, given that mating effort is devoted to producing new organisms. In contrast, the survival effort is related to infant survival and parental effort. When individuals are more “high-K”, they tend to be selective regarding mating, focus on long-term relationships, have a higher parental investment (Figueredo et al., 2005), and have better social conditions (Giosan, 2006).
The K-Factor (α = 0.743) is a five-point Likert item (1 to 5). The 26 items include issues related to health benefits for the family and oneself; self-perception of being attractive; opportunities for promotion and increasing income; physical shape; how easy it is to find and go on a new date. The domain-specific risk and the K-Factor scales are in Appendices A1 and A2.
Results
The correlations between the five risk-domains range from.121 (r(434); p = .011) for recreational and ethical risks and.469 (r(434); p < 0.001) for ethical and health risks (see Table 1). Also, importantly, we found no significant Pearson's correlation between fWHR and the five types of risk propensities: financial risk and fWHR (r(434) = .048; p = .322); recreational risk (r(434) = .059; p = .221); social risk (r(434) = .065; p = .180); ethical risk (r(434) = −.002; p = .965); health risk (r(434) = .085; p = .077). Likewise, we found no significant Pearson's correlation between general risk (a composite measure including all five domains of risk) and fWHR (r(434) = .084; p = .081).
Reliability of Risk and K-Factor Scales (α), Correlations, and Descriptive Statistics.

*** The exact p-values are in Appendix A3.
**Significance at the 0.01 level (2-tailed).
*Significance at the 0.05 level (2-tailed).
We tested if K-Factor moderated the association between fWHR and risk-taking across the five risk domains (financial, recreational, social, health, and ethical). We used IBM SPSS Statistics 22 PROCESS macro model 1 (Hayes, 2018, version 4.0), continuous predictors were mean-centered; n = 434), and age was a covariate variable. There was a significant moderation when predicting recreational risk. Specifically, bigger fWHR (an androgen-dependent feature used as an indicator of high testosterone exposure during puberty) was significantly associated with greater recreational risk among lower-K individuals (−1SD, b = −.449, t = 2.861, p =. 004). There was no significant moderation among participants scoring average K-Factor (MSD, b = .000, t = 1.339, p = .181) or high K-Factor ( + 1SD, b = .449, t = −.932, p = .352) (see Figure 1). Participants scoring average K-Factor had the K-Factor equal to the mean K-Factor score, and participants scoring high K-Factor had the K-Factor greater than the mean K-Factor.
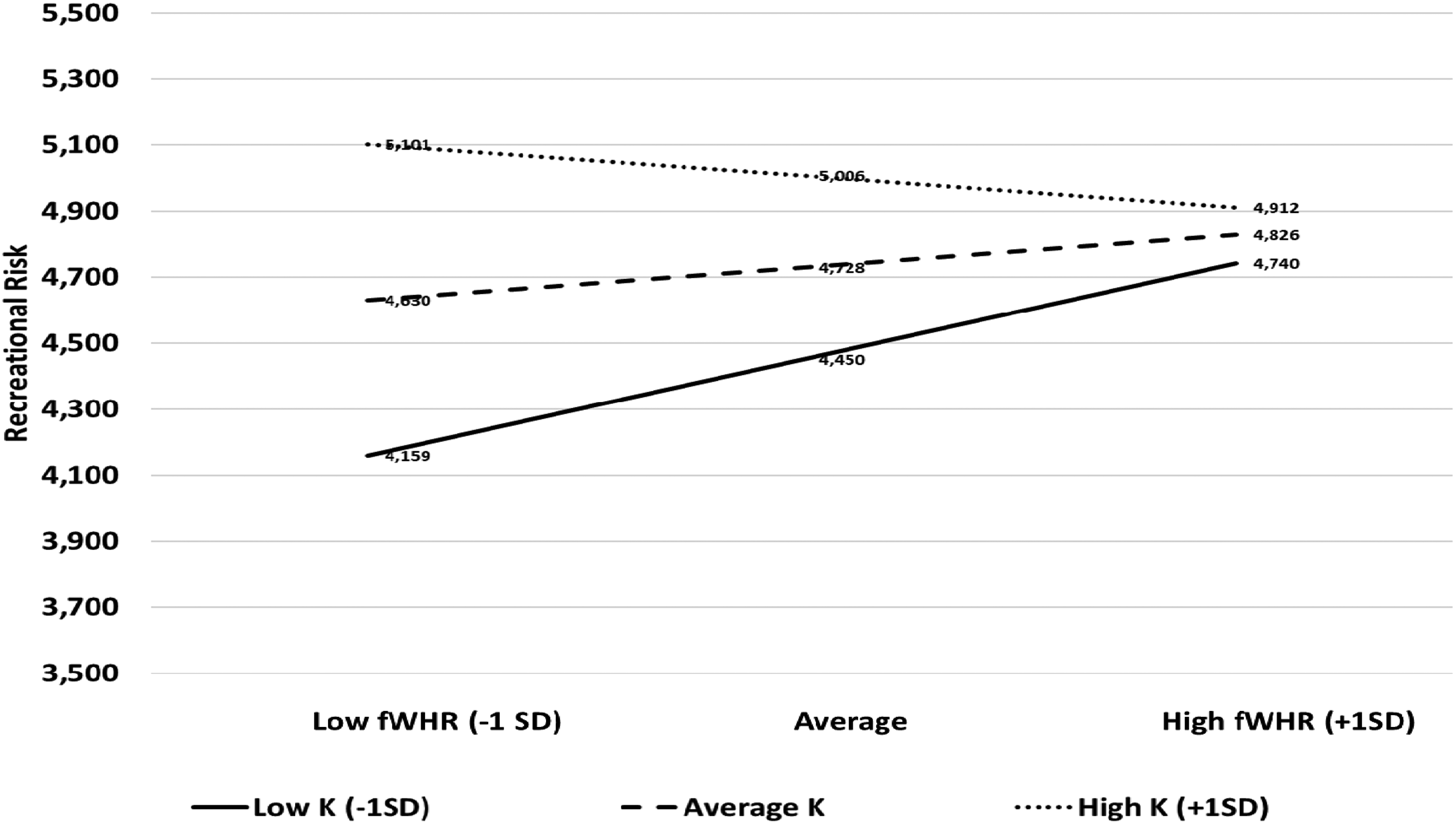
The relationship between fWHR and recreational risk-taking propensity according to reproductive strategies.
The results did not change when we investigated the moderation of the K-Factor without age as a covariate. There was also a significant moderation among lower-k individuals (−1SD, b = .−449, t = 2.850, p = .005). Among participants scoring average K-Factor (MSD, b = .000, t = 1.268, p = .206), or higher-K Factor ( + 1SD, b = .449, t = −1.044, p = .297), there was no moderation. Similarly, when we included sexual orientation as a covariate, the results did not change. There was a significant moderation for lower-K individuals (−1SD, b = -.449, t = 2.799, p = .005). Among participants scoring average K-Factor (MSD, b = .000, t = 1.264, p = .207) or higher-K Factor ( + 1SD, b = .449, t = −.999, p = .318), there was no moderation.
There was a significant moderation of the K-Factor when predicting social risk. Specifically, bigger fWHR was significantly associated with greater social risk among lower-K individuals (−1SD, b = −.449, t = 2.316, p = .021). Among participants scoring average K-Factor (MSD, b = .000, t = 1.088, p = .277) or high K-Factor ( + 1SD, b = .449, t = −.748, p = .455), there was no significant moderation (see Figure 2).

The relationship between fWHR and social risk-taking propensity according to reproductive strategies.
The significance of the moderation did not change when we analyzed the data without age as a covariate. There was a significant moderation among lower-k individuals (−1SD, b = −.449, t = 2.350; p = .019). Among participants scoring average K-Factor (MSD, b = .000, t = 1.369; p = .172) or high-K Factor ( + 1SD, b = .449, t = -.391, p = .696), there was no significant moderation for social risk-taking propensity. Furthermore, the significant results did not change when we included sexual orientation as a covariate. There was a significant moderation for lower-K individuals (−1SD, b = .-.449, t = 2.521, p = .012). Among participants scoring average K-Factor (MSD, b = .000, t = 1.673, p = .095) or high-K Factor ( + 1SD, b = -.449, t = −.122, p = .903) there was no significant moderation.
Regarding financial (b = −.888, t = −1.482, p = .139) and health (b = −.747, t = −1.063, p = .288) risks, there was no significant moderation of the K-Factor. When we analyzed financial risk without age as a covariate (b = −.924, t = −1.549, p = .122), or including sexual orientation as a covariate (b = −.887, t = −1.519, p = .130), K-Factor did not moderate. Regarding health risk, there was no significant moderation when age is not included as a covariate (b = −.764, t = −1.093, p = .275), or when sexual orientation is included as a covariate (b = −.728, t = −1.042, p = .298). For ethical risk, there was no significant moderation of low K-Factor (-SD, b = −.449, t = 1.358, p = .175) or average K-Factor (MSD, b = .000, t = −.015, p = .988) or high-K Factor ( + SD, b = .449, t = −1.385, p = .167), when considering age as covariate. Similarly, there was no significant moderation of low K-Factor (−SD, b = −.449, t = 1.361, p = .174), or average K-Factor (MSD, b = .000, t = -.008, p = .994), or high K-Factor ( + SD, b = .449, t = −1.390, p = .165), without age as a covariate. Likewise, the results did not change when we included sexual orientation as a covariate (−SD, b =- .449, t = 1.469, p = .142); (MSD, b = .000, t = .168, p = .866); ( + SD, b = .449, t = −1.246, p = .213).
Lastly, regarding general risk (a composite measure including all five domains of risk), our results show that larger fWHR was significantly associated with greater general-risk propensity among lower-K individuals (−1SD, b = −.449, t = 3.420, p = .001). There was no significant association for the average-K individuals (MSD, b = .000, t = 1.791, p = .074). There was no moderation for individuals scoring high on the K-Factor ( + 1SD, b = .449, t = −.838, p = .402; see Figure 3).
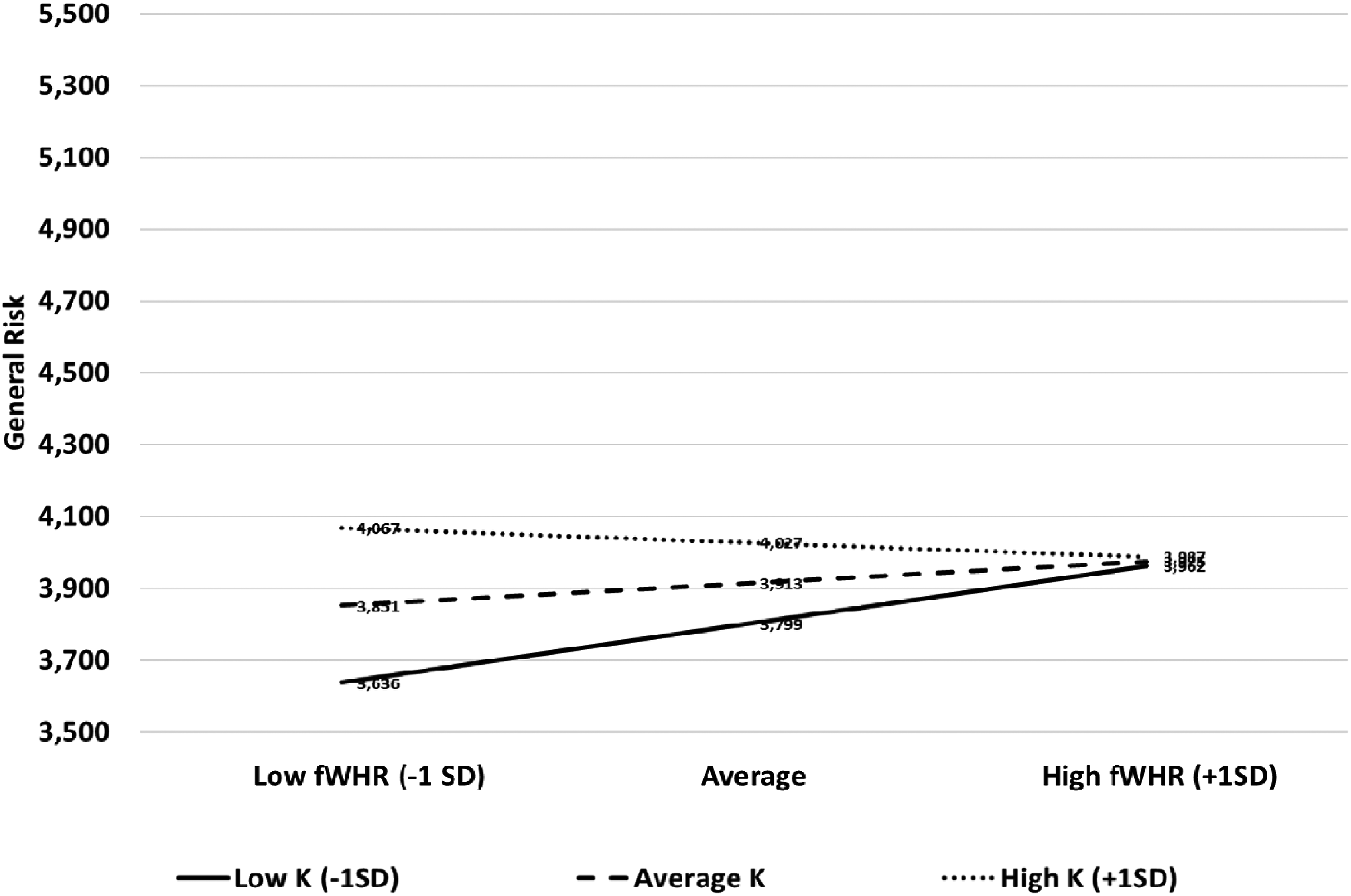
The relationship between fWHR and general risk-taking propensity according to reproductive strategies.
For low-K individuals, the results remained largely unchanged (i.e., there is an association between fWHR and general risk) after removing age as a covariate (b = −.449, t = 3.425, p = .001) or including sexual orientation as a covariate (b = −.449, t = 3.449, p = .001). There was no significant moderation for individuals scoring average-K when we took off age as a covariate (MSD, b = .000, t = 1.807, p = .071) or when we included sexual orientation as a covariate (MSD, b = .000, t = 1.920, p = .056). Finally, there was no significant moderation for high-K individuals when we did not consider age as a covariate (b = .449, t = −.844, p= .399) or when we included sexual orientation as a covariate (b = .449, t = −.705, p = .481). Overall, these analyses show that the results for general risk are largely similar if we include or exclude the covariates. Please refer to Appendix A4 for the full regression equation and Appendix A5 for the non-standardized coefficients.
Discussion
Our results provide three contributions to the literature. First, it shows that an indicator of exposure to pubertal testosterone (i.e., fWHR) is positively associated with recreational and social risk-taking propensity, but only for men motivated to engage in short-term and non-committed relationships (i.e., lower-K men). This finding confirms H1 and H2 and is in line with previous studies that have argued that “low-K” individuals disregard social rules and have extensive risk-taking propensity (Bogaert & Rushton, 1989; Figueredo et al., 2006). Furthermore, by showing that an individual's reproductive strategies influence the relationship between fWHR and risk-taking, we further support the notion that testosterone exposure has a meaningful impact on future behavior. In particular, we demonstrate that fWHR is associated with risk-taking propensity only for individuals following specific reproductive strategies, that is, those more inclined to short-term mating (low-K individuals) over survival effort (high-K individuals).
Second, we found that the association between pubertal testosterone exposure (i.e., fWHR) and risk-taking propensity is not uniform, and it depends on the risk-taking domain. For example, there is a positive association between fWHR and two risk domains, social and recreational, but only for low-K individuals. There was no association between fWHR and ethical, financial, or health risk propensities for any level of the K-Factor. Additionally, we found a positive association between fWHR and general risk (a composite measure including all five risk domains) for low-K individuals. Given that the results are not steady across all risk domains, our research is in line with past research suggesting novel insights when considering a domain-specific perspective (e.g., Blais & Weber, 2006).
Third, we analyze the relationship between fWHR and risk-taking propensity under evolutionary lenses, specifically as a reproductive strategy. With that, we bridge two literatures related to risk-taking. That is, those who research the relationship between fWHR and risk-taking (e.g., Ahmed et al., 2019; Kamiya et al., 2019); and those who study risk-taking propensity under an evolutionary lens or as a sign of advantage in the mating market (e.g., Ellis et al., 2012; Miller, 2001). By demonstrating that reproductive strategies moderate the association between fWHR and risk-taking propensities, we show that testosterone exposure motivates risk-taking propensity for those who focus on short-term mating (lower-K individuals).
Previous research overlooked reproductive strategies (e.g., Kamiya et al., 2019), which could be one possible explanation for the previous conflicting results. Therefore, future research on the relationship between fWHR and risk-taking propensity should consider reproductive strategies.
As we point out in the Literature Review, digit ratios, fWHR, and other measures of testosterone (e.g., saliva samples) measure different constructs. While fWHR measures puberal exposure, digit ratios measure prenatal exposure, and saliva samples measure current testosterone levels (i.e., circulating measure of testosterone). Among the listed measures, fWHR is the least intrusive to measure and more easily accessible by others, making the findings more readily applicable.
Supplemental Material
sj-docx-1-evp-10.1177_14747049221096758 - Supplemental material for Individual's Reproductive Strategies Moderates the Association Between Facial Width-to-Height and Risk-Taking Propensity
Supplemental material, sj-docx-1-evp-10.1177_14747049221096758 for Individual's Reproductive Strategies Moderates the Association Between Facial Width-to-Height and Risk-Taking Propensity by Fabiane de Barros Figueirêdo Cavalcante, Marcelo Vinhal Nepomuceno, Danielle Miranda de Oliveira Arruda Gomes and Samuel Façanha Câmara in Evolutionary Psychology
Footnotes
Acknowledgments
The study was supported with funding given to Fabiane de Barros Figueirêdo Cavalcante by the Fundação Cearense de Apoio ao Desenvolvimento Científico e Tecnológico (Funcap – Ceará, Brazil), from March 2016 to March 2020. The authors would like to thank Ana Beatriz Bernardes, Igor Cordeiro, Elano Rolim, Mark Rodrigues, Laryssa Melo, and José Iran Filho for assisting with the data collection.
Data Availability Statement
The data that support the findings of this study are available on request from the corresponding author. The data are not publicly available due to privacy or ethical restrictions.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the Fundação Cearense de Apoio ao Desenvolvimento Científico e Tecnológico,
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.