
Abstract
The fields of developmental and comparative psychology both seek to illuminate the roots of adult cognitive systems. Developmental studies target the emergence of adult cognitive systems over ontogenetic time, whereas comparative studies investigate the origins of human cognition in our evolutionary history. Despite the long tradition of research in both of these areas, little work has examined the intersection of the two: the study of cognitive development in a comparative perspective. In the current article, we review recent work using this comparative developmental approach to study non-human primate cognition. We argue that comparative data on the pace and pattern of cognitive development across species can address major theoretical questions in both psychology and biology. In particular, such integrative research will allow stronger biological inferences about the function of developmental change, and will be critical in addressing how humans come to acquire species-unique cognitive abilities.
Introduction
Human behavior is strikingly different from that of other animals. We speak language, routinely cooperate with others to reach complex shared goals, engage in abstract reasoning and scientific inquiry, and learn a rich set of sophisticated cultural behaviors from others. Why do we possess such abilities while other species do not? How do we acquire these abilities? One approach in answering these core questions has been to examine the roots of human cognitive systems. The field of developmental psychology investigates the origins of adult cognition through the study of infants and children, assessing how cognitive capacities emerge across ontogeny. This research has revealed that human cognitive development is shaped by capacities thought to be derived in our species, such as language and shared intentionality (Hermer-Vazquez, Moffet, and Munkholm, 2001; Pyers, Shusterman, Senghas, Spelke, and Emmoret, 2010; Tomasello, 1999; Tomasello, Carpenter, Call, Behne, and Moll, 2005). The field of comparative psychology, in contrast, investigates the evolutionary origins of human cognition via research with humans' closest living relatives, non-human primates, as well as other species of evolutionary interest. This field has shown that many primate species exhibit an array of complex abilities previously thought to be unique to humans, including reasoning about others' perceptions and knowledge, episodic memory and planning, and the ability to use and create complex tools (Call, 2008; Flombaum and Santos, 2005; Humle and Matsuzawa, 2002; Martin-Ordas, Haun, Colmenares, and Call, 2009; Mulcahy and Call, 2006; Seed and Byrne, 2010)
Both developmental and comparative approaches have therefore been critical in generating and testing hypotheses regarding the origins of human-unique cognition. Yet little attention has been paid to the intersection of developmental and comparative inquiry: studies of cognitive development across species. Though several researchers have highlighted the importance of a comparative developmental perspective (Gomez, 2005; Langer, 2000; Lickliter, 2000; Matsuzawa, 2007; Matsuzawa, Tomonaga, and Tanaka, 2006), this integrative comparative-developmental approach has not been widely applied in empirical work. Therefore, our current aim is to highlight the types of advances that comparative developmental research can provide. In particular, we aim to show that comparative developmental studies can contribute important insights into the origins of human cognition by illuminating the pace and pattern of cognitive development across species. We begin by examining theoretical models of how cognition can change across species via changes in the pace and pattern of development. We then outline how studies of comparative cognitive development can bear on current open questions in both psychology and biology. Subsequently, we review empirical work incorporating comparative developmental data in three cognitive domains: 1) reasoning about objects, 2) cognitive skills used in foraging contexts, and 3) social cognition. Finally, we conclude by identifying unsolved questions that can be addressed using a comparative developmental framework. Overall, we argue that a comparative developmental approach is necessary to understand the origins of human cognition, including features of development that are unique to our species.
Changes in the pace and pattern of development
We propose that evolutionary changes in development can alter cognitive abilities across species via two main mechanisms: differences in the pace or timing of cognitive development, and differences in the pattern or structure of cognitive development. In terms of change in pace, much research in developmental psychology is concerned with the timing of skill emergence across human ontogeny. Methodological advances over the past 30 years have allowed scientists to probe the minds of even the youngest babies, showing that infants have a sophisticated set of abilities for perceiving, categorizing, and reasoning about social agents (Gergely, Bekkering, and Kiraly, 2002; Gergely, Nadasdy, Csibra, and Biro, 1995; Woodward, 1998; Woodward, Sommerville, and Guajardo, 2001), the physical world (Newcombe and Huttenlocher, 2005; Spelke, Breinlinger, Macomber, and Jacobson, 1992; Spelke, Lee, and Izard, 2010), and even abstract concepts such as number (Feigenson, Dehaene, and Spelke, 2004; Wynn, 1998). Yet whereas some skills emerge early in development, others have a much longer developmental trajectory. For example, whereas younger children rely primarily on geometric information to reorient in an environment, older children construct novel, flexible representations of space that combine both geometric and spatial information (Hermer and Spelke, 1994; Hermer-Vazquez et al., 2001; Hermer-Vazquez, Spelke, and Katnelson, 1999; Lee, Shusterman, and Spelke, 2006; Lee and Spelke, 2010; Wang, Hermer, and Spelke, 1999). That is, different cognitive skills can develop at different paces over ontogeny.
Despite extensive studies documenting the timing of cognitive development in humans, little is known about how human pacing compares to that of other animals. Understanding developmental pacing across species can illuminate differences in developmental start-points, or the earliest-emerging skills, as well as differences in the means by which later-emerging skills arise over development. Studies of evolutionary changes in developmental timing, or heterochrony, suggest that there are three broad categories of change in developmental pacing that can impact the expression of cognitive skills in mature individuals (Gould, 1977; Lieberman, Carlo, Leon, and Zollikofer, 2007; Wobber, Wrangham, and Hare, 2010a). First, two species could differ in their rate of skill acquisition (Figure 1a), developing a skill for a comparable period of time across the lifespan, but at differing speeds. Second, two species could differ in their start point of development (Figure 1b), either because one species exhibits developmental changes in a skill at an earlier age than the other, or because one species already possesses a higher level of skill at the time when developmental change begins. Finally, two species could differ in the age or life stage at which cognitive development concludes (Figure 1c). That is, one species could exhibit a longer period of development compared to another. It is important to note that these possibilities are not mutually exclusive. Consequently, two species could differ across all of these features, or none of them.

Differences in pace of development across species
The second major mechanism by which developmental changes could impact cognitive skills across species concerns the pattern of development, or differences in how development unfolds. In particular, a number of theories in psychology propose that specific skills are essential prerequisites for the acquisition of other skills, therefore requiring a specific pattern of development to acquire the mature adult skill level. For example, in the area of theory of mind, young children have been found to exhibit a relatively consistent pattern of success across different types of problems: children first begin to understand that other people can have conflicting desires, then distinguish between others' knowledge states, and finally understand that others may have different (or even false) beliefs about the world (Wellman and Liu, 2004, although see Onishi and Baillargeon, 2005). Similarly, in the physical domain, human children's capacities to track objects have been found to progress in a consistent pattern as well: infants first represent hidden objects, then overcome the A-not-B error, and finally master more complex relationships such as invisible displacement (Piaget, 1954). Some of these theories suggest that earlier-emegring abilities provide a developmental scaffold for later-emerging abilities. However, it is currently unclear if these same patterns of development are shared across other species.
There are three broad categories in which patterns of cognitive development could differ across species. First, two species could differ in their order of skill acquisition (Figure 2a). Here, early-emerging skills in one species could differ from the early-emerging skills in another. Second, two species could differ in the causal structure of skill acquisition, even if the general order of skills remains similar (Figure 2b). That is, later-emerging skills could be causally dependent on the acquisition of earlier-emerging skills in one species, whereas these causal links could be missing in the comparison species. Finally, some species could have terminal additions to their development, acquiring skills that are not seen in other species (Figure 2c). Even if developmental patterns are similar in earlier phases of development, one species might acquire skills that never arise in other species.
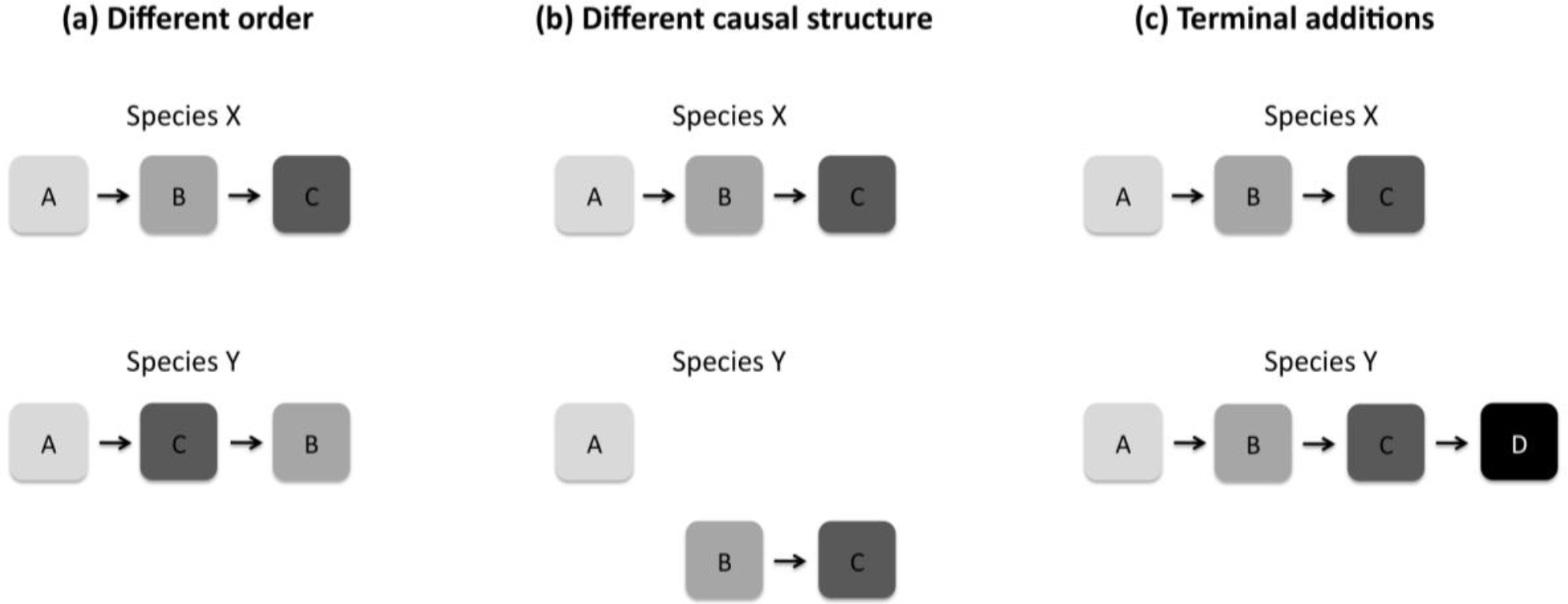
Differences in the patterns of development across species
Insights from comparative development for psychology
The models described above indicate that studies of comparative development can test psychological theories of human development. For example, some nativist theories of human development suggest that young infants possess a core set of fundamental, evolved cognitive capacities very early in life (e.g., Kinzler and Spelke, 2007). Others take a more constructivist stance towards conceptual change, emphasizing the importance of learning-based changes (Carey, 1985; Gopnik and Meltzoff, 1997; Tenenbaum, Griffiths, and Kemp, 2006; Wellman and Gelman, 1992). Finally, developmental systems theories focus more on the developmental process itself—that is, how the mechanisms supporting development evolve and change (Gottlieb, 1976, 1991; Lickliter, 2000). Importantly, these theories of human cognitive development make divergent predictions concerning the pace and pattern of development in non-human species.
The core knowledge theory, for example, suggests that some of the earliest-emerging skills in human infants reflect cognitive abilities that do not change significantly across development (Kinzler and Spelke, 2007). This view argues that a small set of domain-specific, experience-independent capacities forms the basis of our earliest understanding of the world, providing a scaffold on which later learning can build. Later in development, language may allow children to integrate old information in new ways (Hermer-Vazquez et al., 1999, 2001; Pyers et al., 2010). Within our comparative framework, early similarities in core knowledge across species would predict that human and non-human infants should possess a similar set of domain-specific precocious capacities in the first few months of life, exhibiting comparable pace and pattern in development during this period. Later in development, however, there should be more marked divergences between the human and non-human pace and pattern of development (coinciding with emerging language skills). These language-dependent shifts in human cognitive development could occur via a prolonged developmental time course following language emergence, or the terminal addition of novel skills (see Figures 1c and 2c). Studies of non-linguistic species can therefore target the degree to which language is necessary for certain ontogenetic changes to occur, versus whether language facilitates the emergence of some skills without being strictly necessary, by examining if or when non-humans acquire those skills.
Comparative developmental data can also test constructivist views of cognitive development, which hypothesize that cognitive skills may be fundamentally transformed across ontogeny. These types of views suggest that infants undergo a process of conceptual change throughout development such that their “theories” about the world are reorganized, (Carey, 1985; Gopnik and Meltzoff, 1997; Wellman and Gelman, 1992). Recent advances suggest that these changes may occur via Bayesian learning, whereby infants update their probability estimates for a hypothesis as they acquire additional evidence (Tenenbaum et al., 2006). Since Bayesian mechanisms support some types of learning across many species (Gold and Shadlen, 2007; Ma, Beck, Latham, and Pouget, 2006), the constructivist view would imply that species differences in cognition might arise from differences in initial start points (see Figure 1b) or in the total length of the developmental process (see Figure 1c), rather than via differences in the rate or pattern of learning throughout development, as suggested by nativist or core knowledge theories.
Insights from comparative development for biology
Studies of comparative cognitive development can also provide important insights for biology. The comparative method has long been one of the most important tools in evolutionary biology for reconstructing historical processes that often cannot be directly observed, such as evolution (Harvey and Purvis, 1991; Mayr, 1982). This method contrasts traits across species (including cognitive abilities; MacLean et al., 2012) that have faced differing ecological or social pressures in order to understand how natural selection influenced the manifestation of those traits (Harvey and Purvis, 1991; Mayr, 1982). As such, comparative biology can address not just what differences exist between species (e.g., the phylogeny of traits), but also why those differences arose (the evolutionary function of traits; Tinbergen, 1963).
Studies of comparative development have become increasingly important in the field of evolutionary developmental biology, which has revealed that species differences in mature traits frequently arise through alterations in developmental trajectories (Gould, 1977). In particular, novel traits are not typically created de novo. Rather, evolutionary shifts in the pattern and pace of developmental timing can generate variation among individuals and, ultimately, between species. One example of this process is the origin of variation in beak morphology across finch species living in the Galapagos Islands. As initially documented by Charles Darwin (1854), finches in this archipelago possess beaks of differing length and breadth, allowing different species to feed on different types of food. Recent research has revealed that these differences stem from variation in the expression over development of a gene controlling calcium production during beak growth (Abzhanov et al., 2006). That is, changes in the developmental timing of this (shared) gene generate diversity in adult beak morphology.
Importantly, this same approach used to examine morphological differences can be applied to cognitive or behavioral capacities. The human intellect has long been recognized as one of our species' most striking characteristics, creating an evolutionary puzzle for theorists since Darwin (1871). Comparative developmental studies provide a novel means to understand how the human mind emerged, and in particular to assess whether differences in human cognition stem from evolutionary changes in our species' developmental trajectories. More broadly, comparative studies of cognitive development allow complex behavioral patterns to be broken down into their constituent cognitive processes, which may be dissociable in terms of their differing developmental patterns. Thus, this approach facilitates evolutionary analyses of cognition as an important biological trait that also exhibits variation across species, much like morphological traits that have been the focus of previous developmental work.
Empirical Approaches to Comparative Developmental Psychology
The previous sections highlighted how the comparative developmental approach can address outstanding questions in psychology and biology, and proposed theoretical models of how variation in the pace and pattern of development can produce variation in cognitive abilities across species—including humans. In the current section, we apply these models to three empirical domains that have been the focus of recent research on comparative development: 1) reasoning about objects, 2) foraging cognition, and 3) social cognition.
Development of object reasoning in primates
Since Piaget's (1954) early work examining how human children come to understand that objects continue to exist when they are out of sight (e.g., object permanence), children's development of physical concepts has been an area of active research and debate. Some researchers claim that infants only acquire rich representations of objects by interacting with the physical world (Smith and Gasser, 2005), whereas others suggest that, in the absence of direct evidence, even very young infants are equipped with sophisticated abilities to represent objects (Feigenson et al., 2004; Spelke et al., 1992). Comparative developmental studies with other animals—whose interactions with the physical world have both similarities and differences from those seen in humans—can therefore address the role of experience in generating children's mature abilities to reason about objects.
One of the first developmental studies of physical reasoning examined object classification (Langer, 1980, 1986). This work identified a basic developmental pattern underlying infants' object classification in spontaneous play behavior. During their first year, infants play at creating sets of objects, and eventually form sets that share similar properties. In the second year of life, children construct object sets with complex properties, and in the third year they begin to relate objects physically—for example, by making buildings and bridges. Finally, in their fourth year, children play with multiple object sets simultaneously and show mutability of sets, such as first sorting by color and then later by material (Langer, 1980, 1986). These studies indicate that spontaneous play in humans appears to have a relatively fixed pattern of development, with certain skills (such as sorting sets) emerging reliably before other skills (such as encoding multiple sets).
Does object play reflect a human-specific developmental trajectory, or do these patterns reflect a deeply rooted cognitive structure that other primates share? Studies of object play in several species, including chimpanzees (Pan troglodytes), capuchins (Cebus apella), and longtailed macaques (Macaca fascicularis), in fact suggest that their basic order of skill acquisition in object play is similar to that of human infants, even though these species lack much of the experience with objects that human infants have (Hayashi and Matsuzawa, 2003; Poti, 1997; Poti, Hayashi, and Matsuzawa, 2009; Spinozzi, 1983; Spinozzi and Natale, 1989). However, there are also important differences from humans. For example, monkeys of all ages played with set creation, but did not consistently create similarity sets until 2 to 3 years of age. Development was quicker in chimpanzees, but still slower than in humans, and the apes never showed the most complex forms of object constructions. Overall, these results suggest that apes (including humans) exhibit a faster pace of development than do monkeys in skills of object manipulation (as in Figure 1a). Nonetheless, the most complex skills that humans exhibit may be terminal additions in human development (as in Figure 2c).
Another important skill for reasoning about the physical world involves tracking the locations of objects as they move through space. In humans, object-tracking skills emerge in a distinct order. First, infants first begin to retrieve hidden objects (before 10 months of age), and then begin to comprehend visible displacements, solving the “A not B” error, around their second birthday (Call, 2001; Piaget, 1954). Soon after, they solve invisible displacements, but it is not until relatively late in development (3 to 4 years) that children understand complex invisible displacements such as transpositions (changing the location of an object's container) and rotational displacements (where the support is moved; Piaget, 1954; see also Call, 2001; Lasky, Romano, and Wenters, 1980; Okamoto-Barth and Call, 2008; Sophian, 1984). One hypothesis for this consistent pattern of emergence is that all object-tracking skills rely on the same set of cognitive capacities. As these capacities develop, children can complete more complex tasks, such as those involving out-of-sight movements or larger numbers of items (Barth and Call, 2006; Call, 2003). If this is the case, then non-human primates should exhibit similar developmental patterns. Alternatively, these behaviors may rely on a set of independent skills, in which case the patterns of emergence might differ across species.
Some data from adult primates support the claim that these diverse object-tracking behaviors rely on the same cognitive capacities. In particular, both monkeys and apes are successful at “easier” tasks involving visible displacements, but generally only apes are capable of the full range of object-tracking skills, including those with invisible movements (Albiach-Serrano, Call, and Barth, 2010; Barth and Call, 2006; Beran, Beran, and Menzel, 2005; Beran and Minahan, 2000; Call, 2001; Collier-Baker, Davis, Nielsen, and Suddendorf, 2006; de Blois, Novak, and Bond, 1998, 1999; Tinklepaugh, 1928; but see Filion, Washburn, and Gulledge, 1996; Mendes and Huber, 2004; Neiworth et al., 2003). However, other evidence from adult primates does not accord with the patterns seen in humans. For example, even though adult monkeys are not successful at reasoning about invisible displacement tasks, they can understand rotational displacements, which is surprising because this requires mental reconstruction of the movements of hidden objects (Hughes, Mullo, and Santos, 2013; Hughes and Santos, 2012; Poti, 2000). Furthermore, adult apes show similar levels of errors on tasks that seem to differ in their difficulty for human children, such as rotational displacements and translocations (Albiach-Serrano et al., 2010) or transpositions and invisible displacements (Barth and Call, 2006). These findings therefore cast doubt on the hypothesis that the same core skill set underlies performance across these tasks in non-human primates.
Comparative developmental studies also suggest that object-tracking tasks tap into a range of abilities that differ in their time and pattern of emergence. The pattern of success across development by non-human primates in object-tracking tasks broadly mirrors that seen in humans, but appears to proceed at a faster pace at early stages (as in Figure 1a; see also Gomez, 2005). However, for a few select tasks, non-humans show different patterns from those seen in humans, both in terms of the order of emergence (as in Figure 2a) and the causal structure of skills (as in Figure 2b). In particular, the ability to follow translocations develops at a later life-stage in non-human primates than in humans. For example, gorillas first solve invisible displacements by 12 months (Dore and Goulet, 1998), yet they learn to solve translocations sometime after 17 months (Visalberghi, 1986)—the opposite order than that observed in children. Similarly, although human children tend to solve “single” and “double” displacement tasks (which differ in the number of objects that move) around the same time (Call, 2001; Piaget, 1954), double displacements appear to be markedly more difficult for young non-human primates (Antinucci, 1990). Indeed, one young gorilla solved invisible displacements many months before he was able to understand a double visible displacement (Antinucci, Spinozzi, and Natale, 1986; Dore and Goulet, 1998). Overall, these developmental data indicate that object-tracking abilities do not develop in a uniformly consistent manner across species. Consequently, object tracking may indeed depend on multiple underlying cognitive capacities in human development. Comparative developmental inquiry, therefore, provided a key test for theories that could not be fully evaluated with human developmental data alone.
Development of foraging skills in chimpanzees and bonobos
A second area where comparative developmental research has recently addressed the origins of human cognition concerns cognitive skills used in foraging contexts. Wild primates face a myriad of complex spatial problems when foraging, including recalling the location of resources and determining the best route to navigate between these different locations. Despite the ubiquity of such spatial problems, psychological theories suggest that humans are uniquely equipped to solve spatial problems with a degree of flexibility and accuracy not seen in other species. In particular, language is thought to play an important role in human spatial cognitive development (Haun, Rapold, Call, Janzen, and Levinson, 2006; Hermer and Spelke, 1994; Hermer-Vazquez et al., 1999, 2001; Levinson, Kita, Haun, and Rasch, 2002; Pyers et al., 2010). In terms of spatial memory, children exhibit developmental shifts around 2 years of age in many skills, including recall of the locations of hidden targets, recall of multiple locations, and recall of locations over longer temporal periods (Balcomb, Newcombe, and Ferrara, 2011; Newcombe and Huttenlocher, 2005; Newcombe, Huttenlocher, Drummey, and Wiley, 1998; Sluzenski, Newcombe, and Satlow, 2004). Some aspects of these changes may be related to language. For example, when children are asked to search for hidden objects in a large space after they have been disoriented, their proficiency with spatial language predicts the success of their searches (Balcomb et al., 2011). However, the relationship between language and the emergence of a given cognitive ability could be causal (language is required for a skill to emerge), facultative (language speeds up a process that could occur otherwise), or only correlated. Studies of comparative development in apes can therefore be an important tool in differentiating these possibilities, as apes share an extended juvenile period of post-natal brain development with humans but lack language (Matsuzawa, 2007; Matsuzawa et al., 2006).
Increasingly, research indicates that apes have quite sophisticated spatial abilities (Kuhlmeier and Boysen, 2002; Mendes and Call, 2008; Menzel, Savage-Rumbaugh, and Menzel, 2002; Menzel, 1973). Studies tracking the emergence of spatial skills over development further suggest that apes can show human-like changes in spatial abilities in the absence of language. For example, one study examined how chimpanzees and bonobos performed in a naturalistic foraging task where they could search for food they had seen hidden previously in a large outdoor enclosure (Rosati and Hare, 2012b). Whereas infant apes found few pieces of food, older chimpanzees were more successful at recalling multiple locations. Thus, although there are improvements in place memory associated with the acquisition of spatial prepositions in humans (Balcomb et al., 2011), chimpanzees exhibit similar developmental changes without any concurrent language acquisition. This suggests that at least some types of ontogenetic shifts in place-based searching may be due to intrinsic, maturational changes in spatial abilities. In addition, although language may be a developmental prerequisite for the origins of some spatial skills in humans (Hermer-Vazquez et al., 2001), this causal link is not present in apes (following Figure 2b).
Comparative developmental studies also make it possible to address why such differences in developmental trajectories arise, by linking developmental data with information about different species' natural histories. For example, in the foraging task described previously (Rosati and Hare, 2012b), chimpanzees and bonobos exhibited similar skills in infancy, but chimpanzees exhibited a faster rate of improvement than bonobos (following Figure 1a). Why might this be the case from an evolutionary perspective? Importantly, even though chimpanzees and bonobos diverged less than 1mya (Won and Hey, 2005), they show a suite of differences in their behavior and morphology. Chimpanzees exhibit more pronounced sexual dimorphism, increased escalated aggression, and higher rates of extractive foraging and hunting, whereas bonobos exhibit increased sociosexual behavior and an increased importance of female bonds (Hare, Wobber, and Wrangham, 2012; Kano, 1992; Parish, 1996; Parish and de Waal, 2000; Wrangham and Pilbeam, 2001). One hypothesis for this pattern is that chimpanzees and bonobos differ in their wild feeding ecology (Kano, 1992; Wrangham and Peterson, 1996). In particular, chimpanzees are thought to live in environments where they face higher costs to acquire food, including more seasonal variation (White, 1998), more competition for smaller, less-abundant food patches (White and Wrangham, 1988), and have less access to important fallback foods (Malenky and Wrangham, 1993)—a set of differences that is thought to impact the tenor of social interactions in this species. This ecological hypothesis further suggests that chimpanzees develop more accurate spatial memory over ontogeny due to their greater dependence on patchily-distributed resources as adults.
Comparisons of chimpanzees and bonobos therefore provide a test case for the role of comparative developmental psychology in illuminating the functional relevance of cognitive capacities. Increasing evidence suggests that adult chimpanzees and bonobos actually differ in a set of cognitive traits related to foraging, including levels of feeding tolerance (Hare, Melis, Woods, Hastings, and Wrangham, 2007; Wobber, Wrangham, and Hare, 2010b), tool use abilities (Herrmann, Hare, Call, and Tomasello, 2010), spatial memory (Rosati and Hare, 2012b), and patterns of value-based decision-making (Heilbronner, Rosati, Hare, and Hauser, 2008; Rosati and Hare, 2011, 2012a, 2013; Rosati, Stevens, Hare, and Hauser, 2007). One potential evolutionary mechanism to generate these differences is heterochronic changes in dvelopmental timing. Indeed, developmental comparisons have revealed that even though patterns of cognitive development are broadly similar for both species, bonobos appear to exhibit delayed development specifically in this set of skills relevant to the ecology hypothesis, such as in inhibitory control and memory (Herrmann, Hare, et al., 2010; Rosati and Hare, 2012b; Wobber, Herrmann, Hare, Wrangham, and Tomasello, 2013; Wobber et al., 2010b). That is, chimpanzees and bonobos exhibit targeted differences in development only for a specific set of ecologically-relevant skills.
Comparative developmental studies of closely related species (like chimpanzees and bonobos) can allow researchers to integrate hypotheses about the mechanisms supporting cognition with hypotheses about the functional importance of those cognitive skills in natural behavior. Studies that further include both humans and non-human apes can therefore provide critical insights into the evolutionary function of uniquely-human cognitive skills. For example, human feeding ecology differs from that of other great apes in several ways. Hunter-gatherers have larger home ranges than other apes, and exhibit a unique pattern of central place foraging where individuals return to a centralized location with food (Hill, Barton, and Hurtado, 2009; Marlowe, 2005). Consequently, humans are more reliant on distant food sources than are other apes, which poses new problems concerning locating food and navigating between resources. The results from chimpanzees and bonobos predict that humans may also exhibit targeted developmental differences in evolutionarily-relevant skills. For example, over development, children form increasing flexible representation of space integrating multiple cues (Hermer-Vazquez et al., 1999, 2001; Newcombe and Huttenlocher, 2005), which may be related to the new ecological problems that humans must solve. Comparative developmental studies can potentially illuminate the function of these human cognitive abilities.
Development of social cognition in apes and humans
A final area where recent comparative developmental work has advanced our knowledge is that of social cognition. Humans and non-human primates differ not only in how they reason about the physical world, but also in their reasoning about the social world. Humans possess a sophisticated belief-desire psychology that we use to reason about the (unobservable) thoughts, intentions, and beliefs of other people. Other primates share some of the same capacities, such as reasoning about other's perceptions, but they also differ in important ways (Call, 2008; Rosati, Santos, and Hare, 2010; Wobber and Hare, in press). In fact, one influential hypothesis suggests that social cognition represents the most profound difference between us and other animals (Tomasello, 1999; Tomasello and Carpenter, 2007; Tomasello et al., 2005). Recent studies have begun to disentangle how these differences emerge.
First, humans and other primates exhibit many striking similarities in their social cognition in the first few months of life. Chimpanzee and rhesus macaque neonates, for example, both imitate the facial expressions of a human demonstrator in their first few days of life (Ferrari et al., 2006; Myowa-Yamakoshi, Tomonaga, Tanaka, and Matsuzawa, 2004), much like babies (Meltzoff and Moore, 1983). In addition, infant chimpanzees and an infant gibbon were found to prefer faces with direct gaze over averted-gaze faces by the age of 2 months (Myowa-Yamakoshi and Tomonaga, 2001b; Myowa-Yamakoshi, Tomonaga, Tanaka, and Matsuzawa, 2003), paralleling the “2-month revolution” when humans infants begin to actively engage with other's gaze (Rochat, 2001). Finally, infant chimpanzees and an infant gibbon recognize and prefer the face of their mother or human caregiver over less familiar faces at 1 month of age (Myowa-Yamakoshi and Tomonaga, 2001a; Tomonaga et al., 2004), much like human infants (Pascalis, Deschonen, Morton, Deruelle, and Fabregrenet, 1995). Taken together, these results show that humans and apes exhibit similar developmental trajectories in early infancy, suggesting that human patterns have deep biological roots.
However, human and non-human primate social development diverges at older ages. For example, human infants reliably follow other's gaze to targets by 6 months (Butterworth and Jarrett, 1991) and begin to follow gaze in more complex situations, where barriers make it difficult to gauge another's line of sight, by 12 months (Moll and Tomasello, 2004). In contrast, a longitudinal study of a chimpanzee's gaze-following skills indicates that he did not follow a human experimenter's gaze until 13 months (Okamoto et al., 2002). Other studies find that chimpanzees are not proficient in geometric gaze-following or following gaze around barriers until 2 to 3 years old (Okamoto-Barth, Tomonaga, Tanaka, and Matsuzawa, 2008; Tomasello and Carpenter, 2005; Wobber et al., 2013). These results indicate that humans show an accelerated pace of social-cognitive development in this period of late infancy (following Figure 1a). Macaques also develop gaze-following skills at a slower rate than humans (Ferrari, Coude, Gallese, and Fogassi, 2008; Ferrari, Kohler, Fogassi, and Gallese, 2000; Teufel, Gutmann, Pirow, and Fischer, 2010), leading to the proposal that non-human gaze-following is spurred by experience with relevant social interactions over a long critical period. This proposal would therefore suggest a difference between humans and other primates, both in the rate of development (see Figure 1a) as well as a difference in the underlying causal structure of gaze-following skills (see Figure 2b). Indeed, mechanisms supporting gaze-following in adult chimpanzees and humans may differ as well. Eye-tracking studies show that although humans are most interested in looking at eyes, chimpanzees are as interested (or even more interested) in viewing the bodies or mouths of conspecifics (Kano and Tomanaga, 2009; Myowa-Yamakoshi, Scola, and Hirata, 2012).
Another area of divergence in human and non-human social cognitive development concerns the perception of goal-directed action. Human infants rapidly gain proficiency in understanding goal-directed actions: They interpret the acquisition of desirable objects as being the goal of a reaching gesture by 6 months, and reason about the rationality of an experimenter's goal-directed action by 14 months (Gergely et al., 2002; Woodward, 1998). In contrast, chimpanzees only first begin to reason about others' actions in terms of their underlying goals around 2 to 3 years of age, even when reared by humans (Bering, Bjorklund, and Ragan, 2000; Tomasello and Carpenter, 2005; Wobber et al., 2013). Thus, both gaze following and goal understanding appear to emerge relatively late in chimpanzees compared to humans. This again supports the claim that there are differences in both the pace (see Figure 1a) and pattern (see Figure 2b) of human social cognitive development, suggesting that behavioral skills that quickly come online in relatively young humans may depend on additional social experience in non-humans.
Earlier differences in human and primate social cognition may have further cascading effects over development. Human infants typically rely on mechanisms of joint attention to comprehend and direct others' attention and behavior following the “9-month revolution.” At this point, infants first begin to move beyond simple dyadic interactions (self and other) to engage triadic interactions (self, other, and referent; Carpenter, Nagell, and Tomasello, 1998; Tomasello, 1999). In contrast, chimpanzee infants do not seem to exhibit joint attention, even though they can understand and even direct others' behavior at older ages (Tomonaga, 2006; Tomonaga et al., 2004; Wobber et al., 2013). Overall, these findings indicate that although joint attention may be a causal precursor of other skills in humans (such as more complex forms of gaze following, as described previously), this causal structure is lacking in apes (as in Figure 2b). These results highlight that even though certain capacities may be prerequisites for later-developing abilities in humans, these same capacities may not be tightly linked in other species.
Indeed, even enculturated apes—who have been reared in a human-like social and communicative environment—do not exhibit capacities for joint attention. Enculturation studies can assess the degree to which ape developmental trajectories are plastic in response to human-like social inputs. Enculturated apes can use some symbolic language, employ referential signals outside of the linguistic domain (Premack and Premack, 1983; Terrrace, 1979), imitate others (Bjorklund, Bering, and Ragan, 2000; Call and Tomasello, 1996), and even reason about novel tools like humans (Furlong, Boose, and Boysen, 2008). However, they do not develop human-like joint attention (Carpenter, Tomasello, and Savage-Rumbaugh, 1995; Tomasello and Carpenter, 2005; but see Bard et al., 2005 for conflicting definitions of joint attention). These findings suggest that important differences between human and ape social cognition persist even when apes are reared in more human-typical environments.
Finally, the importance of these early differences in social-cognitive development may increase in later ontogeny. Tomasello and colleagues hypothesized that the early emergence of social cognitive skills like joint attention and goal understanding allows humans to rapidly acquire a diverse array of skills across domains (Herrmann, Call, Hernadez-Lloreda, Hare, and Tomasello, 2007; Tomasello and Carpenter, 2007; Tomasello et al., 2005). In particular, skills emerging during the “9-month revolution” (Tomasello, 1999), such as the ability to engage in triadic interactions, appear to enable human infants to learn proficiently from other individuals—including social knowledge such as novel word labels (Baldwin and Moses, 2001; Vaish, Demir, and Baldwin, 2011), as well as non-social knowledge about how to correctly utilize novel tools (Birch, Vauthier, and Bloom, 2008; Casler and Kelemen, 2005). This hypothesis predicts that because non-humans do not acquire this earlier set of social skills, they will begin to exhibit divergences in both social and non-social skills in later ontogeny.
A recent comparison of cognitive development in 2- to 4-year-old humans, chimpanzees, and bonobos assessed this hypothesis by comparing the performance of these species across a diverse array of cognitive tasks. This comparison found that in the social domain (in tasks assessing goal understanding, gaze-following, and imitation, among other capacities), human 2-year-olds were more proficient than same-age non-human apes. However, humans, chimpanzees, and bonobos had comparable performance at this age in tasks measuring cognitive skills in the physical domain (such as assessing comprehension of space, number, and causality). The critical test of the hypothesis was thus to examine humans' and apes' relative change in skills as they aged. In support of the claim that early social differences have cascading effects, humans showed greater improvement in their performance on both social and physical tasks compared to apes (Wobber et al., 2013). These results suggest that children's early competency in social cognitive skills such as goal understanding and imitation might enable them to learn more rapidly across domains, whereas young apes must rely to a greater degree on individual and trial-and-error learning, acquiring knowledge about the world at a slower rate. That is, the causal relationships between the acquisition of different types of skills differs between apes and humans (as in Figure 2b) may in turn facilitate differences in the rate of development later in ontogeny (as in Figure 1a). Concordant with this suggestion, a study of individual differences in performance across a similar battery of tasks found that children possess a distinct social cognition “factor” predictive of their success in tasks across that domain, whereas chimpanzees' success on different social cognition tasks was not interrelated (Herrmann, Hernández-Lloreda, Call, Hare, and Tomasello, 2010). Taken together, these results indicate that human social-cognitive development differs from that of our closest relatives in both pace and pattern. Moreover, these early differences in social abilities may underpin major differences in the adult cognition of these species.
Conclusions: The Future of Comparative Developmental Psychology
We have argued that comparative developmental research can provide new insights into human cognition that cannot be identified via developmental or comparative research alone. Although this approach is not commonly used, recent research has already begun to identify similarities and differences in the pace and pattern of human development relative to other primates in domains as diverse as reasoning about objects, spatial memory, and social cognition. Yet this empirical work represents a small sample of the open questions in psychology and biology that the comparative developmental approach can address. Indeed, there are some cognitive domains where it is already clear that comparative developmental studies can distinguish between alternative accounts of cognitive development and move theoretical debates forward.
For example, one current debate concerns the development of different capacities supporting mature theory of mind. As mentioned previously, results from theory of mind tasks suggest that these skills emerge in a strict order, with children first understanding others' desires, then their knowledge, and finally their beliefs (Wellman and Liu, 2004). However, recent work using looking-time measures to test younger children find that babies may understand something about false beliefs within the second year of life (Baillargeon, Scott, and He, 2010; Kovács, Téglás, and Endress, 2010; Onishi and Baillargeon, 2005; Southgate, Senju, and Csibra, 2007; Surian, Caldi, and Sperber, 2007). Comparative studies of non-human primates further indicate that species such as chimpanzees and rhesus macaques exhibit a number of theory of mind skills (see Rosati et al., 2010 for a review). These skills include visual and auditory perspective-taking (Braeuer, Call, and Tomasello, 2007; Flombaum and Santos, 2005; Hare, Call, Agnetta, and Tomasello, 2000; Hare, Call, and Tomasello, 2006; Melis, Call, and Tomasello, 2006; Santos, Nissen, and Ferrugia, 2006), reasoning about others' knowledge states (Hare, Call, and Tomasello, 2001; Kaminski, Call, and Tomasello, 2009; Marticorena, Ruiz, Mukerji, Goddu, and Santos, 2011), and desire comprehension (Buttelmann, Call, and Tomasello, 2009). Critically, however, non-human primates do not appear to represent the beliefs of others when tested in false belief tasks (Call and Tomasello, 1999; Kaminski et al., 2009; Krachun, Carpenter, Call, and Tomasello, 2009; Marticorena et al., 2011), and may instead represent situations in which an individual has a false belief merely in terms of that individual's ignorance (Kaminski et al., 2009; Marticorena et al., 2011). Comparative developmental studies can thus assess whether the patterns of emergence of different theory of mind capacities in non-human primates are in fact causally dependent on one another, and illuminate why young infants might be successful on non-verbal false-belief tasks whereas non-humans are not. Alternatively, non-human primates' capacities may emerge through different developmental mechanisms, suggesting that their abilities are supported by different representations about social agents.
A second debate that comparative developmental studies can address concerns the development of our capacity to individuate objects. Over the past decade, developmental researchers have examined how infants develop the ability to distinguish objects of different categories. Classic work by Xu and colleagues suggested that emerging language capacities—specifically, learning the labels of different objects—may be necessary for conceptually individuating different kinds of objects (Xu, 2002; Xu and Carey, 1996; Xu, Carey, and Quint, 2004; Xu, Carey, and Welch, 1999). Indeed, Xu and colleagues observed that infants are unable to individuate two different objects in a looking-time task before they know the words for those objects (Xu and Carey, 1996). To test the role of language, primate researchers have begun to examine whether adult non-human primates can succeed on similar individuation tasks. Surprisingly, this work revealed that adult primates are able to individuate objects, even though they lack their linguistic labels (Munakata, Santos, Spelke, Hauser, and O'Reilly, 2001; Phillips and Santos, 2007; Santos, Sulkowski, Spaepen, and Hauser, 2002). How then do non-human primates learn to solve individuation problems that human infants solve with language? Comparative developmental studies can address this topic by focusing on the emergence of these mechanisms in non-human primate ontogeny. For example, monkeys may acquire object individuation skills at a slower pace than humans, suggesting that language may play a facultative role in accelerating human development of these skills. Comparative developmental studies can also identify whether primates' individuation capacities depend on another form of experience that operates similarly to the way human language facilitates individuation in our own species.
Finally, a major question in biology that comparative developmental studies can address concerns the evolutionary significance of differences in human life history traits (i.e., the timing and patterning of key life events such as age of sexual maturity and reproduction). Whereas all primates exhibit a long juvenile period relative to other mammals (Charnov and Berrigan, 1993), the human juvenile period is markedly longer than expected relative to other primates (Bogin and Smith, 1996; Walker, Burger, Wagner, and Von Rueden, 2006). One hypothesis for the evolutionary function of this prolonged juvenile period is that it enables more complex skill acquisition and, therefore, greater behavioral flexibility in human adulthood (Bjorklund and Green, 1992; Bogin, 2010; Kaplan, Hill, Lancaster, and Hurtado, 2000). Yet although there is morphological and physiological evidence indicating that human growth and reproduction is slowed relative to that of other primates, there are little data on comparative cognitive development examining whether humans actually exhibit longer periods of skill acquisition. Indeed, some evidence comparing rates of cognitive development in humans and apes suggests that human cognitive development may in fact be accelerated in comparison to that of non-human primates during early childhood, rather than prolonged (Wobber et al., 2013). Comparative developmental studies of apes and humans at older ages—such as during adolescence, when humans are learning some of their most complex, culturally-acquired skills—can further test this hypothesis.
Overall, we emphasize that studies of comparative cognitive development can address outstanding questions in both psychology and biology. Research using this approach has already made important contributions to our understanding of the human mind. Indeed, the results discussed here highlight that models of human cognitive evolution must integrate data on cognitive development in other species in order to draw strong inferences about what traits are unique to our lineage. Thus, the comparative developmental approach represents an untapped line of evidence that will advance theoretical ideas concerning what cognitive abilities are unique to humans, as well as the ultimate evolutionary function of these abilities in human evolutionary history.
Footnotes
Acknowledgements:
These ideas were developed at an International Conference for Infant Studies (2012) symposium, and we thank attendees and commentators. A.G.R.'s research is supported by a grant from the L.S.B Leakey Foundation. V.W.'s research is supported by grants from the L.S.B. Leakey Foundation, NSF-BCS-0851291 (DDIG), and the Wenner-Gren Foundation. L.R.S. is supported by a McDonnell Scholar Grant (#220020242).