
Abstract
We tested a prediction that females' duration estimates of briefly-viewed male, but not female, photos would be modulated by attractiveness. Twenty-seven female participants viewed sequences of five stimuli of identical duration in which the first four were sine-wave gratings (Gabor discs) and the fifth was either the same sine-wave grating (control trials) or a photo of an attractive or unattractive male or female (test trials). After each sequence, participants had to reproduce the duration of the fifth stimulus. Results confirmed our prediction and showed that duration estimates of attractive male photos were significantly longer than corresponding estimates for unattractive male photos, while there was no significant difference in estimated duration for attractive and unattractive female photos. Our data show that unexpectedly viewing an attractive male affects time perception in females, and are the first demonstration that stimuli relevant to reproductive fitness, which engage the appetitive motivational system, can increase perceived duration.
Introduction
What happens when we unexpectedly see an attractive, opposite-sex person? For a woman, the sight of a physically attractive man is likely to engage the appetitive motivational system, which mediates physiological reactions that underlie affective reactions such as pleasure (e.g., Lang and Bradley, 2010, for review). There is evidence of autonomic response for both men and women when viewing erotic photographs (e.g., increased skin conductance, cardiac deceleration, and startle reflex modulation; Bradley, Codispoti, Cuthbert, and Lang, 2001), and similar results are found with fully clothed models. For example, Laeng and Falkenberg (2007) found that women's pupils dilated more when viewing photos of their boyfriends and favorite male actors (e.g., Brad Pitt) than photos of female actresses or the boyfriends of the other participants. Of particular interest was that this effect was greatest during the ovulatory phase of the menstrual cycle, and was not obtained for women who were taking oral contraceptives.
Psychophysiological studies using electromyographic (EMG) techniques to measure facial muscle activity have shown that females' responses to photos of attractive or unattractive persons depends on whether the targets are male or female (Hazlett and Hoehn-Saric, 2000). Attractive male photos were associated with greater zygomatic EMG activity (i.e., smiling) and rated as more pleasant and arousing than unattractive male photos, whereas viewing of attractive female photos resulted in greater corrugator muscle EMG activity (i.e., eyebrow lowering) than unattractive female photos. Hazlett and Hoehn-Saric (2000) interpreted the corrugator EMG results (which are associated with frowning) as evidence of a defensive reaction to viewing a high-status competitor.
Research has also found that viewing photos of attractive persons influences automatic or implicit cognitive processes. Maner et al. (2003) showed arrays of photos with attractive and unattractive males, or attractive and unattractive females, to participants who were required to estimate the percentage of attractive photos in each array. Maner et al. found that limiting attentional capacity by restricting the time to view the photos, or by presenting them all at once rather than sequentially, increased the frequency estimates for attractive females but not attractive males. Interestingly, these results were obtained for both male and female participants. Maner et al. suggested that both sexes had a selective processing bias for attractive female targets – an automatic bias which they described as “female beauty captures the mind” or “attentional adhesion” (see also Maner, Gailliot, Rouby, and Miller, 2007).
Exposure to attractive opposite-sex photos can also influence evaluative judgments, mood, and self-esteem. Kenrick, Montello, Gutierres, and Trost (1993) found that male and female participants' moods decreased after viewing an attractive same-sex, but not opposite-sex, photo. Gutierres, Kenrick, and Partch (1999) reported that women's self-assessed suitability as a marriage partner was decreased after viewing photos of highly attractive women. Little and Mannion (2006) found evidence of a contrast effect in women's mate choices: Self-rated attractiveness varied inversely with exposure to attractive or unattractive same-sex photos, consistent with Kenrick et al. (1993), but so did preference for facial masculinity in a potential male partner. According to Little and Mannion, the latter result indicated that women's mate preferences were “market-value-dependent” in that they depended on the woman's current evaluation of her own attractiveness compared to potential competitors.
Overall, there is considerable evidence that exposure to photos of attractive opposite-sex and same-sex individuals is associated with a range of physiological and cognitive processes from involuntary autonomic responses to evaluative self-judgments. The goal of the present study was to test a hypothesis derived from an evolutionary rationale – that women's estimates of how long they have viewed a photo will depend on attractiveness for male, but not female, photos. Specifically, we predicted that women will judge the duration of a briefly-viewed attractive male photo to be longer than an unattractive male photo of the same duration, but that duration estimates of attractive and unattractive female photos should not differ systematically.
Our hypothesis is based on the rationale that the human timing system contains adaptations, which provide flexibility in situations related to fitness. According to the leading theory of timing (Scalar Expectancy Theory), perception of short intervals of time is based on a pacemaker-accumulator system (Gibbon, 1977; Wearden, 2003). During a stimulus, the pacemaker generates pulses at a high and variable rate, which are stored in an accumulator. Because subjective time is directly related to the number of pulses stored, it is affected by changes in the rate of the pacemaker. Research has shown that subjective time often slows down (i.e., increased subjective duration) in situations which activate the defensive motivational system (Bradley and Lang, 2010), such as life-threatening events (Ursano et al., 1999), which can be explained as an increased pacemaker rate due to arousal. Recent studies have found that duration estimates for emotion-provoking stimuli, such as angry faces, are longer compared to neutral stimuli (Droit-Volet, Brunot, and Niedenthal, 2004; Effron, Niedenthal, Gil, and Droit-Volet, 2006; Noulhiane, Mella, Samson, Ragot, and Pouthas, 2007), which has also been attributed to increased pacemaker rate.
Has the timing system evolved to enhance reproductive success, in addition to preparing the organism for a defensive action? It would be advantageous for subjective time to increase when seeing an attractive potential mate, as this would enhance their salience and allow more information to be processed quickly. In that case, the unexpected sight of an attractive opposite-sex person should increase arousal and the rate of the pacemaker, leading to an expansion of subjective time. Thus we predict females' estimates of the duration of briefly-viewed attractive male photos should be greater than corresponding estimates for unattractive male photos.
To obtain duration estimates, we used a psychophysical task – the “oddball” procedure – that seemed to capture some of the aspects of the real-world situation we wished to study; that is, unexpectedly seeing an attractive, opposite-sex person. In a typical oddball task, the observer must detect a distinctive stimulus in a stream of identical stimuli. The oddball paradigm has been used extensively with both visual and auditory stimuli to measure event-related potentials (ERPs) and electrophysiological correlates of attention-related processes (e.g., Garcia-Larrea, Lukaszewicz, and Mauguierre, 1992; Polich, 1986; see Goodin, 1990, and Pfefferbaum, Ford, and Kraemer, 1990, for review). More recently, Tse, Intriligator, Rivest, and Cavanagh (2004) used a variety of oddball procedures to study how attentional processes affect perceived duration. For example, in their Experiment 1A observers viewed a series of identical black discs (standard stimuli), all of which lasted for 1050 ms, in which occasional expanding discs (oddballs) appeared, and they had to indicate whether the oddball was shorter or longer in duration than the standard stimuli. Across different experiments, Tse et al. found that duration estimates of oddballs increased for stimuli lasting longer than about 375 ms, which they interpreted in terms of a latency to engage attentional processes. Tse et al. proposed that the oddball stimulus would capture attention and result in a greater number of pacemaker counts (in terms of Scalar Expectancy Theory) being stored, and hence a longer subjective duration. Subsequent research has generally confirmed Tse et al.'s conclusions, and has shown that the subjective duration of longer oddball stimuli (500 ms – 2000 ms) was about 120% of the duration of standard stimuli (Seifried and Ulrich, 2010).
We adapted Tse et al.'s method of stimulus duration reproduction (SDR; see Tse et al., 2004, Experiment 6) to test whether females' duration estimates of male, but not female, photos depended on attractiveness. In our study, female participants viewed sequences of five stimuli. The duration of all stimuli were identical on each trial, and were varied across trials. The first four stimuli were always neutral and the fifth stimulus was either identical to the first four (control trials), or a photo of an attractive or unattractive person (test trials). In different blocks, test trials included male or female photos. The duration of stimuli on each trial ranged from 133 ms to 2100 ms, and participants had to reproduce the duration of the fifth stimulus by pressing the mouse button. We predicted that the reproduced durations of attractive male photos would be longer than the reproduced durations of unattractive male photos, but that there would be no significant difference in reproduced duration of attractive and unattractive female photos.
Materials and Methods
Participants
Twenty-seven women enrolled at a liberal arts college in the eastern USA were recruited for the study. Their average age was 22.4 years (minimum = 18; maximum = 30; SD = 2.37). Each received course credit in return for their participation.
Apparatus and Stimuli
An IBM-compatible PC controlled all experimental events, and responses were registered through the mouse. The experimental program was written in PsychoPy (Peirce, 2007).
All stimuli were presented in the center of the screen (13.3″ diameter) on a gray background. The neutral stimuli were circular Gabor discs, which are sinusoidal wave gratings modified by a Gaussian filter (Yao, Krolak, and Steele, 1995). Although Gabor discs can vary in frequency and orientation, in the present study all neutral stimuli were an identical Gabor disc, 5 cm in diameter, with an orientation 10 degrees clockwise from vertical and frequency of 3 cycles/disc.
The test stimuli were color photos of attractive and unattractive adult males and females. From an initial pool of potential stimuli obtained by downloading from various web sites, sets of five attractive and unattractive male photos, and five attractive and unattractive female photos, were selected. The attractive photos were obtained from modeling and fashion websites and depicted European and Brazilian models and television actors. The unattractive photos were obtained from blogs and were amateur shots. Each set contained a mixture of head and head-and-torso shots, and both smiling and neutral expressions. Photos were cropped and edited to be 400 × 400 pixels, and were displayed at the same size as the Gabor discs in the center of the screen. Copies of all photos used in the experiment are available from the authors.
All photos were shown to 8 female raters, who were asked to rate the photos in terms of attractiveness on a 10-point scale (1 = extremely unattractive; 10 = extremely attractive). The photos were presented in one of two random orders. The average ratings for the attractive and unattractive male photos were 7.95 and 1.78, respectively, t(7) = 9.81, p < .001, and the ratings for the attractive and unattractive female photos were 8.23 and 2.68, t(7) = 8.30, p < .001. Upon debriefing, none of the raters or participants indicated that they recognized any of the individuals in the photographs.
Procedure
Participants were tested individually in a quiet room. They were seated approximately 60 cm from the computer. They were told that they would see sequences of five stimuli on the screen, and that their task was to reproduce the duration of the fifth stimulus in each stimulus by pressing and holding down the left mouse button. They were asked to be as careful and accurate as possible, and completed three practice trials in which all stimuli were Gabor discs before starting the experimental trials.
The experimental session consisted of two blocks of trials, separated by a 10-min interval in which participants completed a filler task. The procedure is diagrammed in Figure 1. In one block of trials, the attractive and unattractive male photos were used as test stimuli, whereas in the other block of trials, the attractive and unattractive female photos were used as test stimuli. The order of blocks was counterbalanced across participants.

Diagram of experimental procedure
Each block consisted of 105 trials. On each trial, four Gabor discs were displayed for an identical duration, separated by an inter-stimulus interval (ISI), followed by a fifth stimulus of the same duration. On one third of trials, the fifth stimulus was either the same Gabor disc (neutral condition), an attractive photo (attractive condition), or an unattractive photo (unattractive condition). There were seven possible stimulus durations: 133 ms, 233 ms, 300 ms, 383 ms, 533 ms, 1050 ms, and 2100 ms, which occurred on an equal number of trials (pseudorandomly determined). These durations were the closest to those used by Tse et al. (2004; Experiment 6) that we were able to produce with our equipment, with the exception of the longest duration studied by Tse et al. (4200 ms). Thus there were five replications of each condition (neutral, attractive, unattractive) at each duration per block. The five attractive and unattractive photos were presented once at each duration. The ISIs within a trial varied randomly within +/− 10% of the stimulus duration. The purpose of the irregular stimulus spacing was to ensure that participants would respond to the duration of the stimuli, and not a beat or rhythm that might occur if the ISI were constant within a trial (Tse et al., 2004).
After the sequence of five stimuli was presented, the participants pressed the left mouse button to reproduce the duration of the fifth stimulus. The next trial began 2 s after they had made their response.
Results
We checked average response durations to screen for potential outliers. Data for one participant deviated by more than four standard deviations from the overall mean and were omitted. However, results were substantially unchanged in terms of statistical comparisons if this case was included (the only difference was that the three-way interaction was not significant; see below).
To test whether responding in the stimulus duration reproduction task conformed to Vierordt's Law (1868), in which shorter intervals are overestimated and longer intervals are underestimated (see Lejeune and Wearden, 2009 for review), we calculated the ratio of each estimated duration to the veridical duration, and averaged these ratios over blocks (male/female) and condition (neutral/attractive/unattractive) for each participant. Results are shown in Figure 2. Durations were overestimated for stimuli lasting less than 500 ms, and overestimated for stimuli lasting more than 500 ms, in accord with expectation based on Vierordt's Law.

Ratio of estimated duration to stimulus duration, averaged across test and control trials
Figure 3 shows average response durations for blocks with male photos as test stimuli (left panel) and blocks with female photos as test stimuli (right panel). Data were entered into a 2×3×7 repeated-measures analysis of variance (ANOVA) with male/female test photos, condition (neutral, attractive, and unattractive) and duration as within-subjects factors. There was a significant effect of condition, F(2,50) = 7.47, p < .01, partial η2 = .23. Post-hoc tests (Newman-Keuls) showed that average response duration for attractive and unattractive photos (Ms = 602 ms and 569 ms, respectively) were both significantly greater than the average response duration for neutral stimuli (526 ms), ps < .01 and .05, but did not significantly differ from each other (p = .10). This confirms the oddball effect, in that unexpected stimuli were judged as longer duration compared with the expected neutral stimuli. The oddball effect and Vierordt's Law (Figure 2) confirm expectations based on previous research with the stimulus duration reproduction task.

Average reproduced durations for blocks with male photos (left panel) and female photos (right panel) on test trials
There was a significant interaction between male/female test photos and condition, F(2,50) = 9.52, p < .001, partial η2 = .28. Post-hoc comparisons (Newman-Keuls) showed that attractive male photos were judged as having longer subjective durations (M = 633 ms) compared to both unattractive male photos (M = 554 ms) and neutral stimuli (M = 529 ms), both ps < .001, while unattractive male photos and neutral stimuli did not differ significantly (p = .11). In contrast, attractive female photos (M = 572 ms) and unattractive female photos (M = 584 ms) did not significantly differ (p = .43), while both were judged as significantly longer than neutral stimuli (M = 522 ms), ps < .01. This confirms the primary hypothesis of the experiment, because it shows that attractive vs. unattractive male, but not female, photos resulted in different estimated durations.
The condition × duration interaction was also significant, F(12,300) = 5.71, p < .001, partial η2 = .19. Post-hoc analyses (Newman-Keuls) showed that the effect of condition was only significant at the three longest durations (533 ms, 1050 ms, and 2100 ms). Thus the oddball effect was obtained primarily at the longest durations. As expected, the effect of duration was significant, F(6,150) = 78.86, p < .001, partial η2 = .76.
The three-way interaction between male/female photos, condition and duration was significant, F(12,300) = 1.80, p < .05, partial η2 = .07. Post-hoc analyses (Newman-Keuls) showed that attractive male photos were judged as significantly longer (p < .001) than unattractive male photos for the two longest durations (1050 ms and 2100 ms), but were not significantly different for shorter durations (all ps > .45). In contrast, for female photos, there were no significant differences between attractive and unattractive photos for any duration (all ps > .30). None of the other main effects or interactions was significant.
Finally, we conducted an analysis to estimate the size of the subjective expansion of time – the oddball effect – produced by the different test stimuli. As expected based on previous results (Tse et al., 2004), the estimated durations for test stimuli were only significantly greater than those for neutral stimuli at the longest durations (500 ms, 1050 ms, 2100 ms). Thus for each participant, we calculated the ratio of the test stimuli (male-attractive, male-unattractive, female-attractive, and female-unattractive) and control stimuli estimated duration, and averaged across the three longest durations to yield an oddball effect for each type of test stimuli. Figure 4 shows the results. A repeated-measures ANOVA found a significant interaction between male/female and attractive/unattractive, F(1,25) = 5,83, p < .05, partial η2 = .19. Post-hoc comparisons (Newman-Keuls) showed that the average oddball effect for male-attractive photos (M = 1.37) was significantly greater than for male-unattractive photos (M = 1.17), whereas the oddball effect for attractive and unattractive female photos did not significantly differ (Ms = 1.31 and 1.30, respectively). Single sample t tests confirmed that the oddball effect was significantly greater than 1 for male-attractive, male-unattractive, and female-attractive photos (ps < .05) and approached significance for female-unattractive photos (p = .05).
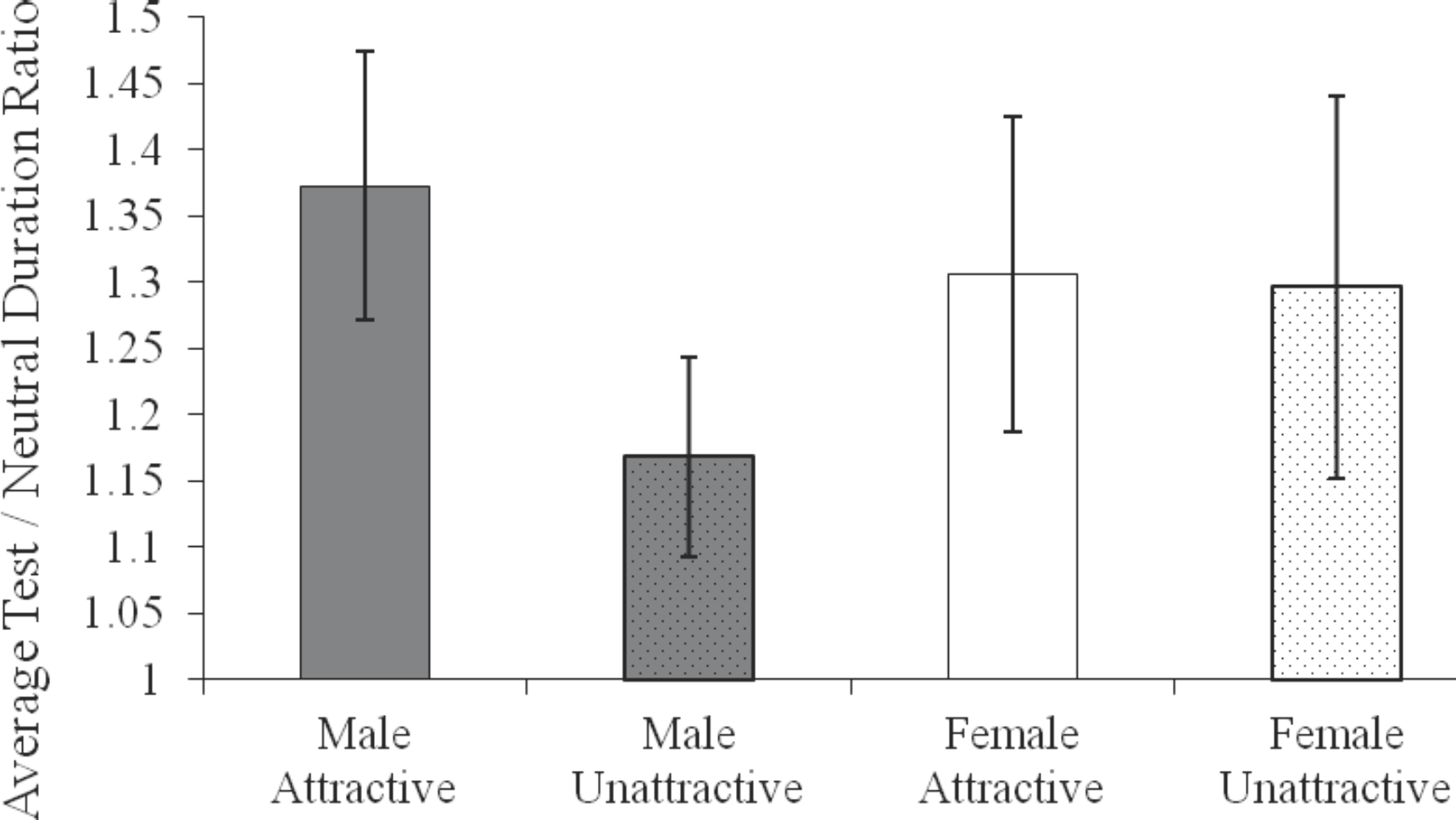
Estimates of the oddball effect obtained on test trials with attractive male photos, unattractive male photos, attractive female photos, and unattractive female photos
Discussion
We asked whether females' estimated duration of unexpected, briefly-viewed opposite-sex and same-sex photos would depend on attractiveness. Based on an evolutionary rationale, we predicted that attractive male photos would be judged longer than unattractive male photos, but that there would be no corresponding difference for female photos. This prediction was confirmed in an oddball task in which participants viewed series of five stimuli of equal duration and had to reproduce the duration of the last stimulus in each series by pressing a mouse button. The first four stimuli on each trial were identical, circular sine-wave gratings, and the fifth stimulus could be another identical sine-wave grating (on control trials), or a photo of an attractive or unattractive male or female (on test trials). Results showed that reproduced durations for attractive versus unattractive photos were significantly longer for males, but not females.
Our results suggest that attractiveness increases females' subjective duration of briefly-viewed male, but not female, photos. Because attractiveness is an important signal for the quality of a potential mate (e.g., Buss, 1989; Feingold, 1990, 1992; Gutierres, Kenrick, and Partch, 1999), it is reasonable to expect that for females, the unexpected sight of a highly attractive male would be a biologically significant event and engage appetitive motivational systems (Lang and Bradley, 2010). An adaptation to the timing system which allowed subjective time to expand when seeing an attractive potential mate would increase their salience and allow more information to be processed quickly, raising the likelihood that their mate value would be recognized. Such an adaptation would have had a selective advantage in ancestral environments.
There is considerable evidence that emotional events can alter the experience of time, particularly by engaging motivational systems related to defense (Lang and Bradley, 2010; see Droit-Volet and Meck, 2007, for review). For example, in a study using the International Affective Picture System (Lang et al., 2005), Angrilli, Cherubini, Pavese, and Manfredini (1997) found that participants' duration estimates of photos viewed for 2 s, 4 s, or 6 s showed an interaction between arousal and affective valence: For high arousal/negative valence photos (e.g., a severe head injury), duration was overestimated, while for high arousal/positive valence photos (e.g., erotica), duration was underestimated. The opposite result was obtained for low arousal photos: Durations of low arousal/negative valence (e.g., dead cow) were underestimated while those for low arousal/positive valence (e.g., pet dogs) were overestimated. Angrilli et al. (1997) suggested that the interaction between arousal and affective valence implied that there were two different mechanisms linked to timing that depended on arousal: An automatic mechanism for high arousal that was linked to defensive, motivational-survival mechanisms, and a controlled-attention mechanism associated with low arousal. Interestingly, Angrilli et al. found that the increased estimates of high-arousal/negative valence photos were only obtained for the 2-s duration. This suggests that the activity of the automatic/high-arousal mechanism was relatively brief.
Droit-Volet and colleagues have reported a series of studies that show that the emotional expression can alter perceived duration. For example, Droit-Volet, Brunot, and Niedenthal (2004) used a bisection task in which participants were first trained to discriminate between a pink oval that was displayed for either 400 ms (short) or 1600 ms (long). In the test phase, they were shown photos of female faces with either neutral or emotional expressions (angry, happy, sad) for intermediate durations, and required to classify these as short or long. Droit-Volet et al. found that compared to neutral faces, angry faces produced a significant leftward shift in the psychometric function, suggesting that the duration of angry faces was relatively overestimated. Psychometric functions for happy and sad faces also showed a leftward shift, but not as large and results approached but did not reach statistical significance.
Our study is the first demonstration that a stimulus attribute related to reproductive fitness and which engages the appetitive motivational system – a photo of an attractive opposite-sex person – can also increase estimates of duration. Presumably the greater reproduced durations for attractive male photos was due to an increased pacemaker rate associated with arousal, which is the same mechanism proposed for increased subjective duration for negative or threatening events associated with high arousal which engage the defensive motivational system (e.g., Droit-Volet, Mermillod, Cocenas-Silva, and Gil, 2010). According to the pacemaker-accumulator model of interval timing, arousal leads to an increased pacemaker, resulting in a larger number of pulses being stored in the accumulator during the stimulus (Penton-Voak, Edwards, Percival, and Wearden, 1996; Triesman, Faulkner, Naish, and Brogan, 1990; Wearden, Edwards, Fakhri, and Percival, 1998). When the duration is later reproduced, the greater number of pulses translates into an increased estimated duration. Our results suggest that the pacemaker rate can also be increased by activation of the appetitive motivational system.
Our results contrast with those of a recent report by Ogden (accepted). In Ogden's study, female participants judged the duration of photos of female faces that varied in attractiveness in a verbal estimation (VE) procedure. Ogden found that duration estimates for unattractive faces were significantly shorter than for attractive or neutral faces, particularly for longer durations. Interpreting her results in terms of SET, she suggested that unattractive faces captured more attention, resulting in fewer pulses stored in the accumulator and thus a shorter judged duration. Her results are similar to those of Gil, Rousset, and Droit-Volet (2009), who found using a bisection task that duration estimates for disliked foods (e.g., blood sausage) were judged as shorter than corresponding estimates for liked foods (e.g., dark chocolate).
At issue is why Ogden (accepted) observed a decrease in duration estimates for unattractive faces, whereas we found an increase for attractive ones. The most likely explanation is that Ogden's results were due to attention but ours to arousal. There are several differences between the studies that support this view. First, Ogden used only female photos, whereas we used both males and females and observed a significant effect of attractiveness only for male photos. Second, effects of arousal are believed to be transient, and whether they are observed can depend on procedural factors, particularly the amount of time between the stimulus-to-be-timed and the participant's response. For example, Gil and Droit-Volet (2011) studied effects of angry faces on time perception (which had previously been shown to increase duration estimates by Droit-Volet, Brunot, and Niedenthal, 2004) using five different procedures. They found that angry faces were judged as lasting longer than neutral faces when temporal bisection, verbal estimation, and production tasks were used, but for temporal generalization and reproduction tasks there were no significant differences between angry and neutral faces. In the temporal bisection, verbal estimation, and production tasks the participant could respond immediately after the stimulus-to-be-timed was presented. However, in the temporal generalization task the participant had to compare the judged duration with a ‘standard’ that had been presented at the beginning of the trial block, and in the reproduction task the word ‘reproduction’ was displayed on the screen for 500 ms before the participant could respond. Thus, Gil and Droit-Volet's results suggest that arousal effects are most likely to be observed when the task allows the participant to respond immediately after the stimulus is presented and does not require comparison with a previous standard. Similarly, in our procedure the participants could reproduce the duration immediately after the fifth stimulus in each sequence was presented. By contrast, in Ogden's study the participants reported the estimated duration (using the number keypad) following a delay that varied randomly between 1000 and 1500 ms after the stimulus had ended. Thus our procedure was more likely to be sensitive to arousal effects.
As we were adapting the stimulus duration reproduction (SDR) oddball task for use in the present experiment, it is important to consider our results in relation to those obtained in previous psychophysical research using similar procedures. A well-established finding in SDR tasks is Vierordt's Law: Relatively shorter durations are overestimated while relatively longer durations are underestimated. Tse et al. (2004) showed that their SDR data (Experiment 6) conformed to Vierordt's Law, and our results (Figure 2) were similar.
Another important question is how the size of the oddball effect – the increase in estimated duration for longer stimuli (500 ms, 1050 ms, 2100 ms) – compares to those in previous research. The oddball effect is measured as the ratio of the estimated to actual duration. Tse et al. (2004) reported ratios of approximately 1.4–1.5 using the method of constant stimuli (Experiments 1–4) and about 1.2–1.3 using magnitude estimation (Experiment 5) and the SDR task (Experiment 6). Although Tse et al. argued that the method of constant stimuli was more precise, subsequent research by Ulrich, Nitschke, and Rammsayer (2006) and Seifried and Ulrich (2010) showed that Tse et al.'s larger estimates of the oddball effect were likely inflated due to using an asymmetrical distribution of test stimuli. With symmetrical distributions and using both adaptive and non-adaptive methods for obtaining responses, Seifried and Ulrich concluded that the oddball effect was robust with a ratio between 1.1 and 1.3.
Our results were consistent with these prior studies in terms of the magnitude of the oddball effect. Our estimates ranged from 1.37 for male-attractive photos to 1.17 for male-unattractive photos, and were intermediate for attractive and unattractive female photos. Taken together, that our data were consistent with expectations based on previous SDR research (Vierordt's Law) and the range of oddball effects that we obtained, suggests that our procedure was a valid adaptation of the oddball task.
An alternative interpretation of our results should be considered. To the extent that the unattractive male photos might have been aversive, perhaps due to contrast resulting from the attractive male photos being presented in the same trial block, participants may have produced briefer durations because recalling the unattractive photos was unpleasant. However, it is important to note that a significant oddball effect was still obtained for the unattractive male photos – their reproduced duration was longer than that of neutral stimuli. If the prior argument were true and participants produced shorter durations simply because they found recalling the unattractive photos to be unpleasant, then the oddball effect should not have been obtained. Reproduced durations for stimuli greater than 500 ms should have been shorter than for neutral stimuli, but they were not. Thus we do not think it likely that the different reproduced durations for attractive and unattractive male photos resulted because participants found the latter aversive.
We have discussed our results as being due to arousal, but whether attention might have played a role as well should be considered. Tse et al. (2004) proposed that the oddball effect occurred because the low-probability stimulus captured more attention, but Ulrich et al. (2006) suggested that oddballs briefly increased the level of arousal, which in turn speeded up the pacemaker, leading to longer perceived duration. According to the standard information-processing model of timing, changes in attention can modulate perceived duration in different ways depending on the direction of attention. If attention is directed away from the stimulus to be timed (e.g., a distractor task), then perceived duration is shortened (Block, Hancock, and Zakay, 2010), which is interpreted in the model as pacemaker counts being lost prior to storage in the accumulator. However if attention toward the stimulus is increased, then perceived duration becomes longer. Understanding the potentially separate roles of attention and arousal in modulating time perception in situations relevant to reproductive fitness (e.g., seeing an attractive opposite-sex person) is an important task for future research.
One puzzle is the apparent discrepancy between our results and those of Maner et al. (2003): Why would females' estimated duration depend on attractiveness for male photos but not female photos in our study, whereas Maner et al. found that females were more likely to overestimate the frequency of attractive females but not males in a photo array when their processing time was limited? A possible resolution might be that the responses in the two studies were sensitive to different mechanisms. In our study, the unexpected brief viewing of a highly-attractive male presumably stimulated the appetitive system and increased arousal, which affected durations produced immediately afterwards. In Maner et al.'s study the participants viewed an array of 15 male (or female) faces and then were given a questionnaire, which included a question about the frequency of attractive faces amongst various distractor items. Maner et al. do not indicate how long participants were given for the questionnaire task, but it is likely that any arousal effects (which might have lasted only last a few seconds) would have dissipated. Thus, Maner et al.'s results are better explained in terms of attentional processes rather than arousal.
Some limitations of the present study should be acknowledged. By manipulating attractiveness within blocks of trials but manipulating male/female photos between blocks of trials, we used a design that was appropriate for the question we wanted to ask, that is, whether the effect of attractiveness was different for male photos vs. female photos. However this design limited our ability to address other questions, for example whether attractive male and female photos would differ in terms of duration estimates. An additional limitation is that we only used female participants. It would be very interesting to see whether males would show a similar effect in which attractiveness would modulate duration estimates of female, but not male, photos. Another worthwhile question for future research is whether results for females would change with the hormonal cycle, that is, whether there would there be a stronger effect of male attractiveness when female observers were at peak fertility.
Overall, results of the present study show that for female observers, attractiveness modulates the duration estimates of briefly-viewed male but not female photos. Our findings are consistent with the hypothesis that the timing system contains adaptations which provide flexibility in situations related to reproductive fitness.
Footnotes
Acknowledgements
The first author was supported by a fellowship from the Portuguese Foundation for Science and Technology (FCT). We thank Randolph Grace for helpful comments on an earlier draft. Address correspondence to Joana Arantes at the Department of Basic Psychology, School of Psychology, University of Minho, Campus de Gualtar, 4710-057 Braga – Portugal. Email: