
Abstract
Sex differences in mortality rates stem from a complex set of genetic, physiological, psychological, and social causes whose influences and interconnections are best understood in an integrative evolutionary life history framework. Although there are multiple levels of mechanisms contributing to sex based disparities in mortality rates, the intensity of male mating competition in a population may have a crucial role in shaping the level of excess male mortality. The degree of variation and skew in male reproductive success may shape the intensity of male mating competition, leading to riskier behavioral and physiological strategies. This study examines three socio-demographic factors related to variation in human male reproductive success; polygyny, economic inequality, and the population ratio of reproductively viable men to women across nations with available data. The degrees of economic inequality and polygyny explained unique portions in the sex difference in mortality rates, these predictors accounted for 53% of the variance. The population ratio of reproductively viable men to women did not explain any additional variance. These results demonstrate the association between social conditions and health outcomes in modern nations, as well as the power of an evolutionary life history framework for understanding important social issues.
Introduction
Sex differences are usually shaped by the processes of sexual selection; intersexual selection and intrasexual competition. Traditional explanations of sex differences in human mortality rates are based only on proximate mechanistic factors (e.g., Rogers, Hummer, and Nam, 2000). In recent decades, evolutionary researchers proposed that these differences result from the interaction of genetic heritage shaped by sexual selection with conditions in the social and developmental environment (e.g., Wilson and Daly, 1993). Sex differences in human mortality rates arise from genetic, physiological, behavioral, and social causes that are best understood when integrated in an evolutionary life history framework (Kruger and Nesse, 2004, 2006).
Males of most species allocate less effort to parental investment than females. Females are more discriminating in mate choice because of their greater costs in reproduction. Thus, male reproductive success is strongly related to the ability to compete for mating opportunities, whether by winning fights with other males, competing for social status or territory, or by presenting displays preferred by females (Darwin, 1871; Trivers, 1972). Males who succeed in mating competitions have more offspring, and this shapes traits that enhance mating outcomes. This occurs even for traits that may also lead to physiological and behavioral differences that make males more prone to injury, sickness and early death.
Potentially lethal violence is a facet of human male mating competition. This includes both within and between group conflicts (Chagnon, 1988). Violence may result from competition over access to and control of resources, as well as position in the status hierarchy (Buss and Shackelford, 1997). Men can successfully use violence to elevate their social status and gain respect from others (Campbell, 1993; Chagnon, 1992; Hill and Hurtado, 1996). Greater male mortality rates also arise from greater non-violent risk taking, leading to higher numbers of accidents, especially in young adulthood when males are entering into mating competition (Kruger and Nesse, 2004, 2006).
Sex differences shaped by sexual selection interact with the environment to yield a pattern with some consistency, but also with expected variations due to differential environmental conditions (Kruger and Nesse, 2006, 2007). The relationship between male mortality rates and factors related to the shape of the distribution of male reproductive success across species is likely mirrored by variation within our own species. This paper examines the relationship between three socio-demographic factors related to the degree of variation and skew in male reproductive success and the degree of excess male mortality, the degree to which male mortality rates exceed those for females in the same population. Male mortality rates are expected to be higher in nations where indicators of social conditions suggest a higher intensity of male mating competition.
Polygyny
Polygyny is the most common mammalian mating system, probably because of the prevalence of female specialization in infant nutritional provisioning and care and male specialization in mating effort (Low, 2003, 2007; Reichard and Boesch, 2003). In highly polygynous species, a few males will have many offspring while many others will have none. This creates powerful selection pressure for traits that lead to success in mating competition. The results include elaborate ornaments (such as the peacock's tail) and armaments (such as a deer's antlers), all with substantial costs. Recently, Clutton-Brock and Isvaran (2007) have shown that across vertebrate species, the longevity gap between males and females is highest among polygynous species. An archetypal example of polygyny is the elephant seal, where the disproportionately large males compete intensely for large harems of females and most males die before reproducing. The variance in estimated lifetime reproductive success among males is over four times that of females (Le Boeuf and Reiter, 1988).
The intense male competition for reproductive access in polygynous species selects for strategies that enhance competitive success at the cost of longevity (Kirkwood and Rose, 1991; Stearns, 1992, Williams, 1957). Across species, higher degrees of polygyny correspond with greater male-male competition and risky male behavior (Plavcan, 2000; Plavcan and van Schaik, 1997; Plavcan, van Schaik, and Kappeler, 1995), larger size and armor of males, and exaggerated male mortality rates compared to females (Leutenegger and Kelly, 1977). There is a strong correlation between sexual size dimorphism and excess male mortality across mammalian taxa, especially when accounting for phylogenetic constraints (Promislow, 1992).
Humans are far less polygynous than most other primates, but the variation in male reproductive success is still substantially higher than that for females. A few males gain a disproportionately high number of matings, creating a positively skewed distribution of male reproductive success that makes mating competition a potent selection force in humans (Betzig, 1986). Biologists note that increased sexual dimorphism, where males are larger than females, is directly and positively related to the level of male mating competition (see Bribiescas, 2006). Human females are on average 80% as large as males (Clutton-Brock, 1985), suggesting humans are at least mildly polygynous. Polygyny occurs in the vast majority of cultures (84%) documented by anthropologists (Ember, Ember, and Low, 2007).
Human populations vary in their degree of polygyny, corresponding with factors such as high male mortality in war and high pathogen stress (Ember et al., 2007). Polygyny is most abundant where males are heterogeneous in the quality of their territories and in human societies when there is substantial inequality in resources and social status (Borgerhoff Mulder, 1990; Orians, 1969). The degree of polygyny likely reflects the intensity of male mating competition. Higher levels of competition would be associated with higher male mortality rates.
Socio-economic inequality
Sexual selection helps to explain some sex differences in psychology and behavior, including stronger male tendencies for risk-taking, competitiveness, and sensitivity to social hierarchy (Cronin, 1991). Human male parental investment is relatively high amongst mammals (especially primates), and paternal investment in offspring may enhance offspring survival (Hill and Hurtado, 1996) and reproductive success (Geary, 2005). Thus women prefer males with high socio-economic status as marriage partners, as these men have an abundance of acquired resources and high potential for investment (Buss, 1989). Measures of male social status and economic power directly relate to reproductive success across a wide variety of societies (see Hopcroft, 2006). During recent human evolution, males who did not have substantial resources or status may have been unable to establish long-term relationships. Over the course of human genetic and cultural co-evolution, variance in male wealth and power increased through sociopolitical arrangements and intergenerational transfer (Smuts, 1995).
Where there is greater variation and skew in male social status and resource control, there will be greater competition for positions of power and status, leading to higher male mortality rates. Relative socioeconomic position has a stronger influence on mortality rates for males than for females (Bopp and Minder, 2003; Kruger and Nesse, 2006; Martikainen, Makela, Koskinen, and Valkonene, 2001). Increases in the uncertainly of economic opportunities and variability in males social status and resource control within populations are also associated with elevated male mortality rates (Kruger and Nesse, 2007). Thus, the degree of the economic disparity between the elite and impoverished may be directly related to the degree of excess male mortality across human populations.
Operational sex ratio
The operational sex ratio (OSR) was originally defined as the average ratio of sexually active males to sexually receptive females in a population (Emlen and Oring, 1977). Across species, a male biased OSR is associated with polyandry and a female biased OSR is associated with polygyny (Emlen and Oring, 1977). Darwin (1871) realized that the sex ratio of a species was usually even between males and females, the influential factors he described were later incorporated into the formal mathematical models of Düsing (1884) and Fisher (1930). This numerical equilibrium occurs on an evolutionary time scale, specific human populations may have imbalanced sex ratios (Darwin, 1871).
Fisher (1958) noted that the rare sex is more valuable in any marriage market and is able to be more selective of partners. The more plentiful sex faces greater intrasexual competition in securing a potential mate. When the OSR is male biased, available men outnumber available women and the enhanced power of female choice increases the level of male social status and resource provisioning potential necessary for obtaining partners (Pederson, 1991). In these populations, men with lower socio-economic status have an especially difficult time getting married (Pollet and Nettle, 2007). The degree of male bias in a population may be directly related to the intensity of male competition for social status, resources, and reproductive partners. Thus, populations with higher OSRs may also exhibit higher Male: Female Mortality Ratios (M:F MRs). In human populations, the OSR may be operationalized as the ratio of reproductively viable males to reproductively viable females in a population.
Hypotheses
This study examines the relationship between three socio-demographic factors that may be related to the degree of variation and skew in male reproductive success. Each factor comprises a unique aspect presumably related to the intensity of male mating competition. Consistent with patterns observed across species, the degree of excess male mortality in humans should follow the intensity of male mating competition.
Hypothesis 1: The prevalence of polygyny will be directly associated with the Male: Female Mortality Ratio (M:F MR).
Hypothesis 2: The degree of economic inequality will be directly associated with the M:F MR.
Hypothesis 3: The degree of male bias (relative female scarcity) in a population will be directly associated with the M:F MR.
Method
Kanazawa and Still (1999) created a polygyny index based on anthropological classifications of cultural groups within nations weighted by their proportion in the nation's population. The cultural group classifications are taken from the Encyclopedia of World Cultures (Levinson 1991–1995) and include: 0 = monogamy is the rule and is widespread; 1 = monogamy is the rule but some polygyny occurs; 2 = polygyny is the rule or cultural ideal but is limited in practice; and 3 = polygyny is the rule and is widespread. This index provides summary scores for 101 nations, 84 of which are also represented in the World Health Organization's Mortality Database (http://www3.who.int/whosis), used to calculate the Male: Female Mortality Ratio (M:F MR).
The standard measure for the degree of economic inequality in a population is the Gini coefficient (Gini, 1921), which is based on the degree of departure of the Lorenz curve representing the proportion of the total income by the proportion of income cumulatively earned by the bottom x% of the population from the line representing total income equality. Gini coefficients for most nations are available from the USA's Central Intelligence Agency's World Factbook (https://www.cia.gov/library/publications/the-world-factbook/).
The Operational Sex Ratio (OSR) was calculated as the ratio of males to females in a nation in the 15–44 year age range. These data are available from the United Nations' Demographic Yearbook (http://unstats.un.org/unsd/demographic/products/dyb/dyb2.htm). Data for polygyny, economic inequality, the OSR, and mortality rates are simultaneously available for 70 countries. The predictors were allowed to enter stepwise in a linear regression model to determine if they each make a unique contribution to the variance in the degree of excess male mortality across nations.
Results
The degree of economic inequality and degree to which polygyny was widespread were directly and significantly related to the Male: Female Mortality Ratio (M:F MR, see Table 1). These two predictors were also related to each other, sharing 17% of their variance. The ratio of males to females in a nation in the 15–44 year age range was not significantly related to the Male: Female Mortality Ratio and was inversely related to the degree of economic inequality. The bivariate distributions of nations are represented in Figures 1 and 2; the predicted relationships for economic inequality and polygyny were not driven by outliers. There were many nations where monogamy is the rule and is widespread.
Correlations among the Male: Female Mortality Ratio (M:F MR) and factors presumably related to male mating competition.

N = 70
p < .05
p < .001.

National M:F MR by economic inequality.

National M:F MR by degree of polygyny.
Results from stepwise linear regression predicting the Male: Female Mortality Ratio (M:F MR) across nations.
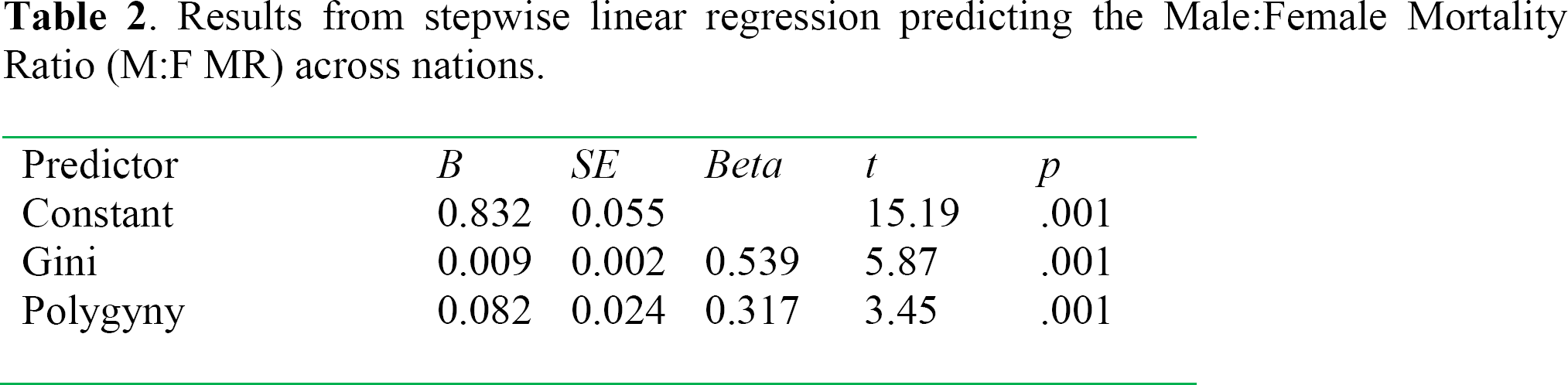
Results from the stepwise linear regression indicated that the degree of economic inequality and degree to which polygyny was widespread each made unique predictions of the degree of excess male mortality. These two predictors collectively accounted for 53% of the variance in sex differences in mortality rates across nations.
Discussion
Study results support the argument that socio-demographic factors arguably related to the intensity of male mating competition influence the extent to which male mortality rates are elevated above female mortality rates. The degree of polygyny directly represents the level of variation and skew in male mating success, differentials in male socio-economic status are associated with reproductive success and larger differentials may intensify male competition for partners and raise the standards (including those related to resource provisioning) for which men are considered suitable for marriage. These findings replicate similar associations seen across species between male mortality patterns and variance in reproductive success. Results did not support a relationship between excess male mortality and a relative abundance of men in the population.
Although the causal framework for excess male mortality incorporates multiple interacting levels of complexity from genetics to population parameters, the intensity of male mating competition may be the most crucial factor influencing the variation observed. Although polygyny and economic inequality are defined independently and explain unique portions of the variance in the degree of excess male mortality, they likely co-vary as differentials in male socio-economic status and resource control underlie many polygynous human mating systems. Future studies could examine meditational pathways between social and environmental conditions, physiology and molecular biomarkers, psychology and behavior, and mortality incidence.
It is possible that the operational sex ratio (OSR) is in part an artifact of higher male mortality rates. In a cross-national comparison, nations with relatively high mortality rates would see a substantial demographic impact of excess male mortality. In effect, excess male mortality would decrease the OSR, thus attenuating this relationship. Within nations with relatively lower mortality rates, localized OSRs have exhibit the predicted relationship with the M:F MR. In the USA, where regional differences in the OSR are largely driven by economic migration (Gwin, 2007), the OSR has a direct positive relationship with the level of excess male mortality (Kruger and Nesse, 2005).
These analyses are limited by the currently availability of data. The indicators are rather crude compared to more detailed and/or sophisticated measurements, which may not be available at this time. The polygyny indicator is ordinal at best, as it is based on classifications of the degree to which polygyny is accepted and considered widespread, rather than the actual degree of variation and skew in quantitative assessments of men's access to mating partners. Population estimates many not be accurate, and even when valid census counts are available these numbers may not truly represent the numbers of men and women who are active in the mating market.
Despite these limitations, the magnitude of associations are substantial, suggesting that even crude and approximate indicators are sufficient to reveal the hypothesized relationships. The strength of the association between socio-economic factors and mortality patterns would likely be considerably stronger if more accurate and precise quantitative representations of these constructs were available. Replications of these results will be likely when additional data becomes available.
The psychological salience and perception of the economic gradient may mediate the relationships with behavior, health, and mortality. Assessments of attitudes and perceptions could reveal the accuracy of popular beliefs. Market driven mass media may foster unrealistic expectations for the level of socio-economic achievement necessary for reaching common goals such as marital success and raising a family.
The relationship between social inequality and adverse health outcomes, even for those in upper socio-economic ranges, is increasingly recognized (e.g., Wilkinson and Pickett, 2009). The dramatic social and health benefits associated with relative egalitarian societies (see Wilkinson and Pickett, 2009) may be substantial incentives for social leveling. However, despite the potential benefits of economic leveling interventions, any effort to substantially reduce variations in wealth and resource control will likely face considerable political opposition. Paradoxically, opposition to such redistributions will be especially prevalent from men. Due to the long association of male status and reproductive success in our evolutionary history, men are both more sensitive to their position in the social hierarchy as well as to perceived threats to their relative status. The fragility of socialist utopias such as the Paris Commune of 1871 and other communities intentionally suppressing status differentials reveals the difficulty in sustainably implementing such social structures. Edward O. Wilson once remarked that “Karl Marx was right, socialism works; it is just that he had the wrong species” (Novacek, 2001). In sum, this study contributes to the growing body of literature demonstrating the substantial benefits that the evolutionary framework offers for understanding social patterns and important social issues.