
Abstract
To answer the question of whether sensory pleasure exists in birds, I trained an African-gray parrot (Psittacus erythacus) named Aristote to speak. Stage 1 of the study consisted in gaining Aristote's affection. In Stage 2 Aristote was taught to speak, following Irene Pepperberg's triangular method: another person and I would talk together and look at Aristote only when it used understandable French words. Thus Aristote learned to say a few words for obtaining toys or getting my attention; e.g. “donne bouchon” (give cork) or “donne gratte” (give scratch/tickle), with the appropriate reward. In Stage 3, the word bon (good) was added to the short list of words used by Aristote. I said “bon” when giving Aristote the stimuli it requested and which would, presumably, be pleasurable; e.g. gratte bon. Aristote started to use short sentences such as “yaourt bon” (good yogurt). Eventually, Aristote transferred the word bon to new stimuli such as raisin (grape), an association I myself had never made. Such a use of vocabulary, and moreover its transfer, likely shows that this bird experienced sensory pleasure.
Introduction
There is indeed in them (the animals) pleasure and pain III 11; the animal possesses senses not for being but for well-being…taste that is agreeable and distressing, in order for the animal to perceive these qualities in food, desire them and moves itself.
(Aristote, 284–322 Ante p. III 13)
In no case may we interpret an action as the outcome of the exercise of a higher psychological faculty, if it can be interpreted as the outcome of the exercise of one which stands lower in the psychological scale (Morgan, 1894 p.24)
Evolutionary psychologists argue that cognitive mechanisms, like anatomical and physiological traits, are shaped by natural selection (Bunge, 1979; Cosmides, Tooby, and Barkow, 1992). It is not known at what point in evolution sensory pleasure emerged. One way to test whether sensory pleasure is present in an animal is to look for taste aversion learning (Garcia, Kimerdorf, and Koelling, 1955; Revusky and Bedarf, 1967). Taste aversion takes place when ingestion associated with a new flavor is followed within a day by visceral illness, such as nausea or diarrhea. This is an adaptive learning mechanism to avoid food that contains toxic compounds (Garcia, Hankins, and Rusiniak, 1974; Kalat, 1977). In taste aversion learning, an initially appetizing conditioned stimulus (the flavor of a novel food) is paired with an unconditioned stimulus (a digestive illness) and thus becomes aversive. In humans, taste aversion learning changes the hedonic value of a flavor associated with a visceral illness. A flavor initially perceived as pleasant becomes unpleasant once aversion has been acquired (Bernstein and Borson, 1986; Bernstein and Webster, 1980; Berridge, 2000, 2001; Garb and Stunkard, 1974). Such a shift in the hedonic value of a stimulus is called alliesthesia and, in humans, is a conscious process (Cabanac, 1971). Taste aversion learning, a long-term example of taste alliesthesia, has been described in a variety of mammals: rats, coyotes, horses, bats, ferrets, and guinea pigs (Garcia, Hankins, and Rusiniak, 1974; Gustavson, Kelly, and Sweeney, 1976; Houpt, Zahorik, and Swartzman-Andert, 1990; Kalat, 1975; Rusiniak, Gustavson, Hankins, and Garcia, 1976; Terk and Green, 1980). It has also been shown in reptiles (Burghardt, Wilcoxon, and Czaplicki, 1973; Paradis and Cabanac, 2003; Terrick, Mumme, and Burghardt, 1995). Because birds and mammals share a common reptilian ancestry, it is highly likely that birds too experience sensory pleasure. Indeed, taste aversion learning, has also been shown to exist in quails and hawks (Brett, Hankins, and Garcia, 1976; Wilcoxon, Dragoin, and Kral, 1971). This makes it highly likely that birds too experience sensory pleasure.
The present study further explores whether birds are capable of sensory pleasure. Instead of indirectly measuring likes and dislikes from ingestive behaviors, I used Pepperberg's method to explore a bird's mind through verbal exchange. The aim was not to measure the bird's cognitive performance but rather to use its verbal response as a way to explore its capacity to experience pleasure. Therefore, no attempt was made to teach it a very broad vocabulary, but rather those words relevant to the aim of the study, i.e. to test whether the bird would express verbally its feeling of pleasure.
Materials and Methods
Subject
The experimental subject was an African-gray parrot (Psittacus erithacus) named Aristote (because of the Middle Ages saying: Aristoteles dixit!). It was bought from a pet shop and resided in my home. Most of the time the bird was free to stay on its stand equipped with feeders. Yet it did not like to stay alone and tended to follow the present person from room to room in our apartment and was moved to its cage only at night. To prevent fleeing the wing extreme feathers were trimmed on one side. According to the seller, the bird was approximately one year old at the time this experiment began. In Stage 1, the parrot was named and acclimatized to its environment.
Training Procedure
General procedure
The procedure followed closely the model/rival training procedure published by Pepperberg over the last twenty years and repeatedly used by her with her African-gray parrot, Alex, and its successors, (cf. Pepperberg, 1981, 1990, 2000) and will be only briefly described here. A triangular relation is established between the experimenter, a second person and the parrot (Fig. 1). The two humans exchange roles as trainer and trainee; the trainee produces the targeted response expected from the parrot and is the parrot's rival for the trainer's attention. Rewards for correct responses are the objects that trainer and trainee exchange through alternating requests. The trainee, or the parrot, is rewarded when he/it vocalizes the correct word for the reward (Pepperberg, 1988).
The training sessions took place at random when I returned home in the evenings and on weekends. The sessions were usually not very long, between 5 minutes and 1 hour depending on Aristote's level of attention (and the trainer's time table).
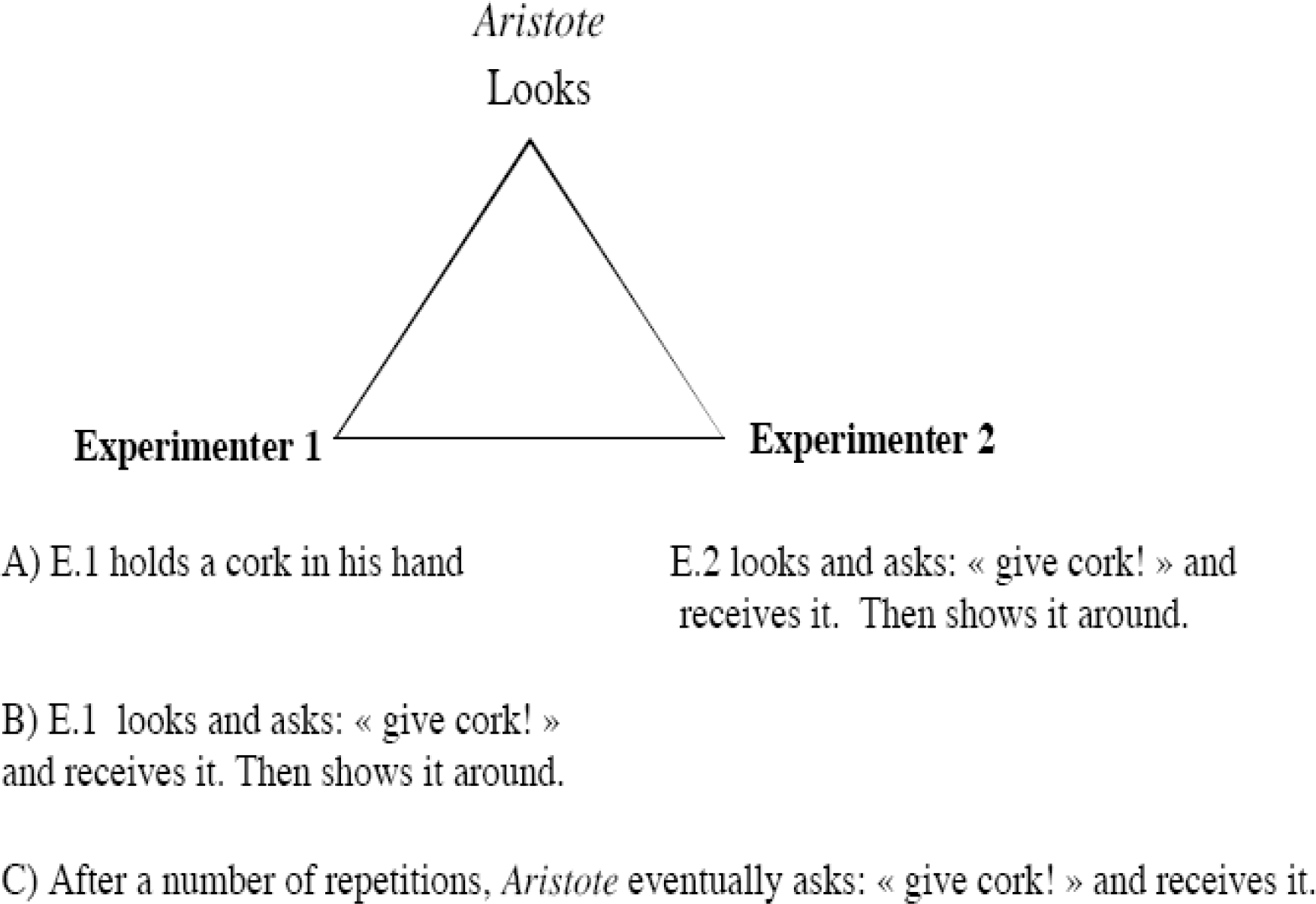
Pepperberg's method: the triangle of the model/rival procedure. E.1 and E.2: experimenters 1 and 2.
Training with pleasure vocabulary
After general training and acclimatization Stage 2 of the experiment began, Aristote was taught the needed vocabulary: names of stimuli that it would seek and that, presumably, gave it pleasure -mostly names of food items (seeds, grapes, apple, yogurt, etc.) but also toys (cork, etc.).
The aim being to explore hedonicity, no attempt was made to teach the bird a very broad vocabulary. Rather, the limited list of taught words focused on tastes and on mechanical stimuli (tickling). Yet, Aristote learned spontaneously all by itself additional words such as papa (daddy), maman (mommy), bonjour (good morning), au dodo (go to bed), that it would use appropriately when it wanted the person to come or when it woke up or wanted to sleep. For example it used to say spontaneously “hello” when the telephone rang. The delay between the attempt to teach a new word and Aristote's use of it was variable and not recorded. N.B. Aristote knew its name as well as those of the family members, but it never spoke its own name although it used the names of the various persons.
Once this vocabulary was acquired the Stage 3 began. The word bon (good) was added to Aristote's vocabulary, in association with the stimuli that presumably aroused pleasure. Use of the word bon by the bird was checked to see whether the word was correctly used. For this purpose, the word bon, was carefully never associated with the stimulus and/or the word raisin. It was expected that an eventual transfer of the word to this “pure” stimulus would ensure that Aristote used it as an expression of its own experience. By so doing, any use by Aristote of the word bon with the stimulus raisin would not simply be an imitation of the trainer's own behavior. It would be a mental association made by the bird itself. As a control, it was expected that Aristote would use the word bon only with stimuli that would, presumably, arouse sensory pleasure.
Results
Stage 1, gaining the bird's confidence, took several months. Over this time span Aristote developed a strong attachment to the experimenter, a condition described as favorable by Pepperberg because in the situation of rivalry the bird would be more likely/willing to seek its trainer's attention.
Stage 2 took place gradually over one year. Aristote learned:
- nouns: bouchon (cork), yaourt (yogurt), raisin (grape), orange (orange), graine (seed);
- commands: “donne!” (give), “gratte!” (scratch, tickle), “au dodo!” (go to bed);
- names of the various members of the household including papa and maman (daddy, mommy) and the children's names;
- and greetings such as “bonjour” (good morning), “bonne nuit” (good night), and “hello” when the telephone rang.
Aristote learned to make sentences by associating a command, mostly “donne,” with an object, e.g. “donne bouchon!” or “donne graine!”
In Stage 3, Aristote learned to add the word bon (good) to the short list of words it was using: yogurt, tickle, seeds, orange, etc. I said “bon” when Aristote obtained the stimulus it requested; e.g. “gratte bon” and Aristote started to use it also: e.g., “yaourt bon” (good yogurt) after it had obtained the stimulus. Finally, Aristote transferred the word bon to qualify any stimulus that apparently it liked, especially in the case of raisin (grape), an association I myself had carefully never made.
Discussion
The first problem when studying a single subject is that of replicability. A question often arises as to the replicability of our work [Pepperberg's] […] Although our studies, which use a single subject previously trained to work within a language paradigm, may actually be difficult to replicate in other laboratories, in principle such replication is by no means impossible. Moreover, the objective of this study (similar to the work on the one sea lion (Schusterman and Krieger, 1986) was to determine if a psittacine subject could learn a relative size discrimination. The implication of a positive finding, whatever the training methods or history of the subject, would be that this aptitude is within the capacity of the species. (Pepperberg and Brezinsky, 1991, p. 288)
Pepperberg's statement about her Alex's capacity to discriminate between relative sizes applies fully to Aristote's capacity to experience sensory pleasure. When the question addressed by a study is to find out whether a species can respond in a given way, a positive finding from a single representative of the species is sufficient to answer the question.
The ability of a parrot to generalize or generate novel combinations of labels is not in itself an original finding (Pepperberg, 2007) and can be seen as evidence for imitation; thus Aristote would spontaneously say au dodo (meaning “go to bed” in child language) in the evening, when presumably it wanted to return to its cage to sleep. Yet, Aristote's use of the word bon (good) in association with stimuli that it obviously sought verbally and behaviorally, and not in association with other hedonically indifferent words may be understood as a sign that the bird experienced sensory pleasure. Moreover, its transfer of the word to other stimuli that the experimenter had never used, corroborates this interpretation. Such a conclusion is in keeping with the existence of taste aversion learning in birds (Brett, Hankins, and Garcia, 1976; Wilcoxon, Dragoin, and Kral, 1971), as that phenomenon likely takes place in the hedonic dimension of consciousness.
It is quite remarkable that Aristote's use of the word bon took place not to obtain a reward, contrary to its use of the words donne raisin or donne gratte, but in absence of motivation. Yet, Aristote did not always express motivation when speaking, as when it said Hello, when the telephone rang. Therefore, bon was not an exception that would arouse suspicion that the word bon could have been used as something like “accept/take.” What made Aristote's use of the word bon more likely that it was told to express pleasure was that the word was used only rarely after an obviously sought stimulus had been given, and never randomly or by chance. Another indication of actual sensory pleasure was that bon was not limited to palatable sensations but was also told when Aristote's neck and head were gently tickled. Thus the word bon was used across other contexts and was not limited to taste responses as the use of good and bad in lexigram trained bonobos (Lyn, Franks, and Savage-Rumbaugh, 2008).
This capacity of a parrot to experience pleasure should be examined in the light of available evidence from comparative and evolutionary psychology (Bunge, 1979, 2003; Cosmides, Tooby, and Barkow, 1992). Phylogeny tells us that present-day reptiles, mammals, and birds share common reptilian ancestry. Therefore if a property or function exists in present-day reptiles, it should exist also in other vertebrates with which they share common ancestry, i.e. mammals and birds. Furthermore, because the fact that lacertilians also displayed taste aversion (Burghardt Wilcoxon, and Czaplicki, 1973; Paradis and Cabanac, 2004; Terrick Mumme, and Burghardt, 1995), but not amphibians (Paradis and Cabanac 2004), would suggest that present-day birds should have inherited their hedonic mental capacity from the early amniotes ancestral to them, to present-day lizards, and to mammals.
It is likely, therefore, that amniotic vertebrates—i.e. including birds—possess some mental capacity with, in it, sensory pleasure. This has two main implications:
Sensation and sensory pleasure take place in consciousness. Mammals, birds, and reptiles show autonomic signs of emotion that resemble those of humans. On the other hand, amphibians and fish show no signs of emotion. This suggests that consciousness emerged among early reptiles (Cabanac, 1999).
The existence of pleasure in birds, as well as in mammals (Berridge, 2001; Cabanac and Johnson, 1983; Conovert and Shizgal, 1994a, 1994b; Shizgal, 1997) and lizards (Balaskó and Cabanac, 1998) underlines its fundamental role in decision making as the optimizer of behavior in all amniotic vertebrates (Cabanac, 1971, 1992; Cabanac, Cabanac, and Parent in press). Indeed, the hedonic dimension of sensation in rats, could be experimentally somewhat dissociated from behavioral consumption of a stimulus (Wyvell and Berridge, 2000); such a dissociation would confirm that consciousness emerged in amniotic brains from the complex circuitry that had to be pre-existing. It remains that in intact brain conditions liking and wanting are associated and thus liking optimizes behavior.
Footnotes
Acknowledgements
I wish to thank my wife whose patience permitted this study at home.