
Abstract
Women have been reported to perform better with hand rather than arm movements (Sanders and Walsh, 2007) and with visual stimuli in near rather than far space (Sanders, Sinclair and Walsh, 2007). Men performed better with the arm and in far space. These reciprocal patterns of sex differences appear as Muscle*Sex and Space*Sex interactions. We investigated these claims using target cancellation tasks in which task difficulty was manipulated by varying target size or the number of distracters. In Study 1 we did not find the Muscle*Sex or the Space*Sex interaction. We argue that ballistic movement was too simple to reveal the Muscle*Sex interaction. However, a trend for the Space*Sex interaction suggested task difficulty was set too high. Study 2 introduced easier levels of difficulty and the overall Space*Sex interaction narrowly failed to reach significance (p = 0.051). In Study 3 the Space*Sex interaction was significant (p = 0.001). A review of the present, and four previously published, studies indicates that task selection is critical if the Space*Sex interaction and its associated reciprocal within-sex differences are to be demonstrated without the obscuring effects of Space and Difficulty. These sex differences are compatible with predictions from the hunter-gatherer hypothesis. Implications for two-visual-system-models are considered.
Keywords
Introduction
In a recent series of studies (Sanders and Perez, 2007; Sanders, Sinclair and Walsh, 2007; Sanders and Walsh, 2007) we focused on a possible evolutionary origin for present day sex differences in cognitive and motor performance. We started with the Hunter-Gatherer Hypothesis in which Silverman and Eals (1992) proposed that present day sex differences in spatial abilities arose from the division of labor associated with our ancestral hunter-gatherer mode of life. Of course, the division of labor between men and women extended beyond hunting and gathering per se. Skills required for hunting would also be needed for defense and attack while the close fine motor movements required for gathering would be needed for caring. Each of these tasks would have provided additional selection pressures for the differentiation of sex-dimorphic skills.
Silverman and Eals noted that the spatial tasks at which males are reported to excel, such as mental rotation, require individuals to orientate themselves with an object and to maintain that relationship during movement by performing mental transformations, skills that would aid hunting, especially in unknown territory. Consequently, they agued that women should have evolved spatial skills, such as object location memory, which would aid gathering, and they demonstrated a female advantage for such tasks (Silverman and Eals, 1992; Eals and Silverman, 1994). Further support comes from studies of navigation (e.g., Galea and Kimura, 1993) in which women tended to use landmarks (effective in the known areas traversed by gatherers) whereas men used distance and cardinal directions (effective in the unknown areas traversed by hunters).
Other evidence that was congruent with the Hunter-Gatherer Hypothesis came from reports of sex differences in manual dexterity, favoring women (Nickolson and Kimura, 1996; Sanders and Kadam, 2001) and in targeted throwing, favoring men (Watson and Kimura, 1991). Both tasks are ecologically valid but they did not point to a location for the sex differences because the studies confounded three variables: the visual space in which the tasks were performed, the muscles used to perform the tasks, and the cognitive demands of those tasks. To overcome this problem, we derived predictions from the Hunter-Gatherer Hypothesis by identifying motor skills (Sanders and Walsh, 2007) and visual skills (Sanders, Sinclair and Walsh, 2007) that would differentially support hunting and gathering and tested those predictions by devising tasks that avoided the previous confounds.
We argued that evolutionary selection for hunting would favor individuals with, among other things, the ability to visually locate appropriate prey in far (extrapersonal) space, then to aim and launch a projectile accurately at that distant target. Conversely, selection for gathering would favor individuals with, among other things, the ability to visually locate appropriate items in near (peripersonal) space, then to reach, grasp and retrieve these items efficiently. From these observations we derived directional within-sex predictions for men and women that were complementary and reciprocal. Men, as the predominant hunters, should be better at processing visual information from far than from near space and better when using the larger proximal muscles of the upper arm and shoulder than when using the smaller distal muscles of the wrist and fingers. On the other hand, as the predominant gatherers, women should be better at processing visual information from near than from far space and better when using the hand than when using arm.
It is important to note that we are not proposing a new theory as an alternative to the Hunter-Gatherer Hypothesis. However, we are deriving novel predictions from the ancestral division of labor between women and men which was highlighted by that hypothesis. Traditional predictions from the Hunter-Gatherer Hypothesis point to unitary between-sex differences, i.e. women will be better than men at some tasks and men better than women at other tasks. In contrast, we are making within-sex predictions: a woman will perform better with her hand than her arm and better with visual stimuli in near rather than far space while a man will show the reverse patterns. When testing our predictions, the traditional between-sex comparisons are not relevant because the relative superior performance of women or men will depend on the nature of the task chosen, i.e. whether it is female-favoring, male-favoring or sex-neutral. Our position will be supported by significant Muscle*Sex and Space*Sex interactions together with significant within-sex paired comparisons between hand and arm, and between near and far space that are in the predicted direction within each sex. Our underlying thinking is that a history of evolutionary selection for hunting in men and gathering in women has led to sex differences in the neural mechanisms supporting motor control and visual processing. If this thinking is correct then the reciprocal patterns of hand/arm and near/far performance that we have predicted for women and men should appear whatever tasks are used.
The definitions of near and far space come from early studies of radial visual neglect which pointed to a functional division of visual space into near (peripersonal) and far (extrapersonal) domains in the sagittal plane. Near and far space, were originally defined by Brain (1941) as “grasping distance” as opposed to “walking distance” and later by Brouchon, Joanette and Samson (1986) as “reaching field” and “pointing or throwing field”. Near space is typically taken to be 500 mm or less, while far is defined as 1000 mm or more. In our first investigation of visual processing (Sanders, Sinclair and Walsh, 2007) we conducted three studies to test our prediction from the Hunter-Gatherer Hypothesis that women would perform better when using visual information from near rather than far space, whereas men would perform better with information from far rather than near space.
Sanders-Sinclair-Walsh Study 1 used a time estimation task conducted via the Internet. Participants watched an image of a hovering toy UFO moving across a table top towards a docking station. The UFO disappeared short of its destination and participants indicated the moment they estimated it would have docked by pressing their space bar. Near and far virtual space conditions were created by having the UFO move above the front or the rear half of the table. Sanders-Sinclair-Walsh Studies 2 and 3 used puzzle completion tasks conducted in the laboratory in which participants saw their hands and a simple five-piece “jigsaw” puzzle as an image projected via a webcam onto a near monitor or a far screen. All three studies generated significant Space*Sex interactions, arising because, within-sex, women tended to perform better in the near condition and men in the far condition, but these within-sex differences between near and far performance varied. In Sanders-Sinclair-Walsh Study 3 women completed the puzzles significantly faster in the near than in the far condition while men were significantly faster in the far than in the near condition. However, this was not the case in the other two studies. In Sanders-Sinclair-Walsh Study 2, which used a more difficult version of the puzzle task, the near/far performance difference was significant for women but not for men. Conversely, in Sanders-Sinclair-Walsh Study 1, which used the time estimation task, the near/far performance difference was significant for men but not for women.
In contrast, a related report (Sanders and Perez, 2007) found no difference at all between women and men in the patterns of their performances on a visuomotor task conducted in near and far space. Participants were required to use either a short (near space) or long (far space) hooked metal stylus to move colored washers from a starting array to color-coded locations in a target array. The stylus and washers were manipulated either by movements of the wrist and fingers (hand condition) or by movements of the upper arm and shoulder (arm condition). These two conditions were included to test an earlier finding (Sanders and Walsh, 2007) of a Muscle*Sex interaction that confirmed the hand/arm prediction we had derived from the Hunter-Gatherer Hypothesis: women would perform better with their hand than with their arm while men would perform better with their arm than with their hand. Although the hand/arm within-sex differences were replicated, Sanders and Perez failed to find the near/far within-sex differences; an outcome that they attributed to the use of a tool to manipulate the washers (see General Discussion for an account of this argument).
Given these somewhat variable outcomes from our reported studies of potential within-sex differences in the processing of near and far space, we decided to further investigate this issue. In selecting an appropriate set of new tasks we paid particular attention to task difficulty as this appears to be a crucial factor in the revelation of sex differences (Sanders, Sjodin, and de Chastelaine, 2002). Indeed, the critical nature of this variable was seen in our study of hand and arm use (Sanders and Walsh, 2007) which used a computer-based tracking task with four levels of difficulty determined by target speed (slow and fast) and trajectory (simple or undulating circle). The Muscle*Sex interaction appeared only in the slow/complex condition, with women tracking significantly better with their hand than with their arm while men showed the reverse pattern. Consequently, for the present studies we chose a computer-based target cancellation tasks in which task difficulty could be varied by manipulating either target size or the number of distracters. The targets were presented either on a computer monitor (near space condition) or projected onto a wall-mounted screen (far space condition). Our primary interest was the reciprocal within-sex differences in the processing of visual information from near and far space for which we predicted a Space*Sex interaction with women performing better in near than far space and men better in far than near space. In addition, for the first of our three studies we also predicted a Muscle*Sex interaction with women performing better with their hand than arm and men better with their arm than hand.
STUDY 1
Study 1 was designed to investigate the possible occurrence of sex differences in two abilities: (a) the visual processing of near and far space; (b) the control of hand and arm muscles. We designed a computer-based target cancellation task. Participants used their preferred hand or arm to operate either a short (hand condition) or a long (arm condition) joystick to position a cursor over a target which they then cancelled by pressing the space bar with their non-preferred hand. Task difficulty was manipulated by varying target diameter.
Materials and Methods
Participants
Forty-eight participants, 24 women (mean age 26.33, SD 5.70) and 24 men (mean age 31.54, SD 14.04), were recruited as an opportunity sample from among our University students and staff. All of the participants were right handed, had normal or corrected to normal vision and all were na ï ve to the purpose of the study. None of the participants had sustained an injury to the right hand or arm within the previous twelve months. The study was approved by the Departmental Ethics Committee. All participants gave informed written consent and were aware that they could withdraw from the study at any time. None withdrew.
Tasks and procedure
We used a mixed design. Sex was a between-participants factor with two independent groups, women and men; Space, Muscle and Difficulty were within-participants factors with repeated measures on near/far, hand/arm, and five levels of difficulty. The computer-based study was run by a custom-written program that recorded the time taken by participants to move a cursor from a central starting position to locate and cancel circular targets that appeared randomly elsewhere on the screen. In the near condition the stimuli were presented on a 430 mm monitor placed 500 mm from the participant with the centre of the screen at eye-level. On this screen the diameter of the targets varied across level of difficulty as follows: Level 1, 70 mm; Level 2, 35 mm; Level 3, 22 mm; Level 4, 10 mm; Level 5, 5 mm. For the far condition the stimuli were projected 2.4 times larger onto a wall-mounted screen placed 3200 mm from the participant with the centre of the screen 600 mm above eye-level so that the display could be seen over the top of the monitor.
In the hand condition the forearm of the participants was restrained by strapping to the table and they moved the onscreen cursor by manipulating a short (70 mm) joystick with wrist and finger movements. For the arm condition the same joystick was moved from the table to the floor and its length extended to 1200 mm by attaching a rod. Participants were instructed to hold a 49 mm diameter ball at the top of the rod in the palm of their hand, to keep their wrist locked and to use their upper arm and shoulder muscles to move the cursor. The size of the ball and length of the rod encouraged, and the instructions ensured, that finger and wrist movements were effectively eliminated and that the extended joystick was manipulated by the proximal muscles of the upper arm and shoulder only. The maximal movement of the top of the joysticks in any direction from the central position, 42 mm for the short (hand condition) and 600 mm for the long (arm condition), produced the same 37.5 mm onscreen movement of the cursor and was sufficient to encompass all of the targets. Participants used the joy stick with their preferred hand or arm to move the onscreen cursor and they pressed the space bar with their non-preferred hand to start trials and cancel targets. The sequence of screens that constituted a trial is illustrated in Figure 1 and described below.

An example of the sequence of screen presentations (not drawn to scale) that were used for the target cancellation tasks in Studies 1 and 2 in which task difficulty was manipulated by varying target diameter. The cross represents the cursor, the square the starting position and the circle the target which was light grey in Study 1 but in Study 2 is was orange as shown in Figure 1c. Further explanation is presented in the text.
Trials began with the presentation of a starting position, a light grey square (12 × 17 mm in the near condition) at the centre of the screen (Figure 1a). Participants moved the cursor (a cross) onto the light grey square which turned dark grey to show it was activated (Figure 1b). Pressing the space bar at this point caused the dark grey square to disappear, timing to begin, and an appropriately sized target to appear elsewhere on the screen (Figure 1c). Participants were required to move the cursor as quickly as possible from its central starting position to a point over the target which turned dark grey (Figure 1d). Pressing the space bar at this point caused the target to disappear, timing to stop, and, following a 100 ms clear screen, the sequence returned to the starting position screen (Figure 1a). At the end of each block of five trials a clear screen was displayed for 5000ms to mark the change to the next level of difficulty.
The order of presentation of the four conditions, hand/near, hand/far, arm/near and arm/far, was counterbalanced across participants. Within each condition, each target size was presented as a block of five trials starting with the easier Level 1 and progressing sequentially to the more difficult Level 5. Targets could appear in any one of six different screen positions so that at each level of difficulty participants experienced targets presented at 5 of the 6 screen positions randomly selected by the computer. Response times, i.e. the time between cancelling the central square and cancelling the circular target, were recorded for each trial in ms. The median response times for the five trials in each condition were used for statistical analyses. Before starting the experiment, participants were given verbal instructions and, to ensure familiarity with the procedure, they completed 10 practice trials at Level 1, 5 in the near and 5 in the far space condition.
Results and Discussion
Initial analysis
Target cancellation times were submitted to a 4-way mixed ANOVA with Sex (women/men) as a between participants factor and Space (near/far), Muscle (hand/arm) and Difficulty (Levels 1 to 5) as within participants factors. Three of the four main effects were significant, Space, Muscle and Difficulty but not Sex (Table 1). Participants were faster in near than far space (F 1,46 = 4.781, p = 0.034), faster with the hand than the arm (F 1,46 = 16.302, p < 0.001), and faster with larger than smaller targets (F 1,46 = 323.094, p < 0.001).
Study 1: Mean (SEM) response times for men and women when using their hand or arm muscles to cancel targets presented in near or far space. Task difficulty was manipulated by varying the target size from large (Level 1) to small (Level 5). Significant main effects are high-lighted in bold color (

Further analysis
Apart from the interaction between Muscle and Difficulty (F 1,46 = 7.083, p = 0.001), which arose because responses with the hand were faster at all levels of target size except for the smallest, none of the other interactions was significant. However, we had argued that the predicted Muscle*Sex and Space*Sex interactions may appear only at optimal levels of task difficulty because hand/arm and near/far differences may not be seen if the task is either too easy or too difficult. Differences in target size significantly affected task difficulty which varied markedly from Level 1 (mean response time = 1086 ms) to Level 5 (mean response time = 2649 ms). Hence we conducted separate 3-way ANOVAs at each level of difficulty. As seen in Table 2, the predicted interactions were not significant at any level of difficulty. However, while there is no pattern across levels of difficulty for the muscle data, the space data show larger effects at the easier Levels 1 and 2 than at the more difficult Levels 3-5.
Study 1: F-values, probabilities and effect sizes (ηp 2) for the predicted Muscle*Sex and Space*Sex interactions at each level of difficulty from Level 1 (easy) to Level 5 (difficult).

Sex differences in control of hand and arm
The absence of a pattern in the muscle data (Table 2) suggests the present failure to replicate the Muscle*Sex interaction is not a question of task difficulty but rather the result of differences between the tasks used. The present target cancellation task required a relatively simple ballistic movement from the starting position to the target. In contrast, the tracking tasks used by Sanders and Walsh (2007), which showed women were better with the hand than the arm and men better with the arm than the hand, were more complex, requiring a constant speed and continuous changes of direction. Perhaps a ballistic movement from a start point to an end point is too simple to reveal differences in hand and arm use.
Sex differences in processing near and far space
In contrast to the muscle data (Table 2), in the space data the Partial Eta Squared values, although small, indicate that the Space*Sex interaction accounted for more of the variance at the easier levels (1 and 2) than at the more difficult levels (3-5). There is a related tendency for the Space*Sex interaction to approach significance at the easier levels of difficulty. Indeed, at Level 1, the easiest task, women were faster in near than in far space as predicted (t 23 = 1.762, p = 0.046, one-tailed) while men were nominally faster in far space than near space but not significantly so (t 23 = 0.595, p = 0.279, one-tailed). These observations suggest that the use of easier target cancellation tasks might reveal the predicted Space*Sex interaction.
STUDY 2
Here we continued to investigate sex differences in the visual processing of information from far and near space using a target cancellation task but, in the light of the findings from Study 1, we introduced two changes. First we adjusted the range of target sizes to include some levels of difficulty that were easier than those used in Study 1. Second, given the absence of a hand/arm effect from Study 1, we dropped the Muscle (hand/arm) factor by requiring participants to use a mouse to move the cursor. The same participants completed both Study 2 and 3 (see below), one after the other in a counterbalanced order within the same test session.
Materials and Methods
Participants
Forty-eight participants, 24 women (mean age 27.28, range 20 to 46 years) and 24 men (mean age 27.38, range 19 to 44 years), were recruited as an opportunity sample from among our undergraduate and postgraduate students, and graduates from other universities. All were right handed and had normal or corrected to normal vision. None was color-blind and none had sustained a right hand injury during the previous six months. All were na ï ve to the specific aims and predicted outcomes of the study which was approved by the Departmental Ethics Committee. Each participant gave informed written consent and was aware that they could withdraw from the study at any time. None withdrew.
Task and procedure
Here we describe those aspects of the task and procedure that differed from Study 1. As before we used a mixed design but without the Muscle (hand/arm) factor. In Study 2, Sex was a between-participants factor with two independent groups, women and men; Space and Difficulty were within-participants factors with repeated measures on near/far and level of difficulty from the easy, largest diameter, target (Level 1) to the difficult, smallest diameter, target (Level 6). In the near condition the diameter of the targets at each level of difficulty was: Level 1, 109 mm; Level 2, 94 mm; Level 3, 72 mm; Level 4, 55 mm; Level 5, 36 mm; Level 6, 26 mm. Hence, Levels 1 and 2 were easier than Level 1 in Study 1 while Levels 3 to 6 spanned a similar range to Levels 1 to 3 in Study 1. Image size in the far condition was adjusted so that both near and far stimuli subtended the same visual angle at the retina. The joysticks used in Study 1 were replaced by a standard ball-roll mouse that was used by participants to move the cursor and to click on and cancel targets. As before, within each condition participants completed one block of 5 trials at each level of difficulty which started with Level 1 and continued sequentially through the levels, this time to Level 6. Other aspects of the task and procedure were as shown in Figure 1 and described for Study 1.
Results and Discussion
Initial analysis
Table 3 summarizes the target cancellation times from Study 2 that were submitted to a 3-way mixed ANOVA with Sex (women/men) as a between-participants factor and with Space (near/far) and Difficulty (Levels 1 to 6) as within-participants factors. There were significant main effects of Space, Sex and Difficulty. Overall, participants were faster in near than in far space (F 1,46=59.90, p<0.001), faster with larger than with smaller targets (F 5, 230=317.00, p<0.001), and men were faster than women (F 1,46=4.18, p=0.047). There was a significant two-way interaction between Space and Difficulty (F 5,230=5.03, p=0.001) that arose because the increase in response times from Level 1 to 6 was greater in far than in near space. It is likely that men were faster than women overall because male participants frequently reveal greater competitiveness when completing such tasks.
Study 2: Mean (SEM) response times recorded by men and women when using a computer mouse to cancel targets presented in near or far space. Task difficulty was manipulated by varying the target size from large (Level 1) to small (Level 6). The significant main effects are high-lighted in bold color (

Further analysis
The predicted Space*Sex interaction (Table 3) narrowly failed to reach significance (F1,46=4.00, p=0.051). However, we had argued a priori that the Space*Sex interaction might appear at some but not all of the Levels 1 to 6 because sex differences are sensitive to task difficulty. Consequently, even though the three-way interaction between Space, Difficulty and Sex was a not significant (F 1,46=0.10, p=0.991), we conducted two-way ANOVAs at each level of difficulty. From Study 1 (Table 2) we would expect the Space*Sex interaction to appear at target sizes greater than 70mm, essentially Levels 1 or 2 in Study 2. In fact, we found that Level 2, which had a target size of 94 mm, showed a significant Space*Sex interaction (F 1,46=4.560, p=0.038) arising because, compared with men, women were relatively faster in near than in far space (Figure 3).
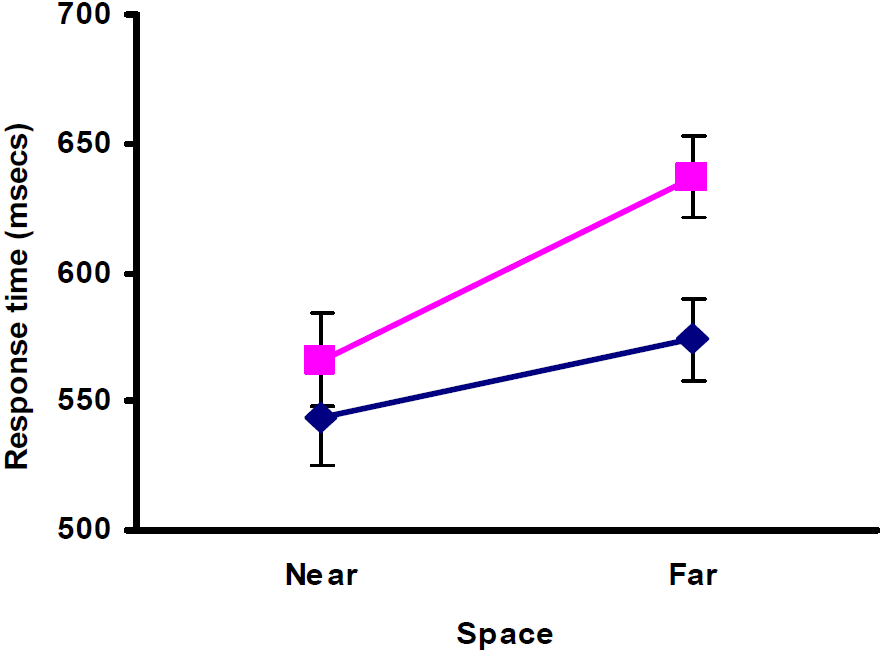
Study 2: Means (+/-SEM) of the time taken to cancel targets illustrating the significant Space*Sex interaction obtained for the relatively easy Level 2 version of the task. (Women – pink /light grey squares; Men – blue /dark grey diamonds.)
The significant main effect of space can be seen in Figure 3 where both women and men were faster in near space. However, the predicted Space*Sex interaction still appeared because, compared with men, women were relatively faster in near than in far space. Near space performance was significantly faster than far space performance for both women (t 23=4.57, p<0.001, one-tailed) and men (t 23=2.74, p=0.006, one-tailed) because far space proved more difficult than near space. However, although there was no significant difference between the scores of women and men in near space (t 46=0.87, p=0.388, two-tailed), women were significantly slower than men in far space (t 46=2.80, p=0.007, two-tailed).
Although the Space*Sex interaction was significant at Level 2 only, effect sizes (Cohen's d) and t-tests show a similar pattern of within-sex and between-sex findings across the levels of difficulty (Table 4). From the within-sex comparisons we see that both women and men were consistently faster in near space than in far space, except for the single case of men at Level 1. From the between-sex comparisons we see that men were never significantly faster than women in near space but, in far space, men were significantly faster than women at all levels except the extremes (Levels 1 and 6).
Study 2: The performance of women and men in near and far space at each level of difficulty as shown by effect size (d) and by within-sex and between-sex t-tests. Significant outcomes are in bold. The between-sex comparisons are presented as men faster than women because men were faster than women (although not always significantly) at each level of difficulty in near and far space (Table 3).

From the these analyses, we see that Study 2 provided some evidence for the predicted Space*Sex interaction but the differential performance of women and men in far and near space was obscured by the greater difficulty that both sexes experienced with far compared with near space and the overall faster response times recorded by the men. To investigate the topic further, in Study 3 we used a different version of the target cancellation task.
STUDY 3
The target cancellation task used for Study 2 manipulated level of difficulty by varying the size of the targets. In Study 3, task difficulty was manipulated by presenting the same sized target with a varying number of distracter stimuli. The same participants completed Studies 2 and 3, one after the other within the same test session.
Materials and Methods
Participants
The same 48 participants were used for Study 3 as were used for Study 2 (see above). They completed the two studies in a counterbalanced order.
Task and procedure
The design, tasks and procedure were the same as for Study 2 with one exception; here task difficulty was manipulated by varying the number of distracters that were presented with the target. To accommodate sufficient numbers of distracters on the screen, both they and the targets were 27 mm in diameter (in the near condition) but they differed in color, light orange for the targets and yellow for the distracters. The six levels of difficulty were defined by the number of distracters presented with the target as follows: zero at Level 1, one at Level 2, three at Level 3, five at Level 4, seven at Level 5, and nine at Level 6. The targets and distracters could appear in any one of 104 different screen positions. The procedure was the same as that used for Study 2 except that (i) difficulty was manipulated by using distracters rather than target size, (ii) a small blue circle (22 mm diameter in the near condition) was used for the starting position, and (iii) Level 2 (the easiest distracter condition) was used for the practice trials. A typical sequence of screens that constituted a trial is illustrated in Figure 3a.

An example of the sequence of screen presentations (not drawn to scale) that were used for the target cancellation tasks in Study 3 in which difficulty was manipulated by varying the number of distracters. The cross represents the cursor, the central blue circle the starting position, the two larger yellow circles the distracters and the light orange circle the target. Further explanation is presented in the text.
Results and Discussion
Initial analysis
Table 5 summarizes the target cancellation times from Study 3 that were submitted to a 3-way mixed ANOVA with Sex (women/men) as a between-participants factor and with Space (near/far) and Difficulty (Levels 1 to 6) as within-participants factors.
Study 3: Mean (SEM) response times recorded by men and women when using a computer mouse to cancel targets presented in near or far space. Task difficulty was manipulated by varying the number of distracters from zero (Level 1) to nine (Level 6). Significant main effects are high-lighted in bold color (

There were significant main effects of Space and Difficulty but not Sex (F 1,46=2.270, p<0.139). Overall, participants were faster in near than in far space (F 1,46=86.17, p<0.001) and faster with fewer than with more distracters (F 5, 230=18.19, p<0.001). The predicted Space*Sex interaction was significant (F 1,46=12.17, p=0.001). It arose because, compared with men, women were relatively faster in near than in far space. In addition, there was a significant three-way Space*Task*Sex interaction (F 5,230=2.49, p=0.032) arising because greater differences between near and far space performance were recorded by women at Levels 1, 2, 3 and 5 but by men at Levels 4 and 6.
Further analysis: the Space*Sex interaction
In Study 2 (Table 3), the predicted Space*Sex interaction narrowly failed to reach significance (F 1,46=4.00, p=0.051), however, in Study 3 (Table 5) both the two-way Space*Sex and the three-way Space*Task*Sex interactions were significant. The three-way interaction confirms our a priori argument that the two-way Space*Sex interaction may appear at some but not all of the Levels 1 to 6 because sex differences are sensitive to task difficulty. The two-way interactions at each level of difficulty are plotted in Figure 4. Two-way ANOVAs revealed that the Space*Sex interaction approached significance at Level 1 (F 1,46=2.90, p=0.096), was significant at Level 2 (F 1,46=11.82, p=0.001), Level 3 (F 1,46=9.68, p=0.003), and Level 5 (F 1,46=9.06, p=0.004), but not at Level 4 (F 1,46=0.00, p=0.969), or Level 6 (F 1,46=0.70, p=0.406). On the assumption that a task may be too easy or too difficult to reveal the predicted interaction then we might expect it to be non-significant at one or both extremes but, if significant at all, it should be consistently significant between the extremes. Consequently, the present pattern, with an isolated non-significant Space*Sex interaction at Level 4, is puzzling. Inspection of the data for Level 4 revealed a possible explanation for this anomalous finding.

Study 3: Mean (+/-SEM) time taken by men and women to cancel targets presented in near or far space. Task difficulty was manipulated by varying the number of distracters. Significant Space*Sex interactions were found at Levels 2, 3 and 5 but not Levels 1, 4 and 6. (Women – pink /lighter grey lines and squares; Men – blue /darker grey lines and diamonds.)
As seen in Figure 4 and Table 5 the male far space scores increased steadily from Level 1 to Level 6 except for the high score at Level 4 which is responsible for the absence of the Space*Sex interaction at that level. Response time is dependent on target location with higher scores recorded for peripheral than for more central targets. The target positions were determined randomly by the computer program. We found that, by chance, there were more peripheral target locations in the male Level 4 condition than in any other condition. Crucially, at Level 4, male participants who experienced three or more peripheral targets were 262 ms slower than those who encountered fewer than three. Thus our use of a random rather than a quasi-random selection procedure for target location biased the male Level 4 far condition towards higher response times. It would appear that this bias led to the anomalous absence of a significant Space*Sex interaction at Level 4.
Further analysis: the performance of men and women in near and far space
As noted above, the two-way Space*Sex interactions at each level of difficulty (Fig. 4) were significant at Levels 2 to 5 with the anomalous exception of Level 4. We suggested these interactions arose because, compared with men, women were relatively faster in near than in far space. This contention is supported by the associated effect sizes (Cohen's d) and t-tests conducted on data from each of the levels of difficulty (Table 6). From the within-sex comparisons we see that women, but not men, were consistently faster in near space than in far space. From the between-sex comparisons we see that men were never significantly faster than women in near space but, in far space, men were significantly faster than women at all levels except the anomalous Level 4 and the extremes (Levels 1 and 6). Thus Study 3 provides substantial evidence for the Space*Sex interaction although the predicted differential performance of women and men in far and near space was partially obscured by the greater difficulty that both sexes experienced with far compared with near space.
Study 3: The performance of women and men in near and far space at each level of difficulty as shown by effect size (d) and by within-sex and between-sex t-tests. Significant outcomes are in bold. Following the convention adopted for Table 4, the between-sex comparisons are shown as men faster than women although they were not faster at every level of difficulty in near and far space (Table 5).

General Discussion
In the present article we have reported three studies that further investigated the predictions that we have made from the Hunter-Gatherer Hypothesis and found no support for the Muscle*Sex interaction and limited support for the Space*Sex interaction. However, we shall argue that the present null findings, and some of the previous ones, may be explained by the nature of the tasks used.
Motor control: Failure to find the predicted Muscle*Sex interaction
Sanders and Walsh (2007) predicted that women should perform better with the muscles of the hand and wrist, while men should perform better with the muscles of the arm and shoulder. These reciprocal sex differences should appear as a Muscle*Sex interaction. Sanders and Walsh demonstrated these predicted outcomes in two separate studies using computer-based tracking tasks and their findings were confirmed by Sanders and Perez (2007) with a token relocation task. Consequently, we were surprised when we failed to find the same Muscle*Sex interaction in the present Study 1 which used a target cancellation task based closely on the original tracking tasks to the extent that the same short (hand condition) and long (arm condition) joysticks were used. We suggest that the present failure to find the Muscle*Sex interaction arose from the nature of the movement required to complete the tasks. The tracking and token relocation tasks demand relatively complex movements whereas all that was needed for the present target cancellation task was a ballistic movement with relatively little re-adjustment. Perhaps the motor component was too simple to reveal the interaction. In summary, there are currently three studies using complex motor tasks that have found the Muscle*Sex interaction and one using a simple ballistic movement that has not.
It has been argued (Peters and Campagnaro, 1996) that that the female advantage at manual dexterity tasks arises simply because women have smaller fingers than men. While this is a plausible explanation for a female advantage on pegboard tasks it seems less likely for the joystick control required in the present Study 1 and in the original studies reported by Sanders and Walsh (2007). Peters and Campagnaro supported their claim by showing that the female advantage became a male advantage when the size of the pegs was increased. Sanders and Perez (2007), pages 795-796) concluded that the appearance of a female or male advantage depended not on finger size (as reflected in the size of the pegs) but on the extent to which hand or arm movements were used to complete the task.
Visual processing: Failures to find the Space*Sex interaction
As noted above, on the basis of the hunter-gatherer hypothesis, Sanders, Sinclair and Walsh (2007) predicted that studies using stimuli presented in near and far space should reveal a significant Space*Sex interaction. Evolutionary selection for gathering-related skills should lead women to perform better with visual information from near space, while selection for hunting-related skills should lead men to perform better with visual information from far space.
Table 7 summarizes seven studies that have investigated the processing of visual information from near and far space by women and men. Two studies have not found a significant Space*Sex interaction and its associated within-sex differences. The other five studies found the interaction but three of those did not find clear cut evidence for the associated reciprocal within-sex differences. We have identified three possible reasons for the absence of these predicted outcomes: tool use, task difficulty, and a main effect of space.
Summary of studies of visual information processing of near and far space by women and men. Significant statistical outcomes are shown for the predicted Space*Sex interaction and for the reciprocal sex differences, i.e. for women performing better in near than far space (N>F) and for men performing better in far than near space (F>N). Comments, which are expanded in the text, replace the statistical data where the outcome was not clear cut.

Tool use
Sanders and Perez (2007) Study 2 did not find the Space*Sex interaction nor the associated within-sex outcomes (Table 7 [2] Study 2). The authors attributed this failure to the nature of the task in which participants used tools, a short and a long stylus, to interact with near and far space. A number of studies have indicated that if a tool is used to reach into far space then that portion of far space touched by the tool is re-mapped as near space. For example, Berti and Frassinetti (2000) describe a patient with visual neglect for near, but not far, space who showed neglect for both near and far space when using a tool (a stick) to bisect lines. Similar findings have emerged from studies of cross-modal extinction in patients with unilateral cortical lesions (Farne and Ladavas, 2000; Maravita, Clarke, Husain and Driver, 2002; Maravita, Husain, Clarke and Driver, 2001) and from a study of cross-modal interference in humans with intact brains (Maravita, Spence, Kennett and Driver, 2002). Electrophysiological evidence from macaques trained to retrieve objects from beyond hand reaching distance using a rake has actually shown the re-mapping taking place (Iriki, Tanaka and Iwamura, 1996). The receptive fields of neurons receiving convergent visual and somatosensory inputs, which normally covered the hand and near space, expanded to include the whole of the rake and the portion of far space that it reached. These findings imply that a tool is processed by the brain as part of the hand and that the portion of far space that can be reached with the tool is temporarily re-mapped as being within reaching distance and processed as near space. If so, this would explain the failure of Sanders and Perez (2007) to find the near/far sex difference because their participants' tool use would have caused their brains to process the far space task as if it were in near space.
Task difficulty
In Study 1 of the present report the interaction did not appear overall (Table 7 [3] Study 1) nor at any level of difficulty (Table 2). However, task difficulty is a crucial factor for the demonstration of sex-related effects (Sanders, Sjodin, and de Chastelaine, 2002; Sanders and Walsh, 2007) and we have argued above that a trend across the levels of difficulty in the data indicates that the range of difficulty used in this study was too high and that a lower level would be required to reveal the interaction. This view was effectively confirmed by Study 2 of the present report which introduced easier levels of difficulty and where the overall Space*Sex interaction was significant at p=0.051 (Table 7 [3] Study 2). Although Studies 1 and 2 may imply a precise effect, in Study 3 the range of difficulty was quite broad, from Level 2 to Level 5 (one to seven distracters) if we ignore the anomalous finding at Level 4. Levels of difficulty are critical when ceiling or floor effects come into play. A sex-dimorphic task will not reveal a difference if it is too easy (ceiling effect) or too difficult (floor effect). For tasks where the dependent variable is percent correct we would define the optimum level of task difficulty as one that fell mid-way between 100% and a chance level of performance. For tasks where the dependent variable is response time then the only guide would be to avoid ceiling and floor effects by avoiding levels of difficulty that lead to very fast or very slow response times.
A main effect of space
Interpretation of the reciprocal within-sex differences is difficult when there is a significant main effect of Space as was the case for three of the studies summarized in Table 7. In Sanders, Sinclair and Walsh (2007) Study 1 participants overall were better in far than in near space (Table 7 [1] Study 1), an outcome that could have accentuated the “far better than near” prediction for men which was supported (t 63=3.76, p=0.001) but reduced the “near better than far” prediction for women that was not supported (t 63=0.273, p=0.237). Similarly, in the present Study 2 and 3 participants overall were better in near than in far space (Table 7 [3] Study 2 and 3), outcomes that accentuated the “near better than far” prediction for women and reversed the “far better than near” prediction for men. Thus both sexes were better in near than far space (women: Study 2 t 23=6.09, p<0.001, Study 3 t 23=9.30, p<0.001; men: Study 2 t 23=4.78, p<0.001, Study 3 t 23=3.99, p=0.001). Of course, for all three studies the Space*Sex interaction was significant but, as we have argued in detail for the present Studies 2 and 3 (Tables 3, 4 and 6, Fig. 4, and the associated text), the appearance of the predicted within-sex differences was obscured by the fact that all participants were faster in near than far space. It is important to note that, in these two studies, participants used a standard computer mouse to perform the target cancellation task. We suggest that the reversal of the predicted male performance arose because of the extensive practice that our participants will have had using a mouse to interact with visual stimuli displayed on a computer monitor in near space. Notwithstanding the main effect of Space, significant Space*Sex interactions arise because there are relative differences in the performance of women and men in near and far space that are consistent with the predictions.
Thus, of the seven studies discussed, two (Table 7 [2] Study 2 and [3] Study 1) did not find the predicted Space*Sex interaction and the associated reciprocal within-sex differences. However, plausible reasons (tool use and level of difficulty) are offered as explanations for these failures. The remaining five studies report significant Space*Sex interactions (albeit, one at p=0.051) but for three the associated reciprocal within-sex differences were obscured by a main effect of space. We conclude that the weight of the evidence supports the view that women perform better when visual information is presented in near rather than far space and that men show the reverse pattern. Our current analysis of the failures to find the Space*Sex interaction and associated reciprocal sex differences in the present studies and those reported elsewhere (see Table 7 for references) indicate that future studies should identify and use tasks of an appropriate level of difficulty that do not have an inherent bias towards near or far space, i.e. there should be no main effect of Space when the scores of women and men are combined. While the absence of a main effect of Sex would be desirable it is not essential because the hypothesis is that, when compared with themselves, women will be better in near and men in far space; hence it is the within-sex, not the between-sex, differences that are critical.
The two visual systems: Possible near and far components that are sex dimorphic
We conclude that present evidence supports the view that there is a reciprocal sex difference, with women favoring information from near and men from far space. Studies using patients with selective radial neglect and brain scans with non-clinical participants indicate a separation of the brain mechanisms for the visual processing of far (extrapersonal) and near (peripersonal) space. Near space is processed by the dorsal cortical stream and far space by the ventral cortical stream (for a brief review see Sanders, Sinclair and Walsh, 2007). However, the traditional view of these two visual streams was established by Ungerleider and Mishkin (1982). They identified the ventral stream as an object identification system and the dorsal stream as an object localization system, the “what” and “where” processes of visual perception. Goodale and Milner (1992) re-formulated the functions of the ventral and dorsal streams as visuoperceptual and visuomotor, the “what” and “how” processes of visual perception. These traditional-two-visual-system models do not incorporate the far/near processing functions, probably because most studies of visual systems are conducted with stimuli in near space. Consequently, as suggested by Sanders, Sinclair and Walsh (2007), it is advisable that the spatial component be added to the functions of the two visual systems. In addition to their “what”/“where” and “what”/“how” functions we should recognize that the ventral stream has a “there” component interacting with far space and favored by men, while the dorsal stream has a “here” component interacting with near space and favored by women. Future studies of visual systems should be designed, conducted and interpreted with these space and sex differences in mind.
Footnotes
Acknowledgements
We are grateful to our colleague, John Robinson, who wrote the customized computer programs for each of the three studies.