
Abstract
Background:
Fluid retention is linked to the physiology and pathophysiology of humans at high altitude (HA). The angiotensin-converting enzyme (ACE) gene plays a role in the regulation of plasma volume and vascular tone.
Materials and methods:
In this meta-analysis, eligible studies published before 1 September 2015 that focused on the association between the ACE insertion/deletion (I/D) polymorphism and HA adaption were identified by searching the PubMed, Web of Science, Embase and Medline online databases. We used a fixed-effects model and assessed the study qualities multiple times.
Results:
The seven selected studies included a total of 582 HA-native individuals and 497 low-altitude controls, and these subjects were analyzed for the ACE I/D gene polymorphism. A significant association was found between the ACE DD genotype and HA maladaptation. The results for genotype DD versus ID + II were as follows: Odds ratio (OR) = 0.46; 95% CI 0.31–0.70; p = 0.0002. The results for genotype ID versus DD were as follows: OR = 1.97; 95% CI 1.27–3.06; p = 0.002.
Conclusions:
Our findings suggested that the DD genotype of ACE is a risk factor for HA maladaptation and that the presence of fewer ACE DD allele carriers in a population indicates a greater ability of that population to adapt to HA.
Keywords
Introduction
Approximately 140 million people live at altitudes > 2500 m above sea level. 1 These high altitude (HA) areas have been inhabited for several thousand years 2 ; and unlike individuals living at lower altitudes, the natives or inhabitants are better adapted to HA. In the last decade, a significant number of studies have investigated HA adaptation and disorders3,4 and whether HA populations have undergone natural selection for features beneficial to the absorption and utilization of oxygen. Some reviews have explored HA adaptation in relation to genetics.2,5,6 Previous studies report that HA adaptation may be associated with certain genes, including several found in mitochondrial deoxyribonucleic acid (mtDNA)5,7,8; and the genes encoding endothelial Per-Arnt-Sim (EPAS),1,9,10 erythropoietin (EPO),11,12 myoglobin, 13 hypoxia-inducible factor 1 (HIF1), 14 angiotensin-I converting enzyme (ACE),15–17 aldosterone synthase (CYP11B2), 18 nitric oxide synthase 3 (NOS3), endothelin 1 (EDN1)19,20 and beta-2 adrenergic receptor (ADRB2). 21
ACE is a particular key component of the renin-angiotensin-aldosterone system (RAAS); and it plays important roles in the physiology and pathophysiology of humans living at HA22,23 and the pathogenesis of HA sickness 24 : These roles of ACE are most likely achieved through its effects on the regulation of blood pressure,25,26 through the generation of the potent vasoconstrictor angiotensin II (AngII) and through the blockade of the production of the vasodilator bradykinin. 27 In addition, studies report that ACE activity may contribute to adaptive physiology by affecting the adaptive respiratory response in hypoxic conditions,28,29 by influencing the normalization of pulmonary artery pressure, 30 and the maintenance of sodium and fluid balances. 23
An ACE polymorphism has been described that involves the insertion (I)/deletion (D) of a 287-base pair (bp) Alu repeat element in intron 16, 31 which is a marker of the ACE functional polymorphism. 32 Montgomery et al. 33 hypothesized that the D allele is associated with altered levels of serum ACE. Droma et al. 34 reported that the over-representation of the I allele of the ACE gene in Sherpas might have an advantageous physiological role in adapting to a high-altitude environment. In addition, several investigations have focused on the relationship of the ACE gene I/D polymorphism with the human adaptation to HA.13,31 Qadar et al. 35 report that the genotype II may play a beneficial physiological role in HA adaptation; and that the I allele may contribute to routine physical activities, by aiding in adaptation to the hypoxic environment at HA. We performed this meta-analysis to investigate whether an association exists between the ACE gene I/D polymorphism and HA adaptation in native HA inhabitants.
Materials and methods
Literature search
We searched the literature published before 1 September 2015 in the following databases: PubMed, Embase, Web of Science and Medline. We performed an advanced search using several search options. The following keywords were used: angiotensin-converting enzyme or ACE, high altitude or HA, adaptation and polymorphism. We examined the bibliographies of the included papers. The articles were limited to those written in English. This study was approved by the Ethical Committee of the Third Military Medical University in China.
Inclusion and exclusion criteria
Studies with the following criteria were included in the meta-analysis:
The inclusion of HA cases involving native HA inhabitants;
A comparison of high- and low-altitude subjects;
An analysis of the ACE gene I/D polymorphism;
The provision of sufficient data to analyze the results; and
The inclusion of control data that was in agreement with Hardy-Weinberg equilibrium.
Studies with the following characteristics were excluded from the meta-analysis:
Presence of HA diseases;
Lack of information regarding genotype number or frequency;
Lack of comparisons between high- and low-altitude subjects or the inclusion of comparisons of identical subjects at low and high altitudes; and
Inclusion of control data that was not in accordance with Hardy-Weinberg equilibrium.
Data extraction
The following characteristics were recorded for each study: first author, year of publication, study ethnicities or areas, study population and ACE I/D genotypes of the cases and controls. All cases were permanent residents and were members of families that had resided in the HA area since ancient times and had acclimated to HA.
Statistical analysis
In this meta-analysis, we evaluated the associations between the I/D alleles of the ACE gene and HA adaptation, using the following four comparisons:
An allelic comparison (D versus I);
Comparison of the DD homozygous genotype to the other genotypes (DD versus ID + II and DD versus II);
Comparison of the ID heterozygous genotype to the other genotypes (ID versus DD and ID versus II); and
Comparison of the II homozygous genotype to the other genotypes (II versus DD + ID).
We used the odds ratio (OR) as a measure of the strength of the associations between polymorphisms of the ACE I/D gene and HA adaptation risk across studies. We compared the frequencies of the alleles and genotypes between the cases and controls, using the unadjusted OR and the relevant 95% CI. We also used the chi-square-based Q test and I 2 test, to evaluate heterogeneity among the studies. If there was significant heterogeneity (p < 0.05), we used a random-effects model to calculate the pooled OR of each study. If there was no heterogeneity (p > 0.05), the pooled OR was calculated using the fixed-effects model.24,36–38 The Bonferroni correction was used to adjust for multiple hypothesis testing; and p < 0.008 was considered to be statistically significant. We also used Begg’s test and Egger’s test to evaluate potential publication bias, which was considered to be significant at p < 0.05. All analyses were performed with RevMan 5.0 software (The Cochrane Collaboration) according to Qi et al. 39
Results
Study characteristics
We identified 96 reports using the keywords “angiotensin-converting enzyme” and “altitude”. We excluded 90 reports by applying our inclusion and exclusion criteria. Among the 90 excluded reports, 38 did not include information about the ACE I/D gene polymorphism, seven included HA diseases, and 45 of these reports did not compare high- and low-altitude subjects. Finally, for this meta-analysis, we selected seven studies (of note, the report by Rupert et al. 40 provided two sets of data; therefore, we considered these to be two independent studies) with 582 HA cases and 497 low-altitude controls. In Table 1, we present the characteristics of the studies that evaluated the associations between the ACE I/D polymorphism and HA adaptation. Three studies examined Quechua populations; and the remaining studies focused on Sherpa, Tibetan, Peruvian and Indian populations. The sample sizes of these studies ranged from 92 to 257 subjects.
Characteristics of studies evaluating the influences of ACE I/D polymorphism on HA adaptation.
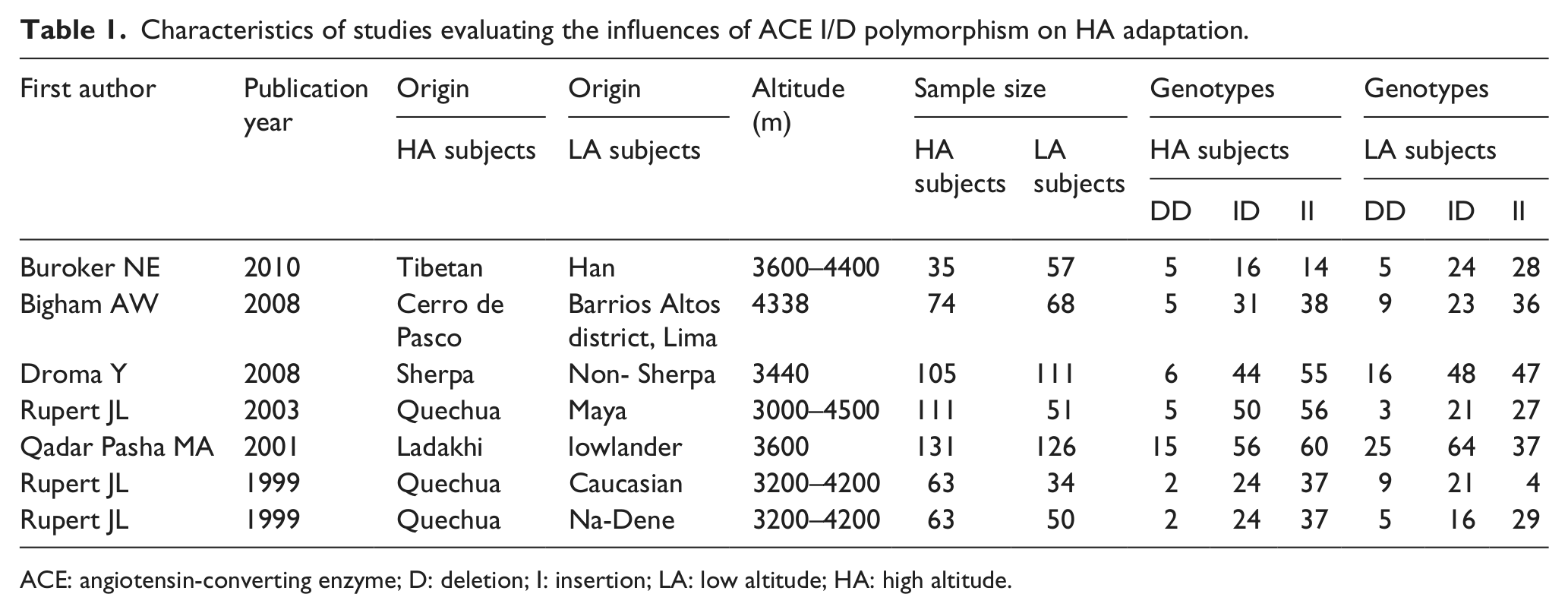
ACE: angiotensin-converting enzyme; D: deletion; I: insertion; LA: low altitude; HA: high altitude.
Meta-analysis results
The summary OR, which was obtained using a random effects model, did not reveal a significant difference in HA adaption between carriers of the ACE D and I alleles (Table 2) (OR = 0.70; 95% CI 0.49–1.02; p = 0.06) (Figure 1); however, the summary OR of the fixed-effects model showed a significant difference in HA adaptation between the DD and ID + II genotypes (OR = 0.46; 95% CI 0.31–0.70; p = 0.0002) (Figure 2). Furthermore, the summary OR obtained using a random-effects model with Bonferroni correction did not reveal a significant difference in HA adaptation between the DD and II genotypes (OR = 0.39; 95% CI 0.18–0.84; p = 0.02) (Figure 3). In addition, no significant difference in HA adaptation was observed between the DD + ID and II genotypes (OR = 1.41; 95% CI 0.87–2.29; p = 0.16) (Figure 4); however, a significant difference in HA adaptation was found between the ID and DD genotypes (OR = 1.97; 95% CI 1.27–3.06; p = 0.002) (Figure 5). Finally, no significant difference in HA adaptation was observed between the ID and II genotypes (OR = 0.80; 95% CI 0.51–1.26; p = 0.33) (Figure 6).
Meta-analysis of the association of ACE I/D polymorphism with HA adaptation.

ACE: angiotensin-converting enzyme; D: deletion; I: insertion; No: number of polymorphisms.
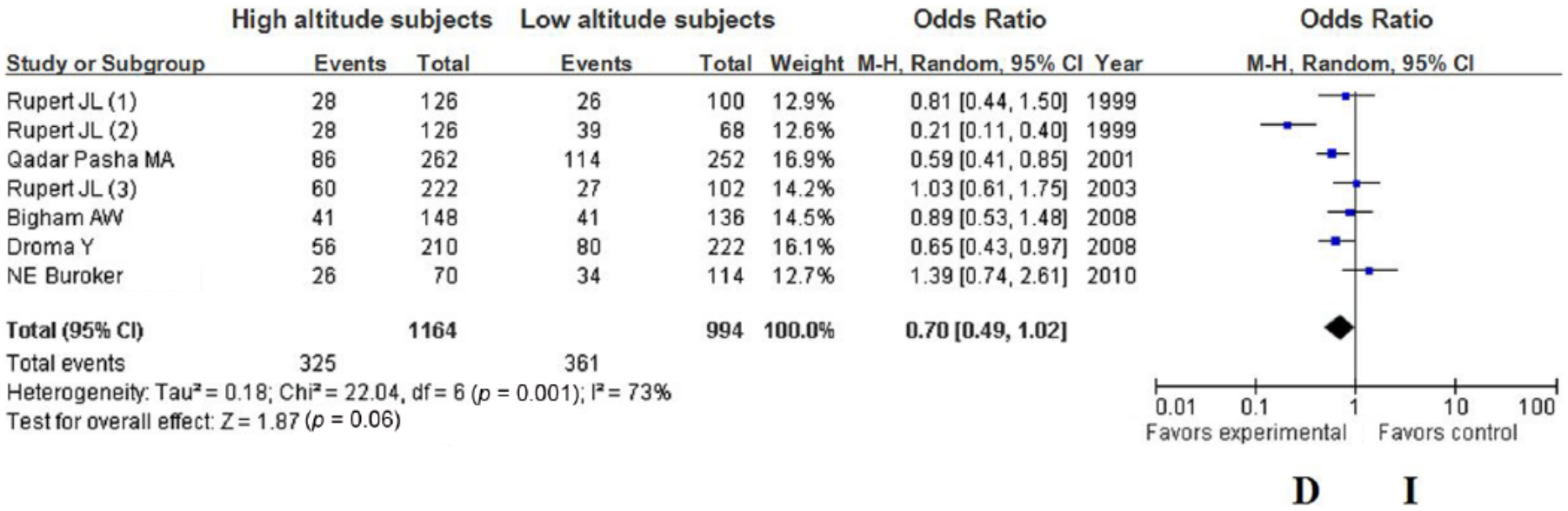
Forest plot of the ACE I/D polymorphism: D versus I.

Forest plot of the ACE I/D polymorphism: DD versus ID and II.
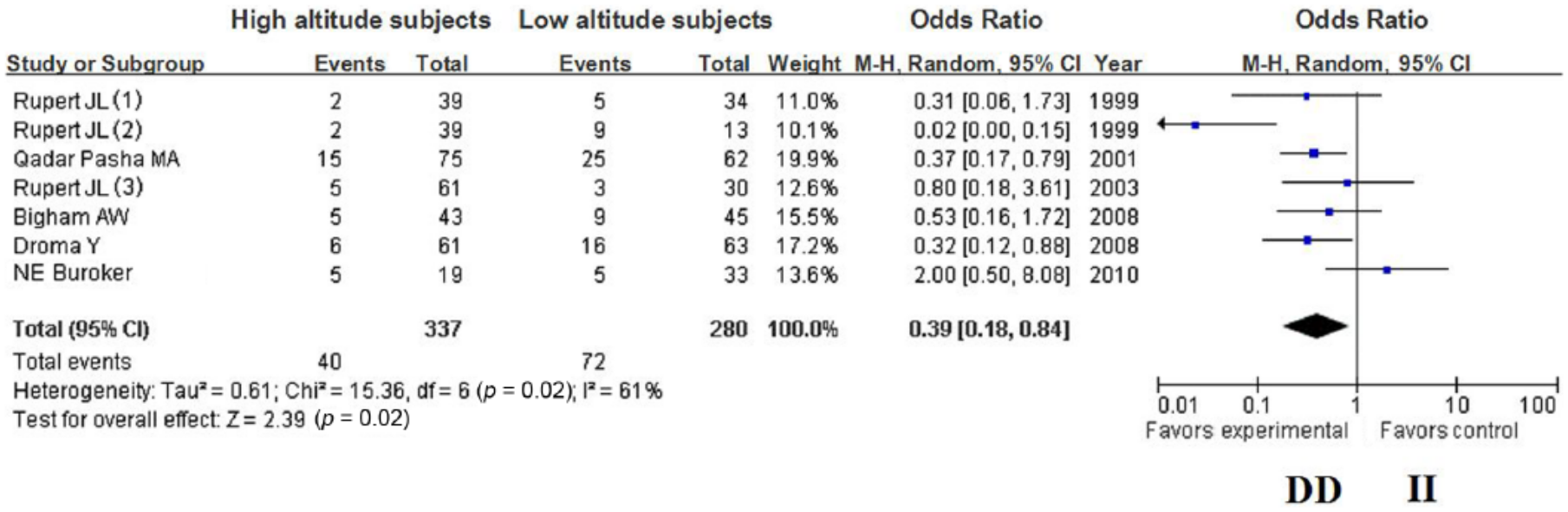
Forest plot of the ACE I/D polymorphism: DD versus II.

Forest plot of the ACE I/D polymorphism: DD + ID versus II.

Forest plot of the ACE I/D polymorphism: ID versus DD.
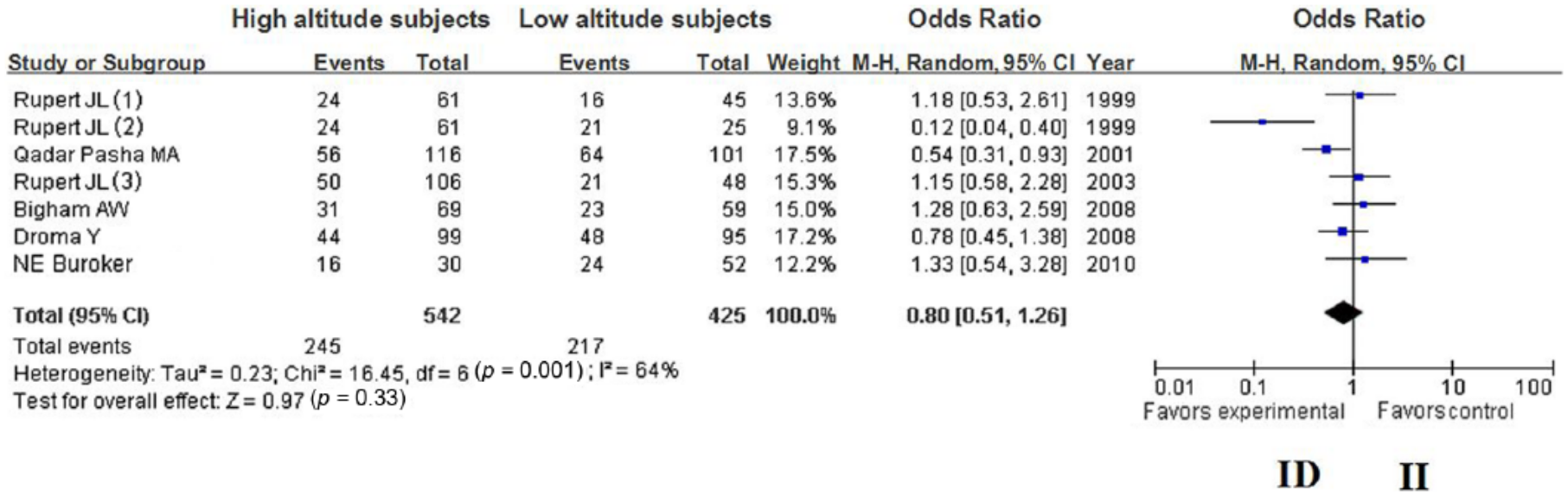
Forest plot of the ACE I/D polymorphism: ID versus II.
Discussion
To our knowledge, this is the first meta-analysis to evaluate the association of the ACE gene I/D polymorphism with HA adaptation. Seven studies with a total of 582 native HA inhabitants and 497 native low altitude inhabitants were included in this meta-analysis. No significant difference in HA adaptation was found between the ACE D allele and the I allele (p < 0.05). For the comparison of the DD versus the ID + II genotypes, the OR = 0.46; and for the comparison of the ID versus the DD genotypes, the OR = 1.97. These differences remained significant after Bonferroni correction. The frequency of the DD genotype in the HA subjects (0.31) was significantly lower than that in the low-altitude subjects (0.52); therefore, the possession of a lower frequency of the representative DD genotype of the ACE I/D polymorphism in a population may be beneficial to HA inhabitants, by increasing their tolerance to hypoxia at HA. We inferred that the DD genotype was a risk factor for maladaptation to HA. Having fewer ACE DD allele carriers in a population indicates a greater ability of individuals in that population to adapt to HA.
In support of our meta-analysis results, the following factors may explain why the DD genotype is a risk factor for HA maladaptation. First, ACE polymorphisms affect circulating ACE activity.41–44 Mean plasma ACE levels in individuals with the DD genotype are higher than those in individuals with the II genotype, and individuals with the ID genotype have intermediate mean plasma ACE levels 31 ; therefore, we could conclude that higher ACE DD genotype might contribute to ACE activity31,42 and therefore increased the production of angiotensin II, and that the higher ACE DD genotype might increase the risk of HA maladaptation. Some studies report that subjects with the DD genotype have higher plasma angiotensin II concentrations after angiotensin I infusion,45,46 and the Type I receptor of angiotensin II was shown to initiate Ca2+ signaling that mediates the contraction of vascular smooth muscle cells, 17 which can influence blood pressure. Angiotensin II also controls water and salt retention, through the release of aldosterone. 47 High blood pressure and water and salt retention are related to the occurrence of HA diseases, so these findings suggest that a higher DD genotype frequency in a population is not beneficial to HA adaption. Lachurie et al. 48 report that the ACE I/D polymorphism has no influence on the production of Ang II nor plasma aldosterone, and does not cause a rise in blood pressure; these findings may have been due to the lack of elimination of potential differences in Ang I conversion rates at the coronary and renal vessels. Furthermore, the ACE DD genotype was shown to be associated with increased bradykinin degradation, which is associated with significant blunting of the vasodilatory action of nitric oxide. 49 Nitric oxide decreases the occurrence of mountain sickness, so the ACE DD genotype decreases a person’s ability to adapt to HA. In addition, reduction in ACE activity level was reported to cause an increase in Ang 1–7, which is a vasodilator. 50 The activities of Ang 1–7 may further contribute to vasodilation, so Ang 1–7 may reduce the incidence of HA disorders. 51 Taking all of the above findings into consideration, we concluded that the members of a population with a lower frequency of the DD genotype have lower ACE activity, angiotensin II production and bradykinin degradation; and higher Ang 1–7 production. This meta-analysis demonstrated that HA native populations have lower frequencies of the DD genotype, and that this condition aids in HA adaptation.
Based on the hypothesis by Montgomery et al. 33 and Qadar et al. 35 that the D allele is associated with serum ACE activity, individuals in populations with a higher frequency of the ACE DD genotype have a reduced ability to adapt to HA; however, Droma et al. 34 report that different ACE haplotypes did not affect the serum level of ACE, and that the serum level of ACE was not related to the D allele in Sherpas and non-Sherpas. The ACE DD genotype is also reported to be a risk factor for many HA illnesses. For example, the frequency of the ACE D allele is higher in Han individuals with acute mountain sickness (AMS) and in Tibetan residents with chronic mountain sickness (CMS). 52 Furthermore, Qi et al. 39 conducted a meta-analysis of five studies and found that carriers of the ACE D allele were at significant risk for developing HAPE.
This study has several limitations. First, only a small number of qualified studies were available, which prevented us from performing subgroup analyses to identify the potential effects on HA adaptation of other factors, such as gender and age; thus, linkage disequilibrium may exist and may be responsible for the observed variations in plasma ACE level. Second, the studies included in the meta-analysis investigated subjects residing at different altitudes; therefore, we just predict a trend about ACE genotype and HA adaptation. Third, the sample size was small, mainly due to the scarcity of existing publications regarding HA adaptation. Further studies should evaluate the associations between the ACE DD genotype and HA adaptation, using larger sample sizes or other populations. Finally, we explored the association between the ACE I/D polymorphism and HA adaptation, but did not evaluate whether other ACE polymorphisms were associated with HA adaptation. Therefore, the role of the ACE I/D polymorphism may be masked by the effects of other genes or by the environment.
Conclusions
In summary, we analyzed the association of an ACE polymorphism with HA adaptation. We found that the ACE DD genotype is a risk factor for HA maladaptation and that a lower frequency of the ACE DD genotype in a population indicates a greater ability of individuals in that population to adapt to HA.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the National Natural Science Foundation of China (No. 81571843).