
Abstract
Introduction:
Since the discovery of the (pro)renin receptor (PRR), it has been considered as a novel bioactive molecule of the renin–angiotensin system (RAS). The activation of PRR can elicit a series of angiotensin II (AngII)-independent effects.
Materials and methods:
In this study, we investigated the effects of prorenin and PRR on the proliferation of human umbilical artery smooth muscle (HUASM) cells and explored the possible mechanisms underlying these effects.
Results:
The binding of prorenin to PRR can promote proliferation and upregulate the anti-apoptotic protein Bcl-2 and downregulate the pro-apoptotic protein Bax independently of AngII in HUASM cells. In addition, the binding of prorenin to PRR can also increase the production of reactive oxygen species (ROS) and the phosphorylation of extracellular signal-regulated kinase (ERK1/2) independently of AngII. The pretreatment of HUASM cells with an NADPH oxidase inhibitor DPI decreased the production of ROS and also decreased the phosphorylation of ERK1/2. Furthermore, pretreatment of HUASM cells with DPI and the ERK1/2 inhibitor PD98059 significantly attenuated the prorenin-induced proliferation and regulation of apoptosis factors.
Conclusion:
Binding of prorenin to PRR can induce HUASM cell proliferation via the ROS generation and ERK1/2 activation.
Introduction
Since the function of the (pro)renin receptor (PRR) was proposed, 1 it has been considered as a new component of the renin–angiotensin system (RAS). Prorenin, which has been generally assumed to be an inactive precursor of renin, can bind to its cognate receptor, inducing a conformational change that renders it enzymatically active.1-4 Prorenin synthesis can occur at adrenal tissue, eye and ovaries,5-7 and the physiological concentration of prorenin in plasma is in the picomolar range. For as of yet unknown reasons, the concentration of prorenin in human plasma is higher than that of renin 8 and is 100 times higher in microalbuminuric diabetic subjects.5,9,10 High levels of prorenin in human plasma may indicate that the body is in a pathological condition, thus prorenin could be predictive of disease. The binding of prorenin to PRR not only increases the production of AngII11,12 but also activates signal transduction pathways that elicit fibrotic13,14 and proliferative effects15,16 independently of AngII. AngII is the key component of the RAS and can elicit proliferative, fibrotic and proinflammatory effects through a series of mechanisms that are commonly linked to the production of reactive oxygen species (ROS). ROS play an important role in cardiovascular diseases, such as hypertension, atherosclerosis and heart failure. 17 The struggle between proliferation and apoptosis of vascular smooth muscle cells (VSMCs), which participate in the initiation and early progression of atherosclerosis, 18 was demonstrated to be mediated by AngII through ROS production, which depends on the activation of NAD(P)H oxidase.19-23 However, whether prorenin could induce apoptosis in human umbilical artery smooth muscle (HUASM) cells has not been reported, and whether and how prorenin and PRR affect the production of ROS in HUASM cells remain unknown.
In this study, we hypothesized that the binding of prorenin to PRR can promote proliferation and inhibit apoptosis, and we investigated the role of ROS in the prorenin-induced effects on HUASM cells. To evaluate the PRR-mediated, AngII-independent effects of prorenin, we used Valsartan (Ang II type 1 receptor blocker) and PD123319 (Ang II type 2 receptor blocker) and PRR-targeted siRNA.
Materials and methods
Chemicals
Prorenin was purchased from Cayman Chemical Company (Michigan, USA). PD123319 and DPI were purchased from Sigma-Aldrich (Saint Louis, MO, USA). Valsartan was obtained from Beijing Novartis Company (Beijing, China). The ERK1/2 inhibitor PD98059 was purchased from Abcam (Cambridge, UK). PRR siRNA and transfection reagents were purchased from Invitrogen (California, USA).
Cell culture
Human umbilical cords were collected from healthy pregnant women at The First Affiliated Hospital of Dalian Medical University. The pregnant women gave written informed consent prior to delivery. This study was approved by the Ethics Committee of The First Affiliated Hospital of Dalian Medical University.
Under sterile conditions, umbilical arteries were isolated from human umbilical cords. The arteries were dissected longitudinally, and the intimas of the arteries were scraped with scissors to remove the endothelium. Next, the arteries were cut into small pieces of tissue (1×1×1 mm). The tissue samples were placed in 35 mm culture plates and incubated at 37°C and 5% CO2. After 2 h, 3 ml of Medium 231 (Gibco, California, USA) supplemented with Smooth Muscle Growth Supplement (SMGS, Gibco, California, USA) and 1× strength antibiotics/antimycotic (HyClone, Logan City, Utah, USA) was gently transferred into the culture plates. The media were refreshed every 3 days.
Cells began growing from the explants within 1 week and became confluent in approximately 3–4 weeks. The HUASM cells were cultured in Medium 231 with SMGS for trypsinization, passage, or freezing, according to standard protocols. We used second-to-fourth passage HUASM cells in our experiments.
Specific siRNA and transfection
The cDNA sequence of the PRR gene was obtained from GenBank (NM_005765.2), and the targeting sequences of the siRNAs were designed and chemically synthesized by Invitrogen (California, USA). The nucleotide sequences were as follows: 5’-GCUCCGUAAUCGCCUGUUU -3’ (sense), and 5’-AAACAGGCGAUUACGGAGC -3’ (antisense). As a negative control, a non-targeting scrambled siRNA (control siRNA) was also purchased. Cells were plated in 6-well plates, and the siRNAs were transfected into the cells using Lipofectamine RNAiMAX Reagent (Invitrogen, California, USA) according to the manufacturer’s protocols.
Immunofluorescence
HUASM cells were seeded into 24-well plates at 5000 cells per well and grown in Medium 231 supplemented with SMGS for 1–2 days. The cells were rinsed with phosphate-buffered saline (PBS), fixed in 4% paraformaldehyde (30 min, 4°C), washed three times with PBS, and permeabilized for 20 min with 0.3% Triton-X100. The cells were washed with PBS and then incubated at room temperature for 1 h in PBS with 5% bovine serum albumin (BSA). Labeling experiments were performed with a mouse anti-human antibody against α-smooth muscle actin (1:100, BOSTER, Wuhan, China) or PRR (1:100, Abcam, Cambridge, UK). Fluorescein-conjugated AffiniPure goat anti-mouse IgG antibodies (ZSGB-BIO, Beijing, China) or Rhodamine (TRITC)-conjugated AffiniPure goat anti-rabbit IgG antibodies (ZSGB-BIO, Beijing, China) diluted 1:100 in PBS (1 h, room temperature) were used for visualization. Hoechst 33258 (37°C, 5 min) was used to stain the cell nuclei. The cells were rinsed and assessed by fluorescence microscopy.
Measurement of cell proliferation
The proliferation of HUASM cells was measured using a Cell Counting Kit-8 (CCK-8) (Beyotime, Shanghai, China), according to the manufacturer’s protocol. Briefly, the HUASM cells were cultured in triplicate at 2×103 cells/well in 96-well plates. After different treatments, 10 μl of CCK-8 reagent was added to each well. After 1 h of incubation at 37°C, the absorbance at 450 nm was measured using a microplate reader. The relative levels of cell proliferation for each group were normalized to the levels of the control group, which represented 100%.
ROS measurement
ROS production was measured using a Reactive Oxygen Species Assay Kit (Beyotime, Shanghai, China). Briefly, 1×106 HUASM cells seeded into 6-well plates were treated with prorenin for 24 h. At the end of the treatment period, the cells were washed once with PBS and then incubated with DCFH-DA (5 μM) in medium for 20 min at 37°C in the dark. Next, the cells were washed with PBS three times, trypsinized, resuspended in PBS, and analyzed by flow cytometry. The fluorescence intensity of the cells was measured at 488 nm using a flow cytometer (BD FACS AriaII, USA).
Real-time PCR
Total RNA was extracted from cells using the TRIzol reagent (Invitrogen, California, USA) according to the manufacturer’s protocols. cDNA was synthesized from total RNA using the PrimeScript RT Master Mix (TaKaRa, Dalian, China). mRNA expression levels were evaluated by real-time PCR using a LightCycler (Roche Diagnostics) and SYBR Premix Ex Taq II (TaKaRa, Dalian, China). The amplification reactions were performed in a total volume of 20 μl and cycled 40 times after initial denaturation (95°C for 30 s) with the following parameters: 95°C for 5 s and 60°C for 20 s. GAPDH was used as the internal control. All of the primers were designed by TaKaRa (Table 1).
Primer series of Bcl-2, Bax, ATP6AP2 and GAPDH gene.

Western blot
Cellular proteins were extracted with lysis buffer (150 mM NaCl, 1% NP-40, 0.1% SDS, 2 mg/ml aprotinin and 1 mM PMSF) for 30 min at 4°C. The extracts were centrifuged at 12,000×g for 15 min at 4°C. Then, supernatants containing the total cellular protein were harvested. The protein concentrations were quantified using the BCA Protein Assay Kit (Keygen, Nanjing, China). The total protein extracts (30 μg) were electrophoretically separated using 10% SDS-polyacrylamide gels and transferred electrophoretically onto a polyvinylidene difluoride membrane (0.45 μm pore size). The blots were blocked for 60 min at room temperature with 5% skimmed milk in PBS with Tween-20. Immunoblotting was performed using anti-phospho-ERK 1/2, anti-ERK 1/2 (Bioworld Technology, Minneapolis, USA, dilution 1:750), anti-Bcl-2 (Santa Cruz, California, USA, dilution 1:200), anti-Bax (Santa Cruz, California, USA, dilution 1:500) and anti-PRR (Abcam, Cambridge, UK, dilution 1:700) antibodies overnight at 4°C. After three washes with PBST at 10 min, the membrane was incubated with horseradish peroxidase-conjugated secondary antibodies (dilution 1:1500, ZHONGSHAN, Beijing, China) for 1 h at room temperature. The blots were visualized using ECL Plus according to the manufacturer’s protocols. Blotting with an anti-β-actin (dilution 1:1000, Bioss, Beijing, China) antibody served as an internal control.
Statistical analysis
The data were subjected to a one-way analysis of variance (ANOVA) using SPSS 16.0 software, and the least significant difference (LSD) test or the Dunnett T3 test were used for post-hoc subgroup analysis. All of the data are expressed as the mean values ± SD of three independent experiments. The data were considered statistically significant when p < 0.05.
Results
Verification of vascular smooth muscle cells and the location PRR in HUASM cells
We observed HUASM cells that appeared fusiform or polygonal in shape by light microscopy (Figure 1(a)). Because smooth muscle cells express the smooth muscle myofilament protein α-smooth muscle actin (SMA), we evaluated α-SMA (Figure 1(b–d)) by fluorescence immunohistochemistry and observed the uniform and filamentous expression of α-SMA in the HUASM cells. We also determined the distribution of PRR in HUASM cells by fluorescence immunohistochemistry and detected the cytoplasmic expression of PRR (Figure 1(e–g)). We can see that the red fluorescence represents the PRR expression, and find that around the cell nucleus the PRR expression was higher.

Verification of vascular smooth muscle cells and the localization of PRR in HUASM cells.
Effect of prorenin on proliferation in HUASM cells
Cell proliferation was assayed using a CCK-8 assay. We treated HUASM cells with 0.1, 1 and 10 nM prorenin for 24 h in the presence of Valsartan and PD123319 and found that 10 nM prorenin most significantly induced the proliferation of HUASM cells independently of AngII (Figure 2(a)). Next, we observed the effect of prorenin on proliferation at different time points and found that with increasing time, the proliferative effect became more significant (Figure 2(b)).
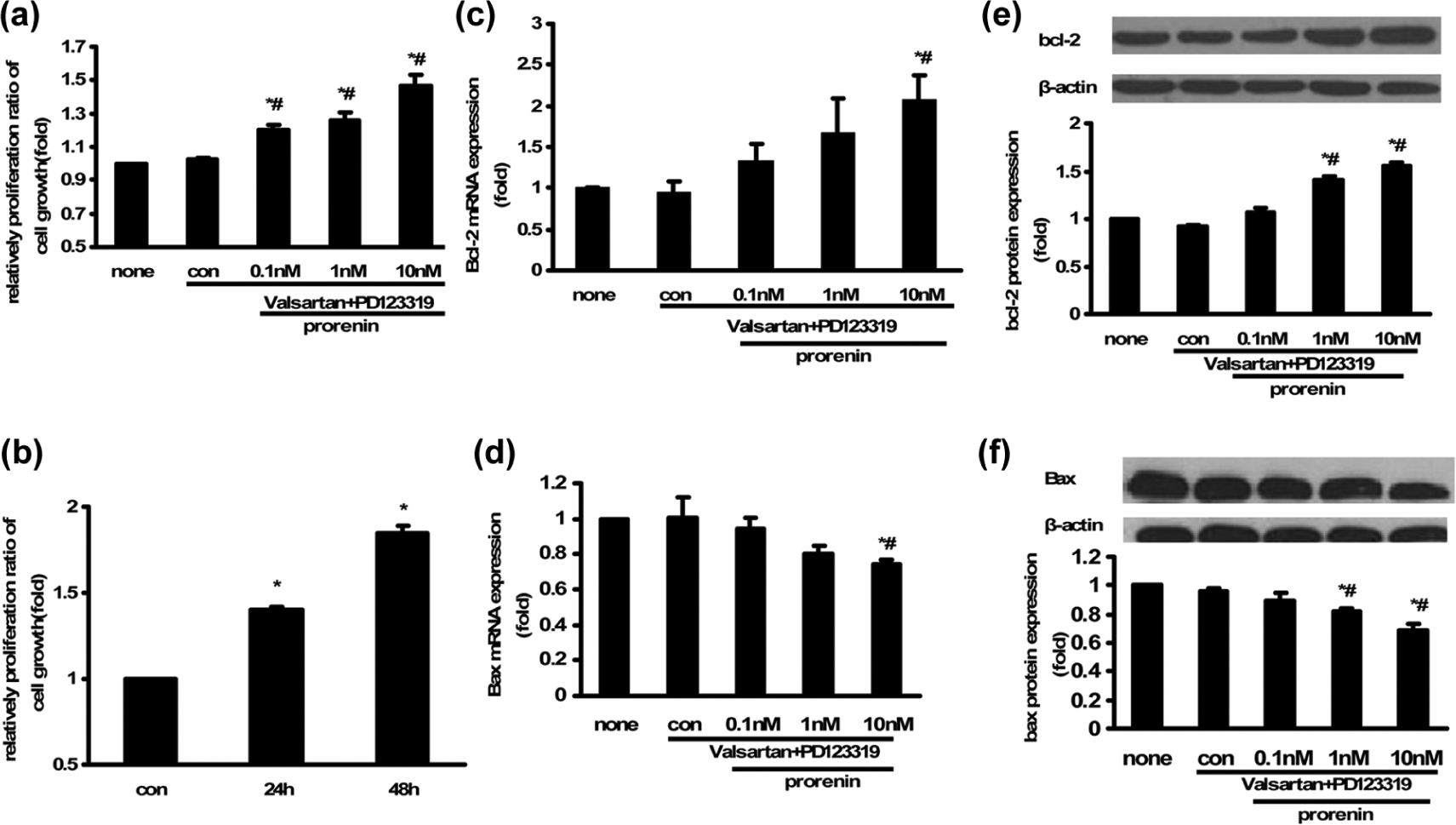
Effects of prorenin on the proliferation of HUASM cells.
We next determined whether prorenin could affect the expression of apoptosis-related factors. As shown in Figure 2, both protein and mRNA levels of anti-apoptotic Bcl-2 and pro-apoptotic Bax were upregulated and downregulated, respectively, in the 10 nM group compared with none and con group. These results indicated that prorenin can regulate the expression of Bcl-2 and Bax in HUASM cells.
Downregulation of PRR inhibits the prorenin-induced proliferation of HUASM cells
To determine whether the effects of prorenin are mediated by the PRR in HUASM cells, HUASM cells were transfected with PPR-targeting siRNA. At 72 hours after transfection, the levels of PRR mRNA were the most significantly reduced by 82% (Figure 3(a)), and the PRR protein was reduced to barely detectable levels (Figure 3(b)).
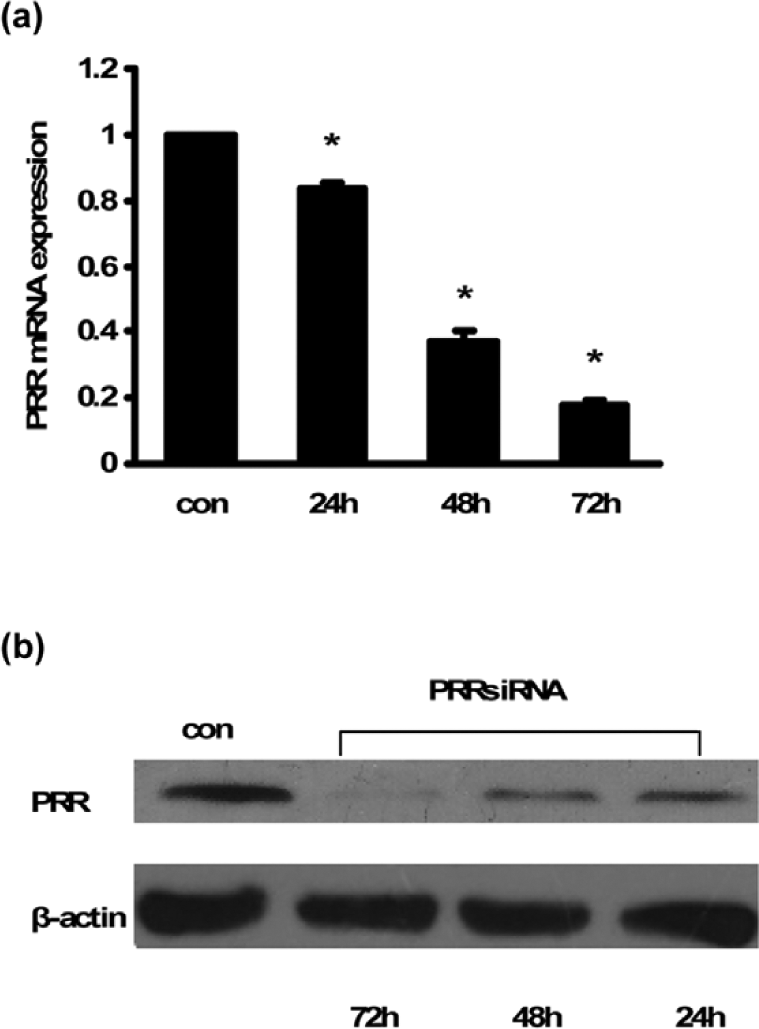
Effect of siRNA-mediated PRR knockdown assessed at the mRNA and protein levels.
Transfection with a PRR siRNA significantly decreased the effects that prorenin (10 nM at 24 h) elicited on the proliferation of HUASM cells (Figure 4(a)). The expression of Bcl-2 and Bax were also affected by PRR. As shown in Figure 4, the upregulated expression of Bcl-2 and the downregulated expression of Bax were altered, at both the protein and mRNA levels, after transfection with the PRR siRNA. These results indicated that the prorenin-induced effects in HUASM cells were mediated by PRR.

Downregulation of PRR inhibits the prorenin-induced proliferation of HUASM cells.
Prorenin promotes the production of ROS in HUASM cells
Although prorenin can promote the proliferation of HUASM cells, the underlying mechanism remains unclear. Thus, we first examined whether prorenin elicits an effect on the production of ROS in HUASM cells. We found that prorenin increases the production of ROS in a concentration-dependent manner and that the treatment with the nitric oxide synthase inhibitor DPI could inhibit this ROS production (Figure 5(a)). Transfection with a PRR siRNA significantly reduced the effects of prorenin (10 nM at 24 h) on the production of ROS (Figure 5(b)). These data indicated that prorenin can induce the production of ROS through PRR.

Effect of prorenin on the production of ROS and the phosphorylation of ERK1/2 in HUASM cells.
Effect of prorenin on ERK1/2 phosphorylation
Prorenin significantly increased the phosphorylation of ERK1/2 independently of AngII, with peaks at 15 min and 30 min compared with the control (Figure 5(c)). Furthermore, pretreatment with DPI (10 μM) decreased the prorenin-induced phosphorylation of ERK1/2 to a level that was similar to that observed with PD98059 (25 μM) (Figure 5(d)). These results indicated that ROS are involved in prorenin-induced phosphorylation of ERK1/2 in HUASM cells.
Prorenin promotes proliferation via the production of ROS and the activation of the ERK1/2 pathways
Prorenin can induce the production of ROS and the phosphorylation of ERK1/2 in HUASM cells, but whether ROS and ERK1/2 are required for prorenin-induced proliferation is unknown. As shown in Figure 6, pretreatment with DPI and PD98059 significantly inhibited the production of ROS, the phosphorylation of ERK1/2 and the proliferative effects of prorenin on HUASM cells. Moreover, the pretreatment also altered the prorenin-induced upregulated expression of Bcl-2 and the downregulated expression of Bax at both the protein and mRNA levels.

Effect of PD98059 and DPI on prorenin-induced proliferation in HUASM cells.
Discussion
In this study, we demonstrated that prorenin can promote the proliferation of HUASM cells and upregulate the expression of Bcl-2 and downregulate the expression of Bax through PRR independently of AngII. We also found that ROS and ERK1/2 are involved in these prorenin-induced effects.
First, we confirmed the expression of PRR in HUASM cells, consistent with previous studies.1,13,24 The functional PRR was first proposed by Nguyen et al., 1 who demonstrated that PRR is present in several organs, including the heart, brain, placenta, kidney and the liver. The role of PRR has been investigated in many cell and tissue types. The activation of PRR promotes the expression of fibrosis-related genes in HEK cells 14 and VSMCs,13,16 and induces oxidative stress in neuronal cells. 25 In animal models, PRR has been shown to be involved in the regulation of myocardial fibrosis and the deterioration of cardiac function. 26 In diabetic rats, the expression of PRR is upregulated 27 and contributes to the development of glomerulosclerosis,28,29 diabetic nephropathy, 30 and elevated blood pressure and heart rate. 31 Extensive evidence has shown that PRR is associated with cardiovascular disease. 32
Prorenin, which is considered as an inactive renin precursor, can bind to PRR within the nanomolar range1,2,33 and trigger a series of molecular events. The binding of prorenin to PRR might not only promote the production of AngII but also might play a role similar to, but independently of, AngII. In this study, we demonstrated that prorenin can promote the proliferation of HUASM cells through PRR independently of AngII and can further regulate the expression of Bcl-2 and Bax. Proliferation and apoptosis are imbalanced processes during the early progression of atherosclerosis.
VSMCs have two types of phenotype: synthetic phenotype and contractile phenotype. VSMCs undergo a phenotypic change from contractile phenotype to synthetic phenotype in response to external stimulation such as growth factors and shear stress. 34 This transformation directly causes abnormal proliferation and migration of VSMCs from the medial layer to the intimal layer, and VSMCs can synthesize extracellular matrix which is responsible for formation of atherosclerotic plaque. 18 Phenotype modulation of VSMCs contributes to the remodeling of vessels, which further leads to the occurrence of cardiovascular diseases. Apoptosis occurs at different stages of atherosclerosis. In the early progression of atherosclerosis, phenotype-transformed VSMCs may express adhesion molecules such as vascular cell adhesion molecule-1 and intercellular adhesion molecule-1 like endothelial cells, and these adhesion molecules could promote VSMC proliferation and also prevent apoptosis of these cells. 18 The Bcl-2 family proteins are important regulators of cellular apoptosis pathways, and Bcl-2 is an anti-apoptotic protein, whereas Bax is a pro-apoptotic protein. 35 These proteins contribute to the balance between proliferation and apoptosis. 36 Our study showed that prorenin can regulate the expression of apoptosis proteins through PRR independently of AngII in HUASM cells, and this anti-apoptotic effect may contribute to the formation of early atherosclerosis.
ROS are known to participate in the development of many cardiovascular diseases. 37 In VSMCs, various stimuli can induce the production of ROS, which might induce further proliferation, 38 increase inflammatory gene expression 39 and modulate matrix remodeling. 40 In endothelial cells, ROS can regulate the expression of adhesion molecules, including vascular cell adhesion molecule-1 and intracellular adhesion molecule-1. 41 The production of ROS, which can represent a second messenger, can trigger a series of downstream signal transduction pathways, such as those mediated by Akt 22 or ERK1/2.42,43 A weakened antioxidative defense system and oxidative stress can contribute to the dysfunction of vascular cells, increasing the relative inflammatory gene expression. 44 Based on the effect of ROS on VSMCs and endothelial cells, ROS can participate in the pathogenesis of atherosclerosis. Our study showed that prorenin can promote the production of ROS through PRR independently of AngII. In HEK 14 and neuronal 25 cells, ROS are also involved in prorenin-induced effects through PRR. In addition, the expression of PRR is regulated by ROS in diabetic rats. 27 In previous studies, the prorenin-induced phosphorylation of ERK1/2 has been shown in VSMCs.15,16,45 Our study further confirmed that the prorenin-induced phosphorylation of ERK1/2 is mediated by ROS production, and that the inhibition of ROS production reduced the phosphorylation of ERK1/2. Early studies have also reported that ROS can mediate ERK1/2 phosphorylation in response to AngII. 46 A recent study claimed that in neuronal cells, the production of ROS through PRR is regulated by ERK1/2 and PI3K/Akt. 25 There may be a crosstalk between ROS production and the phosphorylation of ERK1/2, which remains to be elucidated. Overall, ROS and ERK1/2 play pivotal roles in terms of prorenin-induced effects.
The RAS plays a key role in cardiovascular and renal diseases, and the blockade of the RAS is considered to be beneficial and to reduce cardiovascular risks, despite the fact that its inhibition cannot completely inhibit the development of these diseases. The answer to this question might reside in the AngII-independent effects induced by the binding of prorenin with PRR. Ichihara et al. 47 developed a decoy-epitope peptide of the prorenin prosegment, which is called the ‘handle-region peptide’(HRP) and which can block the binding of prorenin to the PRR. In in vivo studies, the subcutaneous administration of HRP significantly attenuated renal and cardiac damage.48,49 However, certain studies have failed to find any effects elicited by HPR in this regard.50,51 Although discrepancies remain regarding HRP, PRR is clearly a key component of the RAS, in addition to AngII, and PRR represents a potential pharmacological target.
In conclusion, our study has shown that prorenin can promote the proliferation of HUASM cells, upregulate the expression of Bcl-2 and downregulate the expression of Bax through PRR independently of AngII. Moreover, we demonstrated that ROS and ERK1/2 are involved in prorenin-induced effects.
Footnotes
Conflict of interest
None declared.
Funding
This work was supported by the National Natural Science Foundation of China (No.3027143).