
Abstract
Aim:
We investigated whether aliskiren, a direct renin inhibitor, provided protection in a model of diabetic nephropathy in mice and compared its protective effects to valsartan, an angiotensin II type 1 receptor blocker.
Materials and methods:
Hyperglycemia was induced with streptozotocin (STZ, 40 mg/kg/day × 5 days) injection in DBA/2J mice fed on a high fat diet. Mice were treated with either aliskiren (25 mg/kg/day) or valsartan (8 mg/kg/day) for 6 weeks.
Results:
Aliskiren and/or valsartan treatment significantly attenuated albuminuria, urinary nephrin excretion and glomerulosclerosis. Aliskiren and/or valsartan prevented reduction of podocin and WT1 protein abundance in diabetic mice. Aliskiren and/or valsartan significantly prevented increased expression of profibrotic growth factors (TGFβ, CTGF and PAI-1), proinflammatory cytokines (MCP-1, TNFα and IL-1β), endoplasmic reticulum (ER) stress markers (CHOP and XBP-1) and lipid accumulation in the kidney of diabetic animals. Aliskiren showed similar efficacy compared to valsartan therapy and dual treatment in some aspects has synergistic protective effects.
Conclusion:
Our study indicates that aliskiren and/or valsartan protects against diabetic kidney disease through multiple mechanisms, including decreasing podocyte injury, activation of profibrotic growth factors and proinflammatory cytokines, ER stress and accumulation of lipids.
Introduction
Diabetic nephropathy (DN) is the most common renal complication of diabetes mellitus and a leading cause of kidney failure, accounting for 44% of all new cases of kidney failure in the United States in 2008. 1 DN is characterized by functional and structural changes in the kidney such as progressive proteinuria, glomerulosclerosis and tubulointerstitial fibrosis, with over-activation of the renin-angiotensin-aldosterone system (RAAS) playing an important role. 2
Currently, drugs targeting RAAS, including angiotensin-converting enzyme inhibitors (ACEIs), angiotensin II (AngII) type I receptor (AT1R) blockers (ARBs) and aldosterone blockers have been most widely used for slowing down or preventing renal damage in diabetes. However, they are not completely effective in preventing or reversing the development of DN3,4, in part due to a compensatory increase in plasma renin activity (PRA) and AngII, which may potentially override ARB blockade of AT1R. Inhibition of renin, a rate-limiting step in the RAAS activation cascade, provides an additional approach and the potential for achieving a full blockade of RAAS activity by suppressing production of AngI and AngII; meanwhile, renin inhibition may be also associated with reduced production of angiotensin 1-7 [Ang-(1-7)], as a positive component in RAAS cascade, antagonizing effects of angiotensin II in the diabetic kidneys.
Aliskiren as a direct renin inhibitor through combination with other RAAS blockade has shown protective renal and cardiac effects in human and animal diabetes.5–10 Work using mouse models including a eNOS deficient model, db/db and streptozotocin (STZ)-induced type I DN supports that combination of aliskiren and valsartan led to greater reduction in proteinuria and prevented glomerular and tubular injury compared to either intervention alone. However, in a more recent study, in transgenic Ren2 rats with increased AngII and hypertension, combination therapy did not result in better improvement in markers of glomerular injury when compared with single treatment. 11 Similarly, renin inhibition showed equally effective nephroprotection as AT1 receptor antagonism does in mice with chronic kidney disease, without the additional benefit of combination therapy. 12 The reasons for the observed differences among these studies are unclear so far.
Accumulation of excess lipids in non-adipose tissues, including the kidney, may induce cellular dysfunction and may play an important role in the pathogenesis of diabetic kidney injury.13,14 Our previous studies have demonstrated that dysregulated lipid metabolism including increased fatty acid and cholesterol synthesis and/or reduced fatty acid oxidation and/or cholesterol efflux contributes to lipid accumulation in the diabetic kidney.15–21 Direct renin inhibition has been shown to improve insulin resistance and adipose tissue dysfunction in diabetic KK-Ay mice 22 but whether RAAS inhibition by aliskiren can improve abnormal lipid metabolism in the diabetic kidney is not known.
The current work extends studies by comparing the potential beneficial effects of aliskiren with or without valsartan in mice with DN. Here we demonstrate that aliskiren has equal efficacy as valsartan in ameliorating diabetic kidney diseases, by decreasing fibrosis, inflammation, endoplasmic reticulum (ER) stress and lipid accumulation. Combination with aliskiren and valsartan in some, but not in all aspects, showed better nephroprotective effects in diabetic kidneys compared with monotherapy.
Methods and materials
Animals and treatments
All procedures were in accordance with institutional guidelines for animal research. Eight week-old male DBA2/J mice were obtained from Jackson Laboratories (Bar Harbor, ME, USA). They were maintained on a 12-h light/12-h dark cycle. Mice were injected with freshly made STZ (Sigma-Aldrich, St. Louis, MO, USA, 50 mmol/L sodium citrate buffer pH 4.5) intraperitoneally (40 mg/kg) for 5 consecutive days, or with 50 mmol/l sodium citrate solution only. Tail vein blood glucose levels were measured 1 week after the last STZ injection, and mice with non-fasting glucose levels >250 mg/dl were considered diabetic. To replicate the hyperglycemia and hyperlipidemia seen in subjects with type 2 diabetes mellitus all mice were fed a high fat diet (21% milk fat, 0.15% cholesterol, TD88137, Harlan-Teklad, Madison, WI, USA) after the onset of diabetes. 19 Animal studies and relative protocols were approved by the Animal Care and Use Committee at the University of Colorado, Denver, USA.
The mice were assigned to five treatment groups (n=6 each group). They were treated with vehicle (controls (CTL) and diabetes (DM)), aliskiren (25 mg/kg per day, D-Ali), valsartan (8 mg/kg per day, D-Val) and combination (D-AV). Aliskiren was subcutaneously infused to mice via ALZET micro-osmotic pump (DURECT Co., Cupertino, CA, USA). Valsartan was given to mice in drinking water. The drug treatments were carried out for 6 weeks. A 24-h urine sample was collected from each mouse placed in individual metabolic cages to measure urinary albumin and nephrin excretion before sacrifice. Blood was taken from the tail vein in mice for measurement of serum glucose concentration. After 6 weeks of drug treatment, all mice were anesthetized with pentobarbital and the kidneys were rapidly excised from mice to perform biochemical and histological examinations, as described in following sections. Valsartan and aliskiren were kindly provided by Novartis (Basel, Switzerland).
Blood and urine chemistry
The serum concentration of glucose was measured by LabAssay Glucose Kit (Wako, Osaka, Japan). Urinary albumin, nephrin and creatinine concentrations were determined using kits from Exocell (Philadelphia, PA, USA).
Blood pressure measurement
Mean blood pressure (MAP) was measured in conscious mice by tail-cuff sphygmomanometer (Visitech BP2000; Apex, NC, USA). All animals were preconditioned for blood pressure measurements for 1 week before each experiment.
Histology
Sections (4-μm thick) cut from 10% formalin-fixed, paraffin-embedded kidney samples were stained with periodic acid-Schiff (PAS). Thirty glomeruli were randomly selected from each mouse to perform the PAS analysis. Semi-quantitative scoring of glomerulosclerosis was performed using a five-grade method described previously. 19 Frozen sections were stained with oil red O, to identify neutral lipid deposits.
Immunofluorescence microscopy
We performed immunostaining for nephrin (mouse antibody, a gift from Dr. Larry Holzman, University of Pennsylvania, Philadelphia, PA, USA) and synaptopodin (Sigma) and imaged with a laser scanning confocal microscope (Zeiss 510 NLO/META, Jena, Germany). The expression level was quantified as the sum of pixel values per glomerular area using ImageJ (version 1.44) image analysis software.
RNA extraction and quantitative real-time PCR
Total RNA was isolated from the kidney cortex using SV Total RNA Isolation System from Promega (Madison, WI, USA) and cDNA was synthesized using reverse transcriptase reagents from Bio-Rad Laboratories (Hercules, CA, USA). The mRNA level was quantified using Bio-Rad iCyCler real-time PCR machine. Cyclophilin RNA was used as an internal control and the amount of RNA was calculated by the comparative threshold cycle method as recommended by the manufacturer.
Protein electrophoresis and Western blotting
Equal amounts of protein samples from the kidney cortex were subjected to SDS-PAGE, and then transferred to nitrocellulose membranes. After being blocked with 5% fat-free milk powder in 0.1% Tween 20 in Tris-buffered saline (20 mmol/l Tris•HCl and 150 mmol/l NaCl, pH 7.4), the blots were incubated with antibodies against: 1) podocin (1:200; G20; Santa Cruz Biotechnology, Santa Cruz, CA, USA), 2) WT1 (1:200; G20; Santa Cruz Biotechnology), 3) MAS receptor (1:1000, Alomone Labs AAR-013, Jerusalem), 4) CHOP (1:1000, Cell signaling #2895), XBP-1 (1:1000, sc-7160). Corresponding secondary antibodies were visualized using enhanced chemiluminescence (Pierce, Bradford, IL, USA). The signals were quantified with a chemiluminescence detector and the accompanying densitometry software (UVP, Upland, CA, USA).
Statistical analysis
Results are presented as the means ± SE. Data were analyzed by ANOVA and Student-Newman-Keuls test for multiple comparisons. Statistical significance was accepted at the P < 0.05 level.
Results
Treatment of diabetic mice with aliskiren and/or valsartan on metabolic parameters
As shown in Table 1, the body weights of diabetic mice were significantly decreased compared to control mice, whereas the ratio of kidney weight/body weight was significantly increased in all groups of diabetic mice. Blood glucose levels in all diabetic groups were significantly higher than non-diabetic control mice. Aliskiren or valsartan treatment had no effect on the blood glucose levels in these mice. Therapy with aliskiren, valsartan and the combination all significantly reduced blood pressure in diabetic mice compared with non-treated diabetic mice.
Metabolic data.

Values are means ± SE; D-Ali: diabetes with aliskiren treatment alone; D-Val: diabetes with valsartan treatment alone; D-AV: diabetes with aliskiren and valsartan treatment; KW: kidney weight; BW: body weight.
p<0.05 when compared with control; bp<0.05 when compared with diabetes; cp<0.05 when compared with aliskiren or valsartan groups. n=6 in each group.
Treatment of diabetic mice with aliskiren and/or valsartan improved proteinuria and prevented podocyte injury and loss as well as mesangial expansion
As shown in Figure 1(a), diabetic mice treated with vehicle developed moderate albuminuria. Monotherapy with aliskiren and/or valsartan significantly and comparably decreased urinary albumin excretion in diabetic mice. Podocyte injury results in increased podocyte excretion into the urine. We therefore measured urinary nephrin excretion as a marker of podocytes in the urine. Although both aliskiren and valsartan treatment alone significantly reduced urinary nephrin excretion in diabetic mice, combination therapy provided greater reduction in urinary nephrin excretion than single treatment, indicating synergistic effects of aliskiren and valsartan on protection of podocyte integrity in diabetes (Figure 1(b)). Consistent with reduced urinary nephrin excretion, reduced immunostaining of nephrin in glomeruli of diabetic mice was significantly prevented by both monotherapy and combination treatment (Figure 2(a) and 2(c)). Immunofluorescence microscopy showed that both monotherapy and combination treatment also prevented reduced expression of synaptopodin seen in mice with diabetes (Figure 2(b) and 2(c)). Western blots showed that podocin protein abundance was 40% decreased in diabetic mice treated with vehicle, monotherapy with aliskiren or valsartan and combination treatment reversed decreased podocin expression to control level. Although combination treatment was associated with higher expression of podocin when compared to monotherapy, this increase did not reach statistical significance (Figure 3(a) and 3(c)). Protein expression of WT1, a nuclear podocyte marker, was also examined by Western blots (Figure 3(b) and 3(c)). The abundance of WT1 was significantly reduced to about 80% of control level in diabetic mice, whereas monotherapy and combination treatment increased WT1 expression, suggesting podocyte loss seen in diabetic mice may be prevented by the treatments.

Effects of aliskiren and/or valsartan treatment on urinary albumin and nephrin excretion. (a): Urinary albumin excretion expressed as urinary albumin-to-creatinine ratio. (b): Urinary nephrin excretion expressed as urinary nephrin-to-creatinine ratio. *p<0.05 compared with controls (CTL); #p<0.05 compared with non-treated diabetes (DM); &p<0.05 compared with either D-Ali or D-AV; diabetes with aliskiren and valsartan treatment; D-Ali: diabetes with aliskiren treatment alone; D-Val: diabetes with valsartan treatment alone.

Aliskiren and/or valsartan treatment prevented reduction of nephrin and synaptopodin immunostaining in the kidneys of diabetic mice. (a): Immunofluorescence staining of kidney sections for nephrin. (b): Immunofluorescence staining of kidney sections for synaptopodin. (c): Relative staining intensity of nephrin and synaptopodin. *p<0.05 compared with controls (CTL); #p<0.05 compared with non-treated diabetic mice (DM); &p<0.05 compared with aliskiren (D-Ali) or valsartan treated mice (D-Val), D-AV: diabetes with aliskiren and valsartan treatment.

Aliskiren and/or valsartan treatment prevented reduction of podocin and WT1 protein abundance in the kidneys of diabetic mice. (a) and (b): Semi-quantitative immunoblots reacted with anti-podocin (a) and anti-WT1 (b) antibodies; (c): Corresponding densitometric analyses of the data of podocin and WT1. *p<0.05 compared with controls (CTL); #p<0.05 compared with non-treated diabetic mice (DM), D-Ali: diabetes with aliskiren treatment alone; D-Val: diabetes with valsartan treatment alone; D-AV: diabetes with aliskiren and valsartan treatment.
In addition to podocyte injury, glomerular mesangial expansion is also a hallmark of DN. PAS staining and semiquantitative scoring was used to assess mesangial expansion. Compared with controls, diabetic mice with vehicle treatment showed marked mesangial expansion and clearly increased accumulation of extracellular matrix in the mesangium (Figure 4). Mesangial expansion was reduced by aliskiren or valsartan treatment alone, but combination with aliskiren and valsartan had additional inhibitory effects (Figure 4). Vehicle treated mice showed the highest score, and all drug treatments led to a reduction in the mesangial expansion, with combination treatment resulting in additional reduction (Figure 4).

Aliskiren and/or valsartan treatment improved morphological changes in the kidneys of diabetic mice. (a): Representative periodic acid-Schiff (PAS) staining of kidney sections. (b): Mesangial expansion index was defined as the ratio of mesangial area to glomerular tuft area, using a 5 point scale. *p<0.05 compared with controls (CTL); #p<0.05 compared with non-treated diabetic mice (DM);&p<0.05 when compared with either D-Ali or D-Val. D-Ali: diabetes with aliskiren treatment alone; D-Val: diabetes with valsartan treatment alone; D-AV: diabetes with aliskiren and valsartan treatment.
Treatment of diabetic mice with aliskiren and/or valsartan modulated renal fibrosis and inflammation
Renal fibrosis and inflammation are important mediators of DN. To determine the effect of the treatments on profibrotic factors, we measured the mRNA expression of transforming growth factor-β (TGF-β), plasminogen activator inhibitor-1 (PAI-1) and connective tissue growth factor (CTGF) in the kidney. As shown in Figure 5, in non-treated diabetic mice gene expression of TGF-β, PAI-1 and CTGF were all increased compared with the non-diabetic control. The increased mRNA level in these factors was significantly suppressed by the treatments with monotherapy and combination therapy. Combination therapy with aliskiren and valsartan remarkably decreased mRNA expression of the proinflammatory chemokines monocyte chemotactic protein-1 (MCP-1), tumor necrosis factor-α (TNF-α) and interleukin 1β (IL-1β) (Figure 5).
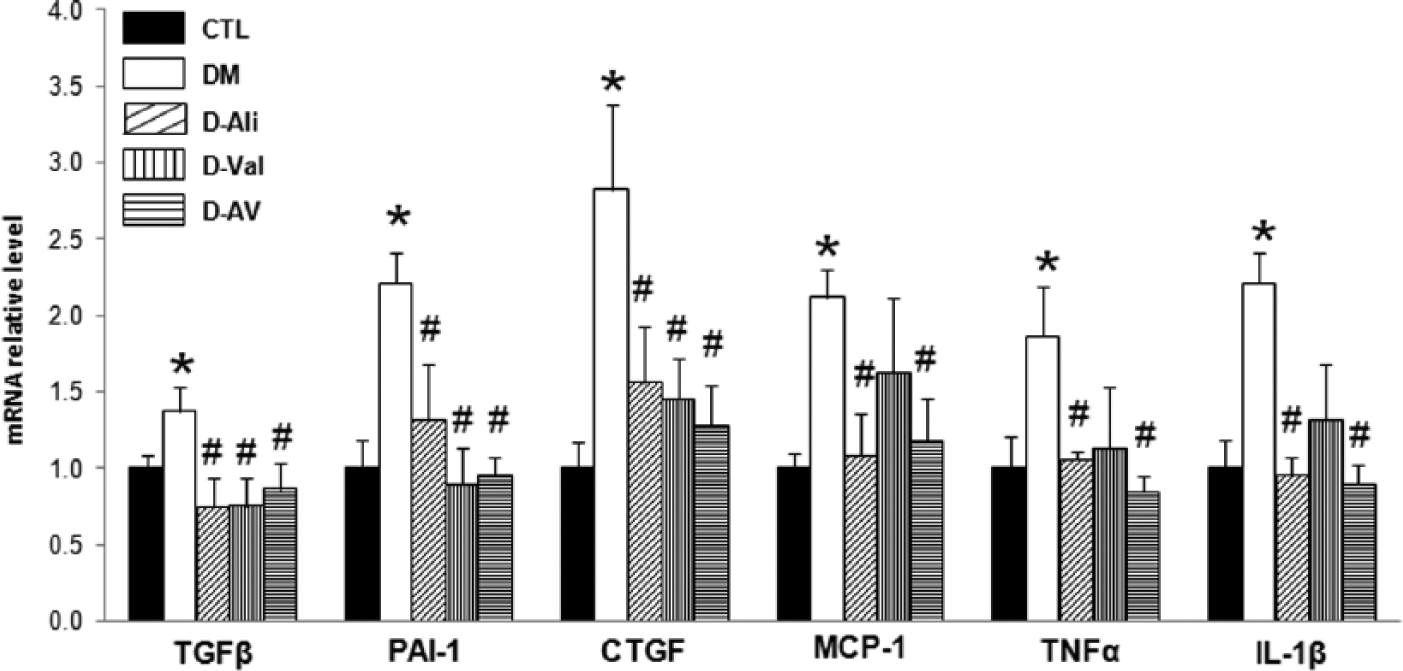
Aliskiren and/or valsartan treatment prevented activation of profibrotic factors and proinflammatory factors in the kidneys of diabetic mice. Analysis of mRNA expression by quantitative real-time PCR for TGFβ, PAI-1, CTGF, MCP-1, TNFα, and IL-1β. *p<0.05 compared with controls (CTL); #p<0.05 compared with non-treated diabetic mice (DM), D-Ali: diabetes with aliskiren treatment alone; D-Val: diabetes with valsartan treatment alone; D-AV: diabetes with aliskiren and valsartan treatment.
Combination with aliskiren and valsartan prevented renal lipid accumulation
Altered regulation of renal lipid metabolism has been shown to play an important role in the pathogenesis of DN. Oil red O staining showed increased kidney neutral lipid accumulation in both glomeruli and tubulointerstitium in diabetic mice. The lipid accumulation was moderately reduced after aliskiren and valsartan treatment (Figure 6(a)) and this was consistent with decreased mRNA expression of adipophilin, a marker of lipid droplets (Figure 6(b)) after combination treatment. We examined the expression of genes that mediate cholesterol and fatty acid/triglyceride synthesis. We found that there was a 2-fold increase in acyl-coenzyme A diacylglycerol acyltransferases (DGAT1) and a 12-fold increase in HMG CoA synthase (3-hydroxy-3-methylglutaryl-CoA synthase) (Figure 6(b)) protein expression in diabetes, which were markedly inhibited by combination treatment with aliskiren and valsartan, but not by single treatment. In contrast to DGAT1, there were no changes in DGAT2 mRNA levels (not shown) among the groups.

Aliskiren and or valsartan treatment prevented increase of lipid accumulation in the kidneys of diabetic mice. (a): Oil red O staining of kidney frozen sections. (b): Analysis of mRNA expression by quantitative real-time PCR for adipophilin, DGAT1 and HMG CoA synthase. *p<0.05 compared with controls (CTL); #p<0.05 compared with non-treated diabetes (DM), D-Ali: diabetes with aliskiren treatment alone; D-Val: diabetes with valsartan treatment alone; D-AV: diabetes with aliskiren and valsartan treatment.
Effects of combination with aliskiren and valsartan on ER stress
Diabetic kidney disease is associated with activation of ER stress response; we therefore investigated protein expression of two important markers of ER stress response: CHOP (C/EBP homologous protein) and XBP-1 (X-box binding protein1). Western blots showed 1.5-fold increases of CHOP and XBP-1 protein expression in diabetic mice, which was completely suppressed by aliskiren, valsartan or combination treatment at the same extent (Figure 7).

Aliskiren and/or valsartan treatment prevented increased protein abundance of CHOP and XBP-1 in the kidney cortex of diabetic mice. *p<0.05 compared with controls (CTL); #p<0.05 compared with diabetic mice (DM), D-Ali: diabetes with aliskiren treatment alone; D-Val: diabetes with valsartan treatment alone; D-AV: diabetes with aliskiren and valsartan treatment.
Effects of combination with aliskiren and valsartan on the RAAS
As shown in Table 2, intrarenal renin mRNA was dramatically induced in mice treated with aliskiren and/or valsartan, due to disruption of the feedback loop, whereas (pro)renin receptor expression was not significantly changed among groups. We also examined the other components of the RAAS. The renal expression of ACE and ACE2 mRNA were significantly reduced in diabetic mice; combination treatment was associated with recovery of their mRNA expression. Interestingly, Mas receptor mRNA and protein abundance was significantly decreased in diabetic mice and combination treatment with aliskiren and valsartan dramatically reversed Mas receptor mRNA and protein abundance to non-diabetic control level (Table 2 and Figure 8). Single treatment with aliskiren or valsartan had no effects on Mas expression.
Effects of aliskiren and valsartan on RAAS mRNA expression.

Values are means ± SE.
D-Ali: diabetes with aliskiren treatment alone; D-Val: diabetes with valsartan treatment alone; D-AV: diabetes with aliskiren and valsartan treatment; ACE: angiotensin-converting enzyme; ACE2: angiotensin-converting enzyme 2; AT1R: angiotensin II type 1 receptor; MR: mineralcorticoid receptor; AT2R: angiotensin II type 2 receptor; Mas: angiotensin 1-7 mas receptor.
p<0.05 when compared with control; bp<0.05 when compared with diabetes; cp<0.05 when compared with aliskiren or valsartan groups.

Diabetes was associated with reduced renal Mas receptor protein expression. Combination treatment significantly increased abundance of Mas receptor protein. Western blots (a) for Mas receptor and corresponding densitometry analysis (b). *p<0.05 compared with controls (CTL); #p<0.05 compared with non-treated diabetic mice (DM); &p<0.05 compared with aliskiren (D-Ali) or valsartan (D-Val) treated mice, D-AV: diabetes with aliskiren and valsartan treatment.
Discussion
In the present study, we demonstrated that aliskiren treatment significantly ameliorated kidney injury in diabetic mice. Indeed, aliskiren was equally beneficial as valsartan in protecting kidney injury in diabetes, including improvement of functional and structural changes and inhibition of fibrosis, inflammation, lipid accumulation and ER stress in the kidney. Dual therapy with aliskiren and valsartan, in some, but no in all aspects, produced additive and synergistic therapeutic effects in the treatment of DN.
Aliskiren and valsartan equally reduced blood pressure, albuminuria and glomerulosclerosis in the diabetic animals. Albuminuria is regarded as a major parameter of monitoring the progression of DN and the pathogenesis of albuminuria is mainly attributed to the alteration of glomerular filtration barrier.19,23 The development of albuminuria in the diabetic mice was associated with damage to podocyte foot processes. There were decreases in podocyte abundance and distribution, and in slit diaphragm protein expression, which were manifested by reduced expression of nephrin, synaptopodin, podocin and WT1 in diabetic animals. Interestingly, monotherapy with aliskiren or valsartan restored distribution and staining intensity of nephrin and synaptopodin and abundance of podocin and WT1 to a similar extent, suggesting a protective effect on structure of podocytes. Dual treatment with aliskiren and valsartan had better therapeutic effects on restoration of nephrin (glomerular slit diaphragm) and synaptopodin (actin cytoskeleton) protein expression, although it had no greater effect on albuminuria.
Although dual treatment was associated with a further decrease of blood pressure when compared with either single therapy group, animal studies in type II diabetic models (db/db mice), 9 STZ-induced diabetic rats 5 and the AVOID clinical study24–26 have demonstrated that the anti-proteinuric effects of aliskiren with or without the AT1 receptor antagonist were independent of its blood pressure lowering effect, suggesting antagonism to the intra-renal RAAS with aliskiren or valsartan potentially playing an important role in kidney protection. Functional expression of key components of the RAAS has been demonstrated in differentiated human podocytes27,28 and these podocytes are able to secrete AngII locally. It is therefore likely that local RAAS blockade in the kidney may prevent AngII-induced damage of podocytes in diabetic conditions.
Renal damage in diabetes is associated with marked increases in gene and protein expression of profibrotic and proinflammatory factors. TGFβ1 stimulates extracellular matrix production and is a key growth factor in diabetic glomerulosclerosis.29,30 TGFβ1 is also a downstream target of AngII 31 and can be up-regulated by renin in mesangial cells through the pro(renin) receptor independently of AngII.32–34 Treatments with aliskiren or valsartan significantly reduced the gene expression of TGFβ, CTGF and PAI-1 therefore preventing kidney injury in diabetes. In addition, at the doses used in this study aliskiren demonstrated a better effect than valsartan in preventing increase of proinflammatory cytokines TNF-α, MCP-1 and IL1-1β in the kidneys of the diabetic mice. A recent study performed a dose-response curve of RAAS antagonism with aliskiren and losartan, and showed much higher doses of aliskiren (50 mg/kg/day) and losartan (167mg/kg/day) than our studies, to yield equal RAAS inhibition. 12 Due to such a difference in dose response, we need to be cautious before we can conclude if the two treatments have different effects or additive effects on target pathways.
One of novel findings in the present study is the improvement in renal lipid accumulation by aliskiren and/or valsartan therapy. There is growing evidence for the role of abnormal lipid metabolism in the pathogenesis of renal disease.15–17,19–21,35 Lipid metabolites can induce profibrotic and proinflammatory cytokine production in the kidney cells, increase generation of reactive oxygen species and promote the expression of extracellular matrix.14,16,19,20,35–37 Oil red O staining showed neutral lipid accumulation (cholesterol ester and triglycerides) in the diabetic kidney. The increase in lipid staining was paralleled with increases in the mRNA level of DGAT1 that catalyzes terminal and the only committed step in triglyceride synthesis, 38 and increased mRNA expression of HMG CoA synthase that is involved in cholesterol synthesis. 39 Although aliskiren or valsartan prevented an increase of these two gene expression, dual treatment with aliskiren and valsartan indeed induced improvement in the alterations of renal lipid accumulation (oil red o staining) which is associated with reduced mRNA levels in DGAT1 and HMG CoA synthase. It has been known that AT1 receptor blockers can prevent atherosclerosis and decrease cholesterol and triglyceride levels, suggesting an interaction between the RAAS and lipid metabolism.40–42 Therefore blockade of the RAAS system with both aliskiren and valsartan, at least in part, protects renal function by preventing renal lipid accumulation.
The ER is the principal site for the folding and maturation of secretary and transmembrane proteins and the main site for the synthesis and uptake of lipids. Aberrant metabolic conditions such as hyperglycemia and hyperlipidemia can affect ER trafficking, activate ER stress and corresponding three signaling pathways that generally relieve the accumulation of misfolded or unfolded ER proteins. Recently, activation of ER stress response has been documented in diabetic animals and patients with progressive DN (e.g. increased expression of CHOP and XBP-1).43–45 Although the role of a classical marker of ER stress C/EBP homologous protein (CHOP), a transcription factor induces apoptosis, in kidney injury is debatable, 46 CHOP deficient mice were resistant to ER stress-induced cell death and DN, 45 and consistent with this, expression of CHOP protein was significantly increased in diabetic mice when compared with controls. In the present study aliskiren with or without valsartan reduced CHOP and XBP-1 protein expression, thus may attenuate ER stress response and prevent cells from injury and apoptosis in diabetic kidneys. However, how intracellular RAAS blockade by aliskiren or valsartan and how renin or AngII was involved in ER stress in diabetic kidneys are still unknown. Interestingly, in contrast, tunicamycin induced-ER stress is associated with increased activation of SREBP-2, a lipogenic transcription factor, and increased do novo biosynthesis and accumulation of lipid in the proximal tubules. 47 Further studies are therefore warranted to clarify the complex, mechanistic link between ER stress and lipid accumulation in the kidneys.
It is well known that angiotensin-converting-enzymes 2 (ACE2) converts AngII to Ang-(1-7) and Ang-(1-7) exerts its effects on antagonizing AngII in the kidney through Mas receptor.48,49 ACE2 gene deletion was recently found to accentuate diabetes-related glomerular damage 50 whereas ACE2 gene overexpression was found to ameliorate glomerular injury in DN. 51 Interestingly, Ang-(1-7) improved lipid and glucose metabolism in transgenic rats with increased circulating Ang-(1-7). 52 In our studies single treatment with aliskiren or valsartan did not prevent decrease of Mas receptor expression in diabetic mice, but dual treatment with aliskiren and valsartan significantly increased expression of Mas receptor mRNA and protein in diabetic mice, indicating that dual or a more complete blockade of RAAS has an effect on activating the ACE2-Ang-(1-7)-Mas receptor axis. Although more complete blockade of the RAAS with renin inhibitor and ARBs have shown beneficial effects in renal protection in diabetes, in clinical practice, these were offset by a high risk of hyperkalemia and hypotension, 53 which may be caused by suppression of aldosterone and alteration of renal hemodynamics. Whether the lack of protective components of RAAS cascade (such as ACE2-Ang-(1-7)-Mas receptor axis) is involved in such a consequence during dual blockade is still unknown. By inhibiting renin, aliskiren suppresses production of ANG I (a substrate of ACE2) resulting in a decrease of Ang-(1-7), as seen in an experimental chronic allograft nephropathy. 54 This showed better protective effects of candesartan than aliskiren, which was likely to be due to lower generation of protective Ang-(1-7) in response to aliskiren when compared with candesartan. Increased protein expression of Mas in dual treatments may be a feedback to complete blockade of RAAS, but it may suggest a potentially beneficial approach by combination with activation of Ang-(1-7) and inhibition of AngII in diabetic kidney diseases.
In conclusion, the direct renin inhibitor aliskiren provided a protective effect against renal damage in diabetic mice through inhibition of the profibrotic processes, inflammation and lipid accumulation, as well as prevention of the activation of ER stress. Aliskiren showed equal therapeutic efficacy when compared with valsartan, and thus the potential of aliskiren is at least comparable to that of valsartan in renoprotection. For certain targets, dual blockade with aliskiren and valsartan produced additive therapeutic effects in preventing the activation of cellular processes associated with the pathogenesis and progression of DN.
Footnotes
Acknowledgements
We thank Novartis Pharmaceuticals, Basel, Switzerland for kindly providing aliskiren and valsartan.
Imaging experiments were performed in the University of Colorado Anschutz Medical Campus Advance Light Microscopy Core supported in part by NIH/NCRR Colorado CTSI Grant Number UL1 RR025780.
We are grateful to Yu Lin and Renfei Luo (Institute of Hypertension and Kidney Research, Sun Yat-sen University, Guangzhou, China) for technical assistance.
Conflict of interest
None declared.
Funding
This work was supported by grants from Grants from Novartis Medical School IISS and NIH R01 AG026529. WW was supported by Novartis grants. This work was also supported by the Natural Science Foundation of China (Grant No. 81370822) and Guangdong Nature Science Foundation (Grant No. S2013010015499).