
Abstract
Introduction:
Angiotensin II (AngII) elicits smooth muscle contractions via activation of AngII type 1 receptor (AT1R) in the intestinal wall and in sphincter regions in several species. Achalasia is a rare swallowing disorder and is characterized by a loss of the wave-like contraction that forces food through the oesophagus and a failure of the lower oesophageal sphincter to relax during swallowing.
Aims and methods:
The present study was undertaken to elucidate expression and distribution of a local renin-angiotensin system (RAS) in the muscular layer of distal normal human oesophagus as well as in patients with achalasia using western blot analysis, immunohistochemistry and polymerase chain reaction (PCR).
Results:
AT1R, together with enzyme renin and cathepsin D expression were decreased in patients with achalasia. In contrast, the mast cells chymase, cathepsin G, neprilysin and the receptor for angiotensin 1–7 peptides, the MAS receptor, were increased in patients with achalasia.
Conclusion:
The results showed the existence of a local RAS in human oesophageal muscular layer. The enzymes responsible for AngII production are different and there has been a shift in receptor physiology from AT1R to MAS receptor in patients with achalasia. These changes in the RAS might play a significant role in the physiological motor control for patients with achalasia.
Keywords
Introduction
Achalasia is a rare swallowing disorder and is characterized by a loss of the wave-like contraction that forces food through the oesophagus and a failure of the lower oesophageal sphincter (LES) to relax during swallowing. 1 The specific cause of achalasia is unknown.
The traditional view of the renin-angiotensin system (RAS) is that of a classical endocrine system with its key components associated to the renal, hepatic and lung circulations. 2 However, it has become evident that various components of the system are expressed and act locally in many tissues. 2 A previous investigation on the wall musculature of the distal human oesophagus and LES demonstrated the existence of a local RAS, and also indicated that Angiotensin II (AngII) induces oesophageal muscle contractions via Angiotensin II type I receptor (AT1R). 3 Local AngII generation can occur through various enzymatic processes, e.g. by renin and angiotensin converting enzymes (ACE) and also through alternative pathways by cathepsin D or mast cells (MCs) proteases cathepsin G and chymase.4–6
Moreover, recent studies suggest that other angiotensin peptides like Angiotensin-(1–7) (Ang-(1–7)), acting via MAS receptor, directly antagonize many actions of AngII.7,8 Ang-(1–7) can be generated through ACE or neprilysin (NEP) activity. To what extent this local oesophageal RAS is involved in the pathophysiology of oesophageal dysmotility disorders remains to be investigated.
The aim of the present study was to elucidate the expression and distribution of the RAS looking at both the enzymatic capability responsible for AngII formation as well as Ang-(1–7) and the presence of its receptors for physiological actions in the muscular layer of the distal normal human oesophagus as well as in patients with achalasia. Mucosal levels of myeloperoxidase (MPO) served as a protein marker of neutrophil infiltration.
Material and methods
Subjects and tissue specimens
Control
Muscular specimens of the distal oesophageal body (EB) were obtained from patients (n=14, mean age 60, range 46–76, three females) undergoing radical surgery due to malignancy in the oesophagogastric junction (superficial adenocarcinoma). Specimens representing EB were obtained from 4 to 8 cm above the mucosal transition zone. Care was taken not to include tissue in close relation to the pathological process as assessed macroscopically.
Achalasia
Muscular specimens from patient with achalasia (n=10, mean age 45, range 25–66, three females) were taken during myotomy surgery. Specimens representing achalasia were obtained 5 to 7 cm above the LES.
Ethics
The study was approved by Ethical Committee of Göteborg University as well as by the Regional Ethical Review Board in Gothenburg and was performed in accordance with the Declaration of Helsinki.
Western blot analysis
Specimens were snap-frozen in liquid nitrogen and kept frozen for later western blot analysis. The frozen specimens were sonicated in a PE buffer (10 mM potassium phosphate buffer, pH 6,8 and 1 mM EDTA) containing 10 mM 3-[(3-cholamidopropyl) dimethylammonio]-1-propane sulphonate (CHAPS: Boehringer Mannheim, Mannheim, Germany) and protease inhibitor cocktail tablet Complete (Roche Diagnostics AB, Stockholm, Sweden). The homogenate was then centrifuged (10,000 g for 10 min at 4ºC) and the supernatant was analysed for protein content by the Bradford method and stored at -70ºC. 9 The samples were diluted in SDS buffer and heated at 70ºC for 10 minute and then loaded on a NuPage 10% Bis-Tris gel, and electrophoresis run using a MOPS buffer (Invitrogen AB, Lidingö, Sweden). The gel was loaded with a prestained molecular weight standard (SeeBlue, NOVEX, San Diego, CA, USA). A positive control was loaded on each gel (Table 1). After the electrophoresis the proteins were transferred to a polyvinyldifluoride transfer membrane (Hybond, 0.45μm, RPN303F; Amersham, Buckinghamshire, UK) using an iBlot (Invitrogen AB). Membranes were then incubated with specific antibodies directed at the Agt, renin, ACE, cathepsin D and G, chymase, AT1R, AT2R, NEP and MAS receptor respectively (Table 1). An alkaline phosphatase conjugated goat anti-mouse or goat anti-rabbit IgG antibody (Santa Cruz) and CDP-Star (Tropix, Bedford, MA, USA) were used as a substrate to identify immunoreactive proteins by means of chemiluminescense. Images were captured by a Chemidox XRS cooled CCD camera and analysed with Quantity One software (Bio-Rad laboratories, Hercules, CA, USA). Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) was used as the control for equal loading. For each tested sample the optical density of antibody/GAPDH represent the results. The membrane was stripped for new primary antibodies using stripping buffer (Re-Blot Plus Mild Solution (10X), Millipore, Temecula, CA, USA).
Antibodies and controls used in western blot analyses and immunohistochemistry.

Agt: angiotensinogen; ACE: angiotensin converting enzyme; MPO: myeloperoxidase; AT1R: angiotensin II type 1 receptor; AT2R: angiotensin II type 2 receptor; GAPDH: glyceraldehyde-3-phosphate dehydrogenase.
Immunohistochemistry
Muscular specimens from EB and patients with achalasia were fixed in 4% formaldehyde, dehydrated and embedded in paraffin. Sections for immunohistochemistry were deparaffinized and then boiled in 50 mM citrate buffer (pH 6.0) for 15 minutes to make the antigens unfold. The CSA II biotin free tyramide signal amplification staining system kit (Dako Sweden AB, Stockholm, Sweden) was used for the immunohistochemistry protocol. After inhibition of endogenous peroxidase activity, the slides were pre-incubated with protein block and then incubated with primary antibodies against Agt, renin, ACE, cathepsin D and G, chymase, MPO, AT1R, AT2R, NEP and MAS receptor (see Table 1) in dilutions of 1:500 to 1:1000 for 15 minutes. Control sections were incubated with wash buffer instead of the primary antibody. After being washed, the slides were covered with anti-rabbit immunoglobulins-HRP and incubated for 15 minutes. This was followed by sequential incubations with amplification reagent (fluorescyl-tyramide hydrogen peroxide) in darkness and anti-fluorescein-HRP, 15 minutes each. The staining procedure was completed by incubation with 3,3’ diaminobenzidine. Muscular sections were also taken for Toluidine blue staining reactions for visibility of MCs and Bielschowsky’s silver staining for neurofibrils, dendrites and axons.
Reverse transcriptase polymerase chain reaction (RT-PCR)
Biopsies were snap frozen in RNA STAT-60 (Nordic Bio Site AB, Stockholm, Sweden) in liquid nitrogen. Frozen tissue was homogenised and total RNA was extracted according to the methods suggested by the manufacturer, following phenol-chloroform extraction and ethanol precipitation. Reverse transcription from 2.5 µg of total RNA was carried out using the SUPERSCRIPT™ First-Strand Synthesis System (Invitrogen, Lidingö, Sweden) with Oligo (dT) Primers (Life Technologies, Täby, Sweden). The resulting cDNA was stored at -20ºC until use. LightCycler Q-PCR (Roche Diagnostics AB) was performed using the FastStart DNA Master SYBR Green I (Roche Diagnostics AB). RT-PCR was performed containing 2 μL of each RT sample using the hot-start technique. MgCl2 concentration was optimised to 4 mM to obtain the highest signal intensity and lowest background. The copy numbers of the MC chymase gene in each sample were interpolated with a standard curve from five to seven dilutions of a positive sample. For each tested sample the copy numbers were divided by the total RNA concentration as an internal standard, measured with a GENios spectrophotometer (TECAN, Salzburg, Austria). Copy numbers of target genes are then expressed as copy number per µg total RNA. The quantification was performed by the software supplied by Roche Diagnostics (Mannheim, Germany). The human chymase mRNA fragment contains 323 bp and was amplified using the following primers; Forward: 5’-GAA GGA GAA AGG CAG CCT GAC CCT G-3’ and Reverse: 5’-CAT CCG ACC GTC CAT AGG ATA CGA TG-3’. 10
Measurements of AngII with enzyme immunoassay
Muscular biopsies were extracted for protein content using the above described protocol for western blotting and analysed for protein content by the Bradford method and stored at -70°C until analysis. Samples were diluted in EIA-buffer (SPI-Bio/Bertin Pharma) and concentrations of AngII (pg/mg tissue) were measured by a commercial enzyme immunoassay (ANG II EIA kit, SPI-Bio/Bertin Pharma, Montigny-le-Bretonneux, France) according to the manufacturer’s instructions. The limit of detectability was 0.09 pg/mg tissue protein and CV was 5%.
Statistical analysis
Data are presented as means ± SEM. Significant differences for multiple independent groups of observation were identified using the Kruskal-Wallis test and contrasted using the Mann-Whitney U test. Linear relationships were tested with Spearman’s correlation. Statistical significance was set at p≤0.05 and the analyses were carried out using the SPSS software, Version 19.0 (SPSS Inc., Chicago, IL, USA).
Results
Expression of angiotensin formation enzymes
Western blotting analysis exhibited the presence of the prohormone Agt, the enzymes renin, ACE, cathepsin G and D, chymase, and NEP in the muscular sample. The distribution of renin (p=0.009) and cathepsin D (p=0.002) were significantly lower (Figure 1(b) and (d)), whereas the expression of cathepsin G (p<0.001, Figure 2(a)) and NEP (p=0.001, Figure 1(e)) were significantly increased in the muscular tissue from patients with achalasia compared to controls. The expression of Agt, ACE (Figure 1(a) and (c)) and chymase (Figure 2(b)) are at a similar level independent of group affiliation.
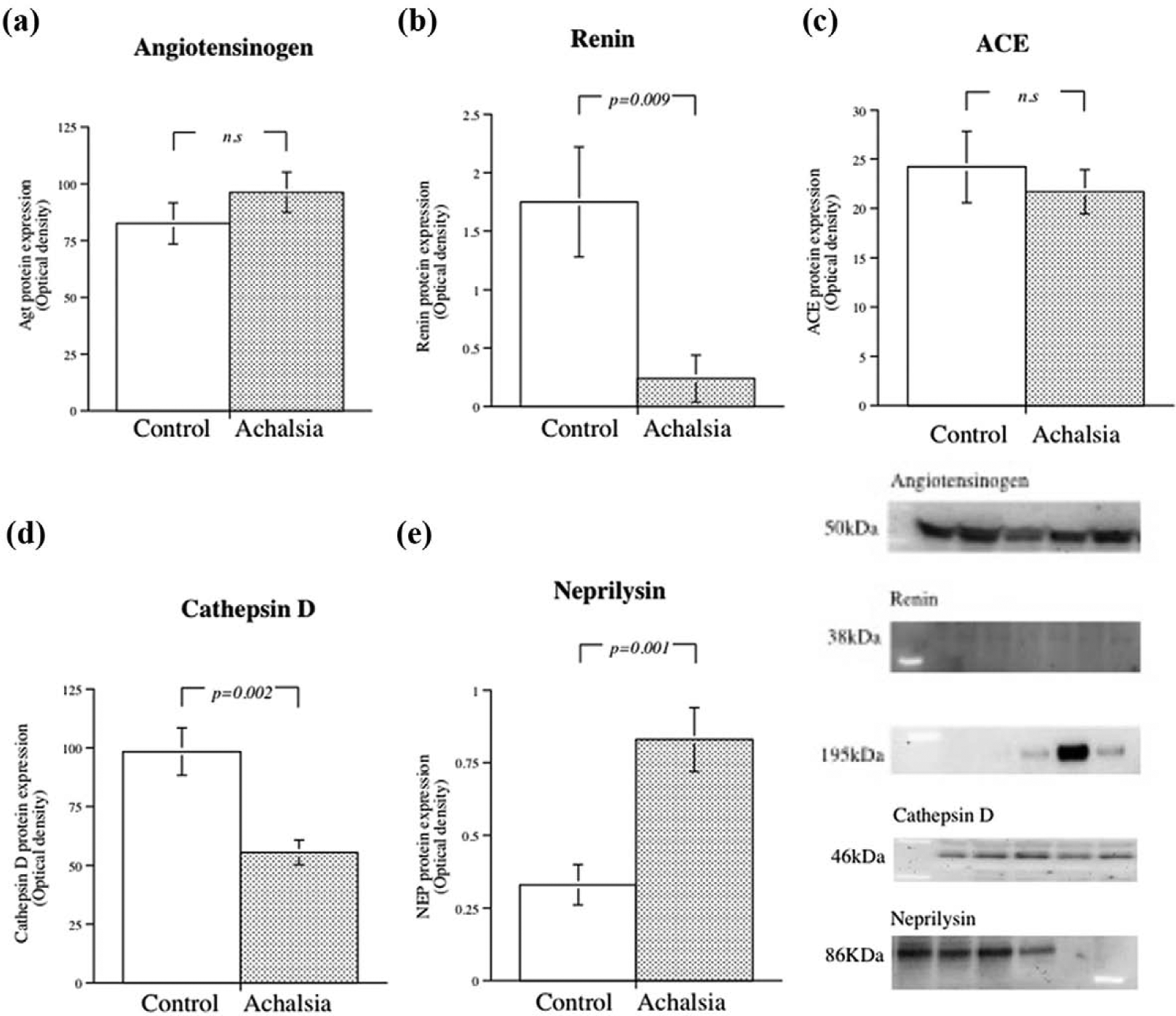
Expression of renin-angiotensin system proteins in the human oesophagus muscular tissue from controls and from patients with achalasia.

Expression of mast cells (MCs) proteases cathepsin G and chymase in the human oesophagus muscular tissue from controls and from patients with achalasia.
Real-time PCR analysis exhibited RNA indicating transcript of human chymase, showed significantly higher expression of chymase in achalasia muscular cells (p=0.039) (data not shown). The expression of MPO protein was markedly higher in patients with achalasia (p=0.016) than in controls (Figure 2(c)). A positive correlation was also seen between chymase and cathepsin G protein expression and the degree of neutrophil infiltration reflected by MPO (chymase; r=893, p<0.01: cathepsin G; r=672, p<0.01, Figure 2(d) and (e)).
Immunohistochemistry revealed expression of the enzyme renin, ACE and NEP in the blood vessel wall; whereas immunoreactivity for cathepsin D was found in the smooth muscle cells lysosomes. Staining for Agt was principally located to the blood vessel wall but was indistinct and somewhat obscure in nature (data not shown). Histological examination with Toluidine blue staining reactions showed presence of MCs in close contact with the muscular cells as well as nearby blood vessels in the oesophageal muscular preparations (Figure 3(a) and (b)). Immunostaining for cathepsin G, MPO and chymase demonstrated co-localization with MCs in the tissue (Figure 3(c) to (f)). Bielschowsky’s silver staining showed neurofibrils, dendrites and axons around each smooth muscle in the tissue preparation (Figure 4).
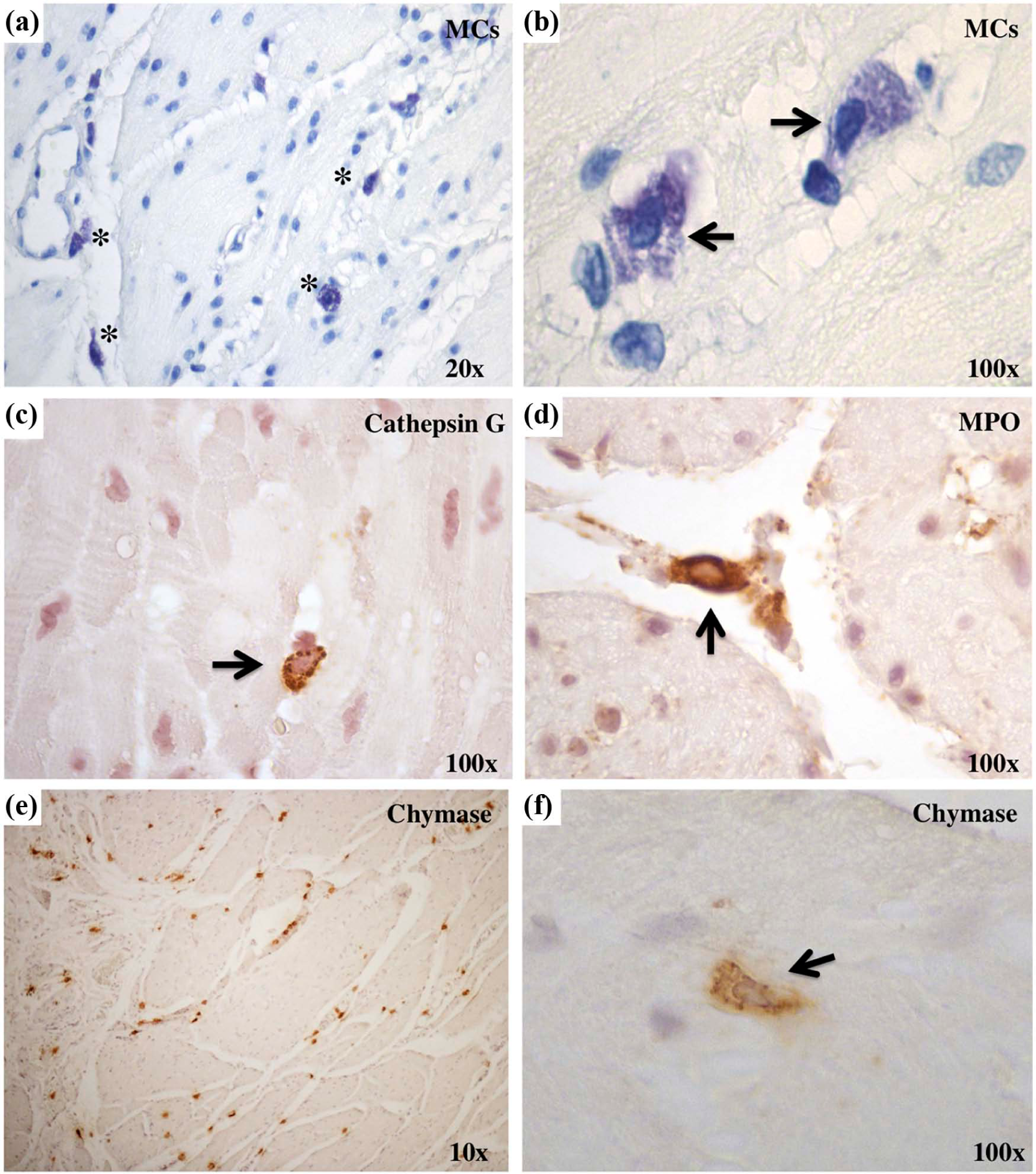
Histological examination of mast cells (MCs) in patients with achalasia.

(a) Bielschowsky’s silver staining showed neurofibrils, dendrites and axons in human oesophageal muscular tissue (magnification 100X). (b) Hematoxylin-eosin staining in human eosophageal muscular tissue (magnification 10X).
Angiotensin peptides formation and their receptors
Tissue contents of AngII tended to be higher in the muscle cells taken from EB (3.0±1.04 pg/mg) compared to achalasia (0.85±0.38 pg/mg tissue protein, p=0.083), but that difference did not attain statistical significance (data not shown). The AT1R protein expression was located direct in the smooth muscle cells (Figure 5(a)) and was significantly lower in achalasia muscle cells specimens (p=0.022, Figure 6(a)), whereas almost no detectible levels of AT2R were found in the samples (data not shown). Distinct staining for MAS receptor was found in the muscle cells (Figure 5(b)) and was markedly higher in patients with achalasia (p=0.028, Figure 5(c)).

Immunohistochemistry of the angiotensin II and angiotensin-(1–7) receptors in human oesophagus muscular tissue.
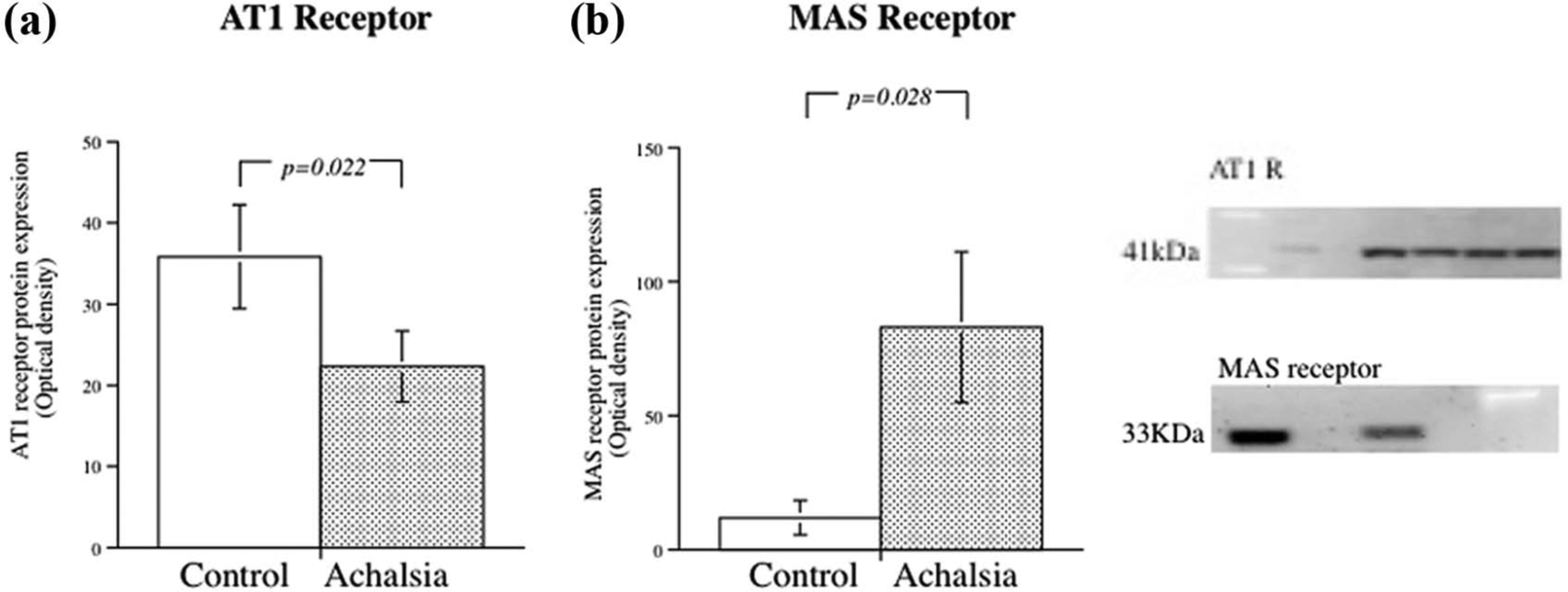
Expression of angiotensin II type 1 receptor (AT1R) and the angiotensin-(1–7) receptor MAS in human oesophageal muscular preparations.
Discussion
AngII has been shown to exert potent contractile effects via AT1R in different gastrointestinal smooth muscles11–14 and also in human EB and LES. 3 The present study investigated the expression of the RAS in muscular tissue taken from patients undergoing myotomy surgery due to the oesophageal dysmotility disease achalasia in comparison to EB taken from patients undergoing radical surgery due to malignancy in the mucosa. The key results from the present study are the decreased expression of AT1R, the main receptor for AngII contraction, together with increased MAS receptor, the main receptor for Ang-(1–7) in patients with achalasia. In addition, muscular specimens demonstrated increased expression of MCs chymase and cathepsin G that correlated with neutrophil infiltration reflected by MPO, suggesting that RAS production are in part regulated by the inflammatory states in patients with achalasia.
The concept of tissue RAS is now well established. At the same time it has emerged that AngII is not the only biologically active peptide generated by the RAS. The RAS is therefore a much more complex system than previously believed. The effects of the angiotensin peptides are related to its binding to specific receptors. The AT1R mediates all classical actions of AngII such as contributing to the maintenance of arterial blood pressure, electrolyte and water balance, thirst, renal function and structural remodelling of cardiovascular tissue.2,15 The presently reported expression of AT1R showed it located directly at the muscular cells, suggesting direct action by AngII on smooth muscle contraction. However, the achalasia muscular tissue under study showed decreased AT1R expression, which may indicate poor response to AngII on the muscle contraction.
The AT2R counterbalancing most effects that AngII exert through the AT1R, such as AT1R mediated contractions, could be counteracted by AT2R mediated relaxations. 16 The AT2R expressions that have been reported also vary according to tissue conditions, for example hypoxia and inflammation promote AT2R expression. 17 However, the achalasia muscular tissue lacked significant expression of the AT2R that was reported earlier for EB and LES. 3 The AT2R mediated relaxation is excluded as an important factor in its failure of the LES to relax completely as observed in achalasia.
The Ang-(1–7) produces vasodilation, diuresis and antiproliferative reactions through the specific MAS receptor. 7 Studies on rat thoracic aorta ring has shown that Ang-(1–7) can inhibit the vasoconstriction induced by AngII indicating that Ang-(1–7) behaves like a true counteracting factor for AngII and its AT1R mediated effects. 7 In the absence of AT2R, the increased expression of MAS receptor in the muscle cells seen in patients with achalasia may therefore act as a new interesting counterbalancing factor for AngII and the AT1R in, for example, muscle relaxation. RAS can be envisioned as a dual function system in which the action are primarily driven by the balance of ACE;AngII;AT1R or by NEP;Ang-(1–7);MAS receptor, respectively. Commonly drugs like ACE inhibitors or AT1R blockers may shift the balance towards the NEP; Ang-(1–7); MAS receptor axis. Indeed, it has been shown that ACE inhibitors and AT1R blockers may prevent stress-induced gastric injury, an action mediated by Ang-(1–7). 18 Future studies are needed to further elucidate the significance of Ang-(1–7) and MAS receptors involving oesophageal motility.
The hallmark functional observation in achalasia is loss of oesophageal peristalsis and a failure of the LES to relax completely. Otherwise the LES is tonically contracted and relaxation occurs when the mechanism that leads to tonic LES contractions are interrupted. This relaxation is predominantly neuronally initiated, coordinated by both spinal and vagal afferent. Studies on the achalasia smooth muscle has shown loss of myenteric nerve fibres in the LES and in the oesophageal wall, as well as reduced number of the interstitial cells of Cajal (ICC).19 –22 ICC may serve as pace makers, as generators of smooth muscle hyper polarisation, as mechanical sensors and as mediators of neurotransmission.20,21 However, routine identification of ICCs using C-kit make it impossible to clearly sort out the difference between ICCs and MCs.
Our histological staining showed MCs in close contact with the muscle cells as well as with blood vessels. Previous studies have found macrophages and MCs in contact with the ICC suggesting functional interaction.21,23 Macrophages and MCs are considered to play important roles in the innate immune defence by producing pro-inflammatory mediators during classical activation, which may in itself result in damage to the tissue, or they may take part in alternative activation, such as with anti-inflammatory mediators, tissue remodelling and homeostasis. 24 MCs proteases chymase has been previously shown to catalyse the formation of AngII which when released might act as a paracrine mediator in the enteric nervous system such as in guinea pig small intestine. 25 Thus, as this dysmotility disorders is predominantly neuronally initiated it would be speculated that AngII might have a main function as a neurotransmitter. Presently Bielschowsky’s silver staining showed neurofibrils, dendrites and axons around each smooth muscle cell and locally produced AngII (through traditionally or alternative pathways) could act as a neurotransmitter in the enteric nervous system (ENS).
The localization of Agt together with the enzymes renin and ACE, in particular the vascular endothelial cells, strongly indicates that endogenous AngII is normally formed in the oesophageal muscular tissue. Present AngII measurements using commercial enzyme immunoassay showed that tissue contents of AngII tended to be lower in achalasia but that difference did not attain statistical significance. This may perhaps be in consequence to increased MCs proteases chymase and cathepsin G in achalasia that are discovered as an AngII-producing enzyme in human neutrophils.5,6 Wang et al. has found that elevated levels of AngII in mucosal biopsies from patients with Crohn’s colitis might be associated with the inflammatory states, which include MC hyperplasia. 25 The present western blot analysis of neutrophil infiltration reflected by MPO was marked increased in patients with achalasia and correlates significantly to chymase and cathepsin G expression. Also, immunohistochemistry demonstrated that chymase, cathepsin G as well as MPO co-localized with MCs in the tissue.
Cathepsin D is not usually thought to participate as an enzymatic alternative to renin, but Naseem et al. has shown that cathepsin D generate AngI in presence of Agt after myocardial infarction. 4 Cathepsin D is a widely distributed aspartyl protease that enzymically breaks down a variety of different protein substrates within the lysosomal compartment of most cells. 26 Present immunostaining showed cathepsin D in smooth muscle cells lysosomes and western blotting showed decreased expression in the groups of patients with achalasia. However, further study is needed to understand the role of cathepsin D protease and its capacity to break down different proteins in the tissue.
Conclusion
The present results showed the existence of a local RAS system in the human EB as well as in patients with achalasia. The enzymes responsible for AngII production are however differently in patient with achalasia, there alternative pathways through mast cells proteases are in favourite, as well as the enzyme responsible for Ang-(1-7) production. In addition, there has been a shift in receptor physiology from AT1R to MAS receptor in the smooth muscle taken from patients with achalasia. It thus remains to be investigated if these changes in RAS might play a significant role in the physiological motor control in patients with achalasia.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.