
Abstract
Although angiotensin (Ang) II blockade has become a standard antifibrotic therapy in kidney disease, the therapeutic efficacy of Ang II blockade is yet to be optimized. Considering the prognostic impact of proteinuria reduction, we hypothesized that titration of Ang II blockade for optimal anti-proteinuric effect would improve renoprotection. One day after induction of Thy 1.1 glomeruonephritis, rats were treated with increasing doses of the Ang II receptor blocker valsartan in drinking water. Six days after disease induction, the therapeutic effect on proteinuria, podocyte injury and glomerular fibrosis was evaluated. Increasing doses of valsartan resulted in increasing reduction of proteinuria. The maximally effective dose of valsartan was determined to be 1000 mg/l, which reduced proteinuria by 80% and maximally reduced glomerular matrix expansion, fibronectin, collagen I and collagen III staining and glomerular mRNAs for TGFß1, PAI-1, FN and collagen I. Notably, valsartan given at this dose prevented podocyte dysfunction by preserving expression of podocin and nephrin and the counter-regulating molecule B7-1 that is involved in podocyte injury. These results support the hypothesis that higher doses of valsartan are required to optimize proteinuria reduction and glomerulosclerosis amelioration. Further, the optimal dose of valsartan also provides an additional therapeutic effect by preventing podocyte dysfunction.
Introduction
Activation of the renin-angiotensin system (RAS) and generation of angiotensin II (Ang II) have long been known to play a crucial role in the pathogenesis of fibrotic kidney disease beyond this system’s hemodynamic actions. 1 Pharmacological blockade of Ang II’s generation and action therefore has become a standard and essential therapy in the management of a variety of kidney fibrotic diseases. However, most studies to date show kidney disease progression in many patients despite treatment with angiotensin-converting-enzyme inhibitors (ACEIs) or angiotensin II-receptor blockers (ARBs). It has been assumed that the limited renoprotection offered by ACEIs or ARBs is due to uncompleted suppression of the RAS. One of the strategies that could improve the efficacy of RAS blockade is titration of ACEI or ARB for optimal therapeutic effect. Emerging evidence has suggested that this approach does confer a further decrement in urinary protein excretion, particularly with higher doses of ARB.2–11 Furthermore, higher doses of ARB are generally well tolerated. However, large, long-term trials with hard outcomes are currently lacking for this potentially promising form of therapy. We have shown that increasing the dose of enalapril or losartan above the respective maximum anti-hypertensive dose produced additional anti-fibrotic effects in nephritic glomeruli 12 when reduction of glomerular transforming growth factor beta (TGFß) overproduction was chosen as the therapeutic targeting. However, glomerular or urinary TGFß reduction is not easily monitored for titration of ACEI or ARB for their optimal therapeutic effect in patients. Increased urinary excretion of albumin or total protein has become firmly established as an important progression promoter of kidney disease, and reduction of proteinuria has been associated with decreased risk of major clinical events. To further investigate whether targeting the optimal dose of Ang II blockade that further reduces proteinuria may yield better kidney outcomes and to investigate the potential protective actions, a systematic prospective study with a broader range of doses of an ARB, valsartan (50 to 1500 mg/l in drinking water), was carried out in the rat Thy 1.1 glomerulonephritis model.
Compared to other models of kidney disease, the model of mesangioproliferative glomerulonephritis is one in which administration of an antibody to the Thy 1.1 epitope on glomerular mesangial cells leads to immune-mediated, complement-dependent mesangial cell lysis. It is characterized by a very rapid and marked proteinuria accompanied by glomerular accumulation of extracellular matrix (ECM). 13 This model has great relevance because it contains all of the elements of human fibrotic diseases, whether acute or chronic. In addition, this model is reproducible and can be used to very sensitively discriminate between therapeutic regimens.12,14,15 Therefore, this model was chosen for titration of valsartan for its optimal therapeutic effect in vivo.
We demonstrate that increasing doses of valsartan resulted in increasing reduction of proteinuria and renal fibrosis. Optimal anti-proteinuric doses of valsartan are also required to optimize renoprotection. Further, more complete Ang II inhibition with the maximally effective dose of valsartan prevents podocyte dysfunction effectively.
Materials and methods
Reagents
Valsartan was kindly provided by Novartis Pharmaceuticals Corporation (Basel, Switzerland). Unless specified, all other reagents were purchased from Sigma Chemical Co. (St. Louis, MO, USA).
Animals
The studies were performed in male Sprague-Dawley (SD) rats (180–200 g) obtained from the SASCO colony of Charles River Laboratories (Wilmington, MA, USA). Animal housing and care were in accordance with the United States National Institutes of Health (NIH) Guide for the Care and Use of Laboratory Animals. The animal studies were approved by the Institutional Animal Care Committee of the University of Utah. Glomerulonephritis was induced by tail-vein injection of 1.75 mg/kg of the monoclonal anti-Thy 1.1 antibody, OX-7. OX-7 binds to a Thy 1.1-like epitope on the surface of mesangial cells, causing immune-mediated, complement-dependent cell lysis followed by exuberant matrix synthesis and deposition. 13 Normal control animals were injected with the same volume of phosphate-buffered saline (PBS).
Experiment design
A dose-response experiment was carried out to determine the effective dose range of valsartan in anti-Thy 1.1 glomerulonephritis. Eight groups of five rats (n = 5, each group) were assigned and treated as normal-control, disease-control and nephritic rats treated with 50, 100, 250, 500, 1000 and 1500 mg of valsartan per liter of drinking water, as outlined in Table 1. Since the dose of 1000 mg of valsartan per liter of drinking water (approximately equivalent to 160 mg/kg per day) has been tested in animal study and was not confirmed as the valsartan threshold, 5 an even higher dosage at 1500 mg/l was included in this study. Treatment was started 24 hours after antibody injection when mesangial cell lysis is complete and the fibrotic response starts. At the same time, based on an average water intake of 40 ml per day, 60% of daily dose was administered by gavage as described previously. 12 Thereafter, the water intake was monitored daily and the amount of drug in the drinking water was adjusted daily to the amount each group of animals would receive by drinking 40 ml water.
Experiment design.

PBS: phosphate-buffered saline; Val: valsartan.
Urine samples were collected from rats housed in metabolic cages for 24 hours from day 5 to day 6 after OX-7 injection. The urinary protein excretion was measured by the Bradford method (Bio-Rad Laboratories Inc., Hercules, CA, USA). Urine albumin and urine creatinine were measured using the DC2000+ microalbumin/creatinine reagent kit (Bayer Healthcare, Elkhart, IN, USA). Urine creatinine was also measured for the calculation of creatinine clearance ratio using the QuantiChromTM creatinine assay kit (BioAssay System, Hayward, CA, USA).
On day 6, animals were anesthetized, 5 to 10 ml blood was drawn from the lower abdominal aorta for detection of plasma blood urea nitrogen (BUN) and creatinine levels (QuantiChromTM urea and creatinine assay kits) and kidneys were perfused with 30 ml ice-cold PBS. The creatinine clearance was calculated as the following formula, (Urine creatinine levels/plasma creatinine levels) × 24-h urine volume (ml)/(24 h × 60 min). For histological examination, cortical tissue was snap frozen or fixed in 10% neutral-buffered formalin for periodic acid Schiff (PAS) staining. Glomeruli were isolated by graded sieving as described previously 15 for RNA and protein isolation.
The dose results indicated that a valsartan dose of 1000 mg/l (40 mg per day, approximately equivalent to 160 mg/kg per day) maximally reduced proteinuria, albuminuria and glomerular fibrotic markers, so the effect of valsartan at this dose on podocyte injury was examined in nephritic rats, compared with normal and disease controls.
Histological analyses
All microscopic examinations were performed in a blinded fashion. Three-micrometer sections of paraffin-embedded tissues were stained with PAS. The images (×400 magnification) of 20 random glomeruli per slide were captured using a Nikon D50 digital camera (Inkley's-Ritz Camera, www.ritzcamera.com, Salt Lake City, UT; Nikon Capture 4 Ver. 4.3, Nikon Inc., Melville, NY, USA), and the area of PAS-positive mesangium matrix indicating mesangial expansion in each glomerulus was quantified using a computer-assisted color image analysis system (Image J 1.38 for Windows; National Institutes of Health http://rsb.info.nigh.gov). The PAS-positive material area in the mesangium was normalized by that of the total glomerular tuft where the percentage of mesangial matrix occupying each glomerulus was rated on a 0–4 scale (0=0, 1=25, 2=50, 3=75, or 4=100%), as described previously. 16
Immunofluorescent (IF) staining for matrix proteins was performed on frozen sections and evaluated in 20 glomeruli from each rat. Monoclonal mouse anti-cellular fibronectin (FN) extradomain A+ (EDA)+ (Harlan Sera-Lab Belton, England), goat anti-human type I collagen and goat anti-human type III collagen (Southern Biotechnology Associates, Birmingham, AL, USA) were used as the primary antibodies for detection of ECM components. Fluorescein isothiocyanate (FITC)-conjugated rat F (ab’) 2 anti-mouse immunoglobulin G (IgG) (H+l) (Jackson ImmunoResearch Laboratories, West Grove, PA, USA) and FITC-conjugated rabbit anti-goat IgG (DAKO North America Inc., Carpentaria, CA, USA) were used as the secondary antibodies. Intraglomerular deposition of these ECM components was quantified as described previously. 16 A whole kidney average positive staining for matrix protein was obtained by averaging scores from all glomeruli.
IF staining for antibodies against podocin and Wilms tumor protein 1 (WT-1) was performed on frozen sections. The polyclonal goat anti-podocin IgG (G-20) and polyclonal rabbit anti-WT-1 IgG (C-19) (Santa Cruz Biotechnology Inc., Santa Cruz, CA, USA) were used as the primary antibodies. The Rhodamin (tetramethylrhodamine-5-(and 6)-isothiocyanate (TRITC))-conjugated donkey anti-goat IgG (H+L) and TRITC-conjugated goat anti-rabbit IgG (Jackson ImmunoResearch, Laboratories Inc.) were used as the secondary antibodies. Intraglomerular positive staining for podocin was also quantified in a blinded fashion by a computer-assisted method as described above. The number of WT-1-positive podocytes per glomerulus was counted in 20 glomeruli selected randomly per section.
Western blot analysis
Glomeruli from individual rats were isolated and resuspended at 2 × 104 glomeruli/ml in lysis buffer (Cell Signaling Technology Inc., Beverly, MA, USA) with 1% NP40, 1mM phenylmethylsulfonyl fluoride (PMSF) and one tablet/5 ml protease inhibitor mix (Complete, Mini; Roche Diagnostics Corp., Indianapolis, IN, USA). Glomeruli were homogenized twice on ice by sonication. Each 15-second sonication was followed by a 15-second cool down. After two centrifugations at 10,000 × g for 10 minutes at 4ºC, the supernatant was stored at −70ºC until analysis. The same amount of glomerular protein (determined by bicinchoninic acid (BCA) protein assay, Pierce, Rockford, IL, USA) from five rats of each group was pooled. Twenty µg of protein samples were separated by 4–12% Bis-glycine gel electrophoresis (Invitrogen, Carlsbad, CA, USA) and transferred to a 0.45-µm immobilon-P transfer membrane (Millipore Corp., Bedford, MA, USA). The glomerular protein levels of nephrin and B7-1 were developed using polyclonal rabbit anti-nephrin IgG and monoclonal rabbit anti-B7-1/CD80 IgG (OriGene Technologies Inc., Rockville, MD, USA). The immunostaining band was visualized and quantified using a Bio-Rad GS-700 imaging densitometer as described previously. 16 Changes in nephrin and B7-1 protein expression were determined by normalizing against the densitometric intensity of its ß-actin (mouse monoclonal anti-ß-actin IgG) for each sample. For comparison, this ratio was set at unity for normal-control samples and other lanes on the same gel were expressed as fold-change over this value. All blots were run at least three times in duplicated wells.
RNA preparation and real-time reverse transcriptase-polymerase chain reaction (RT-PCR)
Total RNA was extracted immediately from isolated glomeruli using TrizolTM Reagent (GibcoBRL, Gaithersburg, MD, USA) according to the manufacturer’s instructions. RNA from five rats of each group was pooled. Two micrograms of total RNA were reverse-transcribed using the superscript III first-stand synthesis system for RT-PCR kit (Invitrogen). Real-time RT-PCR was performed using a SYBR Green dye I (Applied Biosystems, Foster City, CA, USA) with the ABI 7900 Sequence Detection System (PE Applied Biosystems) as described previously. 16 Samples were run as triplicates in separate tubes to permit quantification of the target gene normalized to glyceraldehyde-3-phosphate dehydrogenase (GAPDH). Sequences of primers used are listed in Table 2. The specificity of the PCR products was confirmed on a 1.5% agarose gel by showing a specific single band with the expected size.
Primers used for real-time reverse transcriptase-polymerase chain reaction.
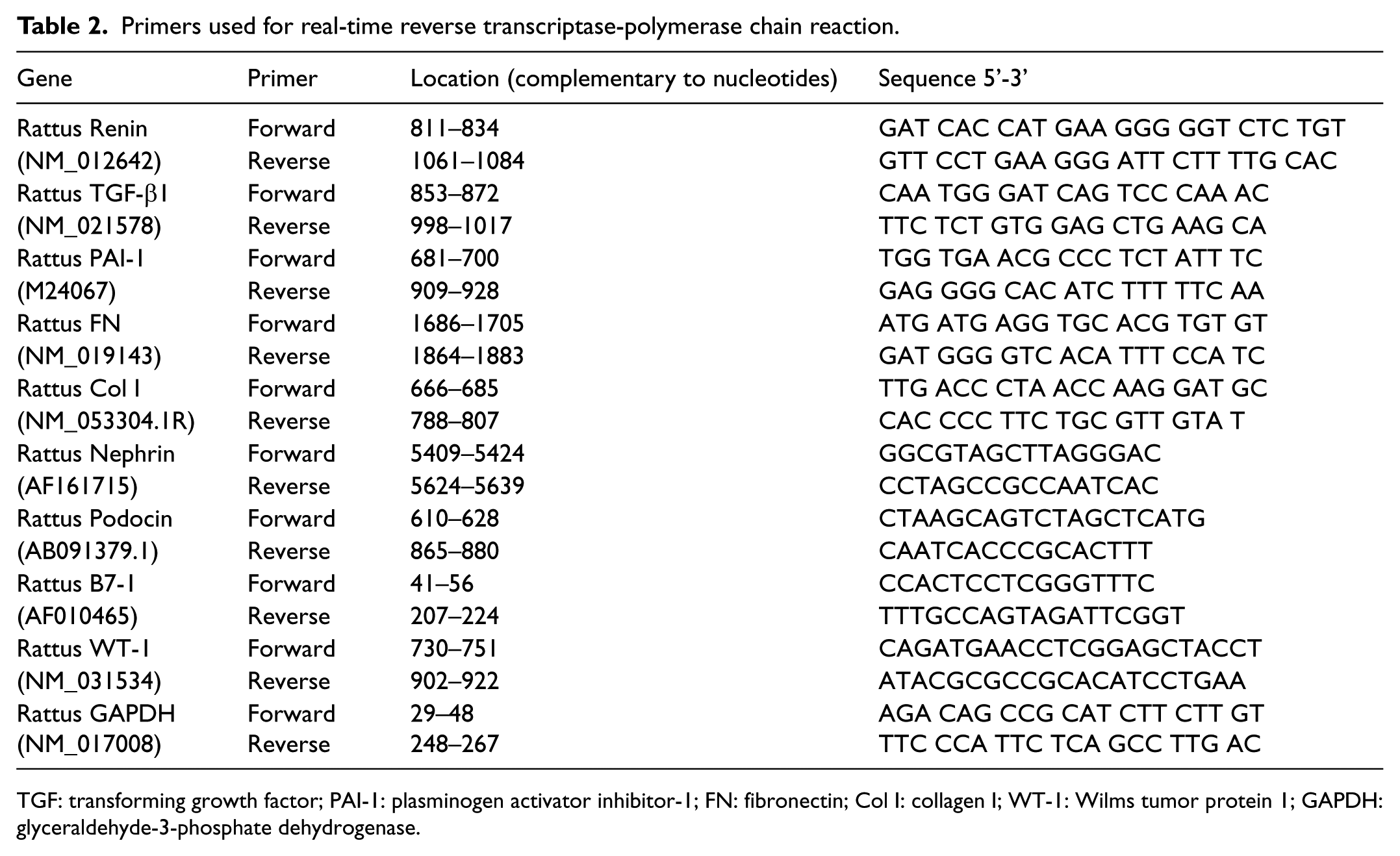
TGF: transforming growth factor; PAI-1: plasminogen activator inhibitor-1; FN: fibronectin; Col I: collagen I; WT-1: Wilms tumor protein 1; GAPDH: glyceraldehyde-3-phosphate dehydrogenase.
Statistical analyses
Data are expressed as mean ± standard deviation (SD). Statistical analysis of difference among groups was performed by Tukey multiple comparisons test; p < 0.05 was considered statistically significant. The disease-induced increase in a variable was defined as the mean value for the disease control group minus the mean value of the normal-control group. The percentage of reduction in disease severity in a treated group was calculated as before. 16
Results
Therapeutic efficacy of valsartan
Effects of valsartan on renal function and urinary protein excretion in anti-Thy 1.1 nephritis
The induction of disease by the administration of OX-7 induced significant increases in plasma BUN and creatinine levels (Table 3) and a significant decrease in creatinine clearance rate (Ccr) levels, indicating damaged renal function. Treatment with valsartan did not affect plasma BUN and creatinine levels. The Ccr levels after valsartan treatment in the drinking water were not decreased but increased, although the increases did not reach statistical significance. Twenty-four-hour urinary protein excretions were measured from day 5 to day 6 (Figure 1). The induction of disease produced a marked increase in total proteinuria at day 6 (Figure 1A). Treatment with valsartan significantly reduced disease-induced proteinuria in a dose-dependent manner. The addition of 50 mg/l valsartan to the drinking water reduced proteinuria by 21%. A maximal decrease of approximately 80% was seen at 1000 mg/l valsartan (40 mg per day, approximately equivalent to 160 mg/kg per day). However, the inhibitory effect was paradoxically attenuated, with a further increase in the valsartan dose to 1500 mg/l; the decrease in urinary protein excretion was 55%, which was close to the effect of 500 mg/l valsartan (~48% reduction, p > 0.05). In order to assess not only quantitative but also qualitative effects of valsartan on proteinuria, we measured urinary albumin level and calculated the ratio of urinary albumin level to urinary creatinine level at various doses of valsartan. As shown in Figure 1B, valsartan therapy similarly reduced disease-induced urinary albumin excretion in a dose-dependent manner, with the maximal reduction observed at 1000 mg/l. Similar to its effect on proteinuria, a further increase in valsartan dose paradoxically attenuated the inhibitory effect to that of 500 mg/l valsartan on albuminuria (Figure 1B). These results were consistent with those determined by the ratio of urinary albumin level and urinary creatinine level, as shown in Figure 1C.
Renal function of the experimental groups of rats.

NC: normal control: DC: disease control; Val: valsartan; BUN: blood urea nitrogen. * p<0.05 vs NC; # p<0.05 vs DC.

Effect of increasing doses of valsartan in drinking water on (A) 24-hour (h) urinary protein excretion, (B) 24-h urinary albumin excretion and (C) the ratio of urinary albumin level and creatinine level. Treatment was started one day after injection of OX-7. The 24-h urine samples were harvested from day 5 to day 6 after disease induction. * p < 0.05 vs normal control (NC); # p < 0.05 vs disease control (DC).
PAS staining
Representative glomeruli stained with PAS are shown in Figure 2A. The glomeruli from the disease-control rats showed marked accumulation of ECM expressed as PAS-positive material at day 6 (Figure 2A-DC) compared to normal glomeruli (Figure 2A-NC). Increasing doses of valsartan (50–1500 mg/l) produced progressive decreases in glomerular matrix accumulation (Figure 2A-V50 to V1500). Figure 2B shows a graphical representation of the matrix score for each group. A significant (p < 0.05) decrease of 50% in PAS staining was seen in rats treated with either 500 mg/l or 1000 mg/l valsartan, compared to disease-control animals (Figure 2A). However, this inhibitory effect was attenuated by 9% when an even higher dose of valsartan (1500 mg/l) was given (p < 0.05).

Effect of increasing doses of valsartan in drinking water on glomerular matrix accumulation. (A) Representative photomicrographs of glomeruli from normal-control rats (NC), disease-control rats (DC), and various doses of valsartan-treated nephritic rats at day 6 (Val 50 to Val 1500, 50 to 1500 mg/l valsartan in drinking water). Original magnification × 400. (B) Graphic representation of glomerular extracellular matrix score. * p < 0.05 vs. NC; # p < 0.05 vs. DC.
IF staining
In order to determine the contribution of specific matrix proteins to the PAS-positive material, IF staining for various specific glomerular proteins was performed. The results of the semi-quantitative analysis of those stainings are shown in Figure 3. Substantial increases in staining for matrix proteins were seen in disease-control animals (2.1-fold for FN EDA+, 5.3-fold for collagen I and 1.87-fold for collagen III), compared to normal-control animals (Figure 3A–C). Treatment with valsartan caused a dose-dependent reduction of these proteins. The maximally effective trend for reduction of collagen I and collagen III accumulation in glomeruli was seen at the valsartan dose of 1000 mg/l, while the maximally effective trend for reduction of FN EDA+ accumulation was achieved at the dose of 500 mg/l. Although the highest dose of valsartan showed fewer effects on those matrix protein accumulations in glomeruli (26% for collagen I, 45% for collagen III and 52% for FN EDA+) when compared to those of 1000 mg/l valsartan (33% for collagen I, 53% for collagen III and 55% for FN EDA+), the differences between the effects of these two dosages did not reach significance (p > 0.05).

Effect of increasing doses of valsartan in drinking water on glomerular immunofluorescent staining score for extracellular matrix (ECM) proteins in anti-Thy 1.1 nephritis at day 6. Glomerular staining for fibronectin (FN)-extradomain A+ (EDA+) (A), collagen I (Col I) (B) and collagen III (Col III) (C) are lower in the valsartan-treated nephritic groups in a dose-dependent manner. * p < 0.05 compared with normal control (NC); # p < 0.05 compared with disease control (DC).
Effects of valsartan on glomerular mRNA levels of TGFß1, plasminogen activator inhibitor-1 (PAI-1), FN and type I collagen in anti-Thy 1.1 nephritis
As shown in Figure 4A–D, glomerular mRNA analysis revealed a three-fold increase in TGF-ß1, a 17-fold increase in PAI-1, a 12-fold increase in FN EDA+ and a 43-fold increase in type I collagen mRNA expression in disease-control rats compared to normal-control rats. Valsartan administration significantly reduced the gene expression of TGFß1, PAI-1, FN EDA+ and type I collagen in a dose-dependent manner. Consistently, the maximal effect for inhibition of TGFß1, PAI-1, FN EDA+ and type I collagen expression was achieved at the valsartan dose of 1000 mg/l. Valsartan maximally reduced TGFß1 mRNA expression by 76%, PAI-1 by 65%, FN EDA+ by 76% and collagen I by 70%, respectively (Figure 4A–D). There was no further reduction of TGFß1 mRNA expression when the highest valsartan dose was given (p > 0.05). The effect of the highest valsartan dose of 1500 mg/l on the gene expression of PAI-1 and these matrix proteins appeared to be attenuated to that of the 500 mg/l valsartan dose.

Effect of increasing doses of valsartan in drinking water on glomerular mRNA expression in anti-Thy 1.1 nephritis at day 6. Glomerular mRNA levels of transforming growth factor beta 1 (TGFß1) (A), plasminogen activator inhibitor-1 (PAI-1) (B), fibronectin (FN) extradomain A+ (EDA+) (C) and collagen I (Col I) (D), determined by real-time reverse transcriptase-polymerase chain reaction (RT-PCR), were significantly reduced in the valsartan-treated nephritic groups in a dose-dependent manner. * p < 0.05 compared with normal control (NC); # p < 0.05 compared with disease control (DC).
Together, these results indicate that increasing doses of valsartan led to dose-dependent anti-fibrotic effects. The maximal reduction in pathological glomerular fibrosis was seen at the same dose of valsartan (1000 mg/l) that caused a maximal proteinuria reduction. Since some of the measures we employed to assess efficacy were actually less effectively blocked in the group receiving the highest dose of 1500 mg/l valsartan when compared to those in the group receiving 1000 mg/l valsartan but were similar to those in the group receiving 500 mg/l valsartan, it is likely that the true maximally therapeutic dose is between 500 mg/l to 1500 mg/l and very close to 1000 mg/l.
Effects of valsartan on glomerular renin mRNA expression in anti-Thy 1.1 nephritis
Glomerular mRNA expression of renin is shown in Figure 5. Induction of disease significantly stimulated glomerular renin mRNA expression by 3.5-fold (p < 0.05) at day 6. Treatment with valsartan in the drinking water significantly further stimulated glomerular renin mRNA expression in a dose-dependent manner by interrupting a negative feedback loop. A maximal increase of approximately 5.5-fold was seen at a dose of 1000 mg/l or higher dose of valsartan; however, the increase of renin expression in the highest dose of the valsartan-treated group was not significant when compared to that in 1000 mg/l valsartan-treated animals (p > 0.05). These results also confirm that the dose around 1000 mg/l of valsartan in drinking water may reach the maximally effective dose.

Effect of increasing doses of valsartan in drinking water on glomerular mRNA expression of renin in anti-Thy 1.1 nephritis at day 6 (d6). Glomerular mRNA expression of renin was significantly increased in the valsartan-treated nephritic groups at d6 in a dose-dependent manner. * p < 0.05 compared with normal control (NC); # p < 0.05 compared with disease control (DC); § p < 0.05 compared with 250 mgl/l of the valsartan-treated nephritic group.
Effect of valsartan at optimal dose on podocyte injury
From data presented above, it is clear that valsartan reduced albuminuria/proteinuria and fibrotic disease markers in a dose-dependent manner in anti-Thy 1.1 nephritis, with maximal effects observed at 1000 mg/l. It has been shown that proteinuria is often the result of podocyte injury in human diseases and animal models, including anti-Thy 1.1 nephritis.17–21 The protective effect of valsartan (1000 mg/l) on podocytes in anti-Thy 1.1 nephritis was also evaluated.
Although the primary injury, i.e. injection of a complement-activating anti-Thy 1.1 antibody, OX-7, is highly selective for renal mesangial cells, intraglomerular capillaries are destabilized in the course of mesangial cell destruction, leading to capillary dilation and intraglomerular microaneurysms after disease induction. 13 All of these changes will contribute to secondary podocyte damage via altered physical forces and/or potential biochemical alterations of the glomerular basement membrane (GBM),19,22 which may lead to the occurrence of proteinuria in anti-Thy 1.1 nephritis. Indeed, diminished staining of podocin, a critical component of the podocyte slit diaphragm of podocyte, 23 was observed in the nephritic glomeruli on day 6 (Figure 6A-DC) when compared to that in normal glomeruli (Figure 6A-NC). However, staining intensities for podocin were effectively restored after valsartan treatment (Figure 6A-Val). The results of the quantitative analysis of IF staining for podocin are shown on the right of Figure 6A. WT-1 is a nuclear protein specific for podocyte and parietal glomerular epithelial cells in the adult kidney.24,25 WT-1-positive cells within the glomerular tuft excluding the parietal epithelium were counted in anti-Thy 1.1 nephritic rats and the cell number per glomerular was calculated as described in the method. As illustrated by IF staining in Figure 6B, consistent with the staining of podocin, nephritic glomeruli from disease-control rats contained much fewer podocytes than did glomeruli from normal-control rats (6±0.46 vs 16±0.08; p < 0.05). The average number of podocytes per glomerular cross section in nephritic rats treated with valsartan was 50% higher than that in disease controls (11±0.42 vs 6±0.46, p < 0.05).

Effect of optimal dose of valsartan (1000 mg/l) in drinking water on glomerular podocin and Wilms tumor protein 1 (WT-1) protein production in anti-Thy 1.1 nephritis at day 6 (d6). Representative photomicrographs of glomerular immunofluorescent staining for podocin (A) and WT-1 (B) from normal-control rats (NC), disease-control rats (DC) and optimal dose of valsartan-treated nephritic rats at d6 (Val, 1000 mg/l valsartan in drinking water) are shown on the left. Original magnification × 400. Graphic representations of glomerular podocin score and number of WT-1 positive cells are shown on the right. For comparison, glomerular densitometric intensity of podocin staining was set at unity for the normal-control group and other groups’ were expressed as fold-change over this value. * p < 0.05 vs NC; # p < 0.05 vs DC.
By Western blot analyses of the isolated glomeruli from different groups of rats, we also observed that the protein levels of nephrin in nephritic glomeruli, another important podocyte slit diaphragm-associated protein, 26 were substantially down-regulated by 47%, compared with normal glomeruli. Valsartan treatment significantly increased the levels of nephrin by 36.4% (p < 0.05) (Figure 7B). B7-1, a co-stimulation factor of lymphocytes, is synthesized in podocytes in response to slit-diaphragm protein rearrangement and foot process effacement. 27 B7-1 protein levels were also markedly increased in nephritic glomeruli (Figure 7C), which was consistent with the changes of the two slit diaphragm-associated proteins nephrin and podocin. Valsartan treatment reversed the elevated B7-1 protein expression by 62.2% (p < 0.05).

Effect of optimal dose of valsartan (1000 mg/l) in drinking water on glomerular nephrin and B7-1 protein levels in anti-Thy 1.1 nephritis at day 6. (A) The representative Western blots illustrate nephrin and B7-1 protein expression. (B and C) The respective graphs summarize the results of band density measurements. Changes in nephrin (B) and B7-1 (C) protein expression were determined by normalizing against the densitometric intensity of its ß-actin for each sample. For comparison, this ratio was set at unity for normal-control samples and other groups on the same gel were expressed as a fold-increase over this value. * p < 0.05 compared with normal control (NC); # p < 0.05 compared with disease control (DC). Val, 1000 mg/l: valsartan-treated group.
In agreement with protein levels, the gene expression of podocin, WT-1 and nephrin was down-regulated substantially in this model (Figure 8A–8C). Valsartan administration largely preserved the expression of these podocyte proteins. In contrast, as a marker of podocyte injury, mRNA expression of B7-1 was significantly increased in nephritic glomeruli by 240%, which was reversed by valsartan treatment (Figure 8D).

Effect of optimal dose of valsartan (1000 mg/l) in drinking water on glomerular mRNA expression of podocin (A), WT-1 (B), nephrin (C) and B7-1 (D) in anti-Thy 1.1 nephritis at day 6 determined by real-time reverse transcriptase-polymerase chain reaction (RT-PCR) and standardized to glyceraldehyde-3-phosphate dehydrogenase (GAPDH) mRNA levels. Relative values are expressed relative to normal control (NC), which was set at unity. * p < 0.05 vs NC, # p < 0.05 compared with disease control (DC). Val, 1000 mg/l: valsartan-treated group.
Collectively, these data indicated that valsartan treatment given at a dose that maximally inhibited proteinuria and ECM deposition also prevented podocyte dysfunction in anti-Thy 1.1 nephritis by preserving expression of slit-diaphragm proteins and counter-regulating pathogenic molecules involved in podocyte injury.
Discussion
In this study of the anti-Thy 1.1 nephritis model, we observed that valsartan reduced albuminuria/proteinuria in a dose-dependent manner, with the maximal reduction seen at 1000 mg/l of drinking water (40 mg/day, approximately equivalent to 160 mg/kg per day) and no further reduction at a yet higher dose of valsartan (1500 mg/l of drinking water). Together with the previous report that the maximal effect of valsartan in the type 2 diabetic rat model was also seen at their highest tested dose, 160 mg/kg per day, 5 which was the same dose required to maximize proteinuria reduction in the present study, it is likely that the maximally therapeutic dose of valsartan is close to 160 mg/kg per day in animal models. In the present study, we did not measure rat blood pressure. However, it has been repeatedly shown that the model of nephritis in rats is normotensive.12,14,28 The anti-proteinuric effect of valsartan therefore may be independent of blood pressure. In addition, treatment with valsartan did not result in any reduction in Ccr levels even at a high dose (Table 3). The dose-dependent anti-proteinuric effect of valsartan in this model therefore may not be due to any reduction in GFR.
In corroboration with the effects on proteinuria, valsartan in this study also resulted in a dose-dependent improvement in kidney pathology. The valsartan dose that achieved a maximal proteinuria reduction also induced maximal reductions in glomerular over-expression of TGFß1 and PAI-1, two powerful fibrogenic molecules, and the accumulation of ECM protein. We and others have used other Ang II blockers in the same animal model previously,12,29 showing that increasing the dose of enalapril or losartan or telmisartan above the respective maximum antihypertensive dose produced additional antifibrotic effects in nephritic glomeruli. These findings in the anti-Thy 1.1 model are also consistent with the report in the 5/6 nephrectomized rats, showing that a much higher dose of ARB (losartan, 500 mg/kg/d) reduced histological evidence of renal fibrosis more effectively than a lower and conventional dose (50 mg/kg/d) despite a similar effect on blood pressure. 3 All these studies suggest that dosing is very important. At the lower doses of ARB, only a moderate antifibrotic effect was seen. It is likely that these doses are insufficient to block most of the abnormally expressed angiotensin type 1 receptor (AT-1R) in the kidney. Maximal renal protection may require adequate doses of ARB.
We have observed that glomerular Ang II levels are significantly increased in isolated nephritic glomeruli vs. normal glomeruli. 30 The antifibrotic action of valsartan is clearly Ang II mediated. However, the mechanisms underlying the renoprotective effect of valsartan independent of blood pressure lowering are complex and not fully understood. Generally, because Ang II stimulates mesangial cell proliferation,31–33 blocking its action sufficiently with enough dosing of valsartan may maximally reduce proliferation so that the number of activated mesangial cells overproducing TGFß and PAI-1 is reduced and expression of downstream fibrogenic molecules, such as collagen I and FN, is significantly reduced maximally. In addition, it is believed that proteinuria is one of the most important factors leading to the progression of kidney disease. Valsartan sharply reduced proteinuria. The exact mechanisms of this action of ARB have not been completely elucidated. With increasing attention being paid recently to the multiple roles of the podocyte in maintaining glomerular function,17,19,21 several lines of studies have demonstrated that ARB has a potential to protect podocytes from being damaged and thereby promoting reduction of proteinuria.34–37 Indeed, the podocyte injuries caused by the induction of Thy 1.1 nephritis were evaluated in the present study not only by the amount of albuminuria/proteinuria, but also by the expressions of podocin, nephrin, WT-1 and B7-1. The immunohistological staining of podocin and WT-1 and the protein level of nephrin dramatically decreased, and those mRNA expressions for podocin, nephrin and WT-1 also decreased in diseased glomeruli. Podocin and nephrin are reported to be the critical molecules for maintaining the barrier function of the podocyte slit diaphragm.23,26 Dysfunction of podocin and nephrin may cause the consequent podocyte shape changes, such as the effacement of the foot processes, and may attenuate the contractile function of the podocyte against the GBM distention and then leads to loss of adhesion to the underlying GBM and detachment.23,26 B7-1 is a transmembrane protein normally expressed on the surface of B cells and other antigen-presenting cells (APC) cells.38,39 More recently, it has been demonstrated as a de novo synthesized molecule that is involved in the induction of foot process effacement and disruption of the slit diaphragm protein complex in the podocyte in response to a variety of stress conditions. 27 In agreement with the changes of the two slit diaphragm-associated proteins nephrin and podocin, mRNA expression and protein production of B7-1 were significantly increased in diseased glomeruli in this study. Taken together with all of the observations on podocytes, it is conceivable that the marked proteinuria in anti-Thy 1.1 glomerulonephritis resulted from the podocyte dysfunction and even reduction of the podocyte number. These findings suggest that not only mesangial cell lysis and proliferation, but also the disorder of podocyte function, are involved in the development of mesangial alterations and thereby further promote proteinuria and ultimately glomerular sclerosis in anti-Thy 1.1 nephritis. However, the podocytic expression of nephrin, podocin and WT-1 was well preserved and the elevated B7-1 in injured podocytes was counter-regulated when the optimal dose of valsartan was given. Since suppression of RAS markedly lowers glomerular pressure, lessening the mechanical stress with valsartan may directly impose on podocytes. Moreover, it has been shown that Ang II locally causes podocyte apoptosis, podocyte actin rearrangement, increased TGFß1 and PAI-1 levels, and reduction of nephrin levels.37,40–42 Thus, another mechanism whereby valsartan may protect podocytes is by inhibiting Ang II's direct and deleterious effects on podocytes. In addition, there might be a pathogenic cross-talk that occurred between mesangial cell (cause) and podocyte (injury) in anti-Thy 1.1 nephritis.20,21 Ang II may act as the messenger of the injury signal from mesangial cell to podocyte in this model. Blocking Ang II’s action with valsartan may block this potential cross-talk, thereby reducing the response of podocyte to injury. Very recently, Villa et al. also observed that high-dose telmisartan treatment reduced podocyte damage as shown by counting cells positive for both WT-1 and desmin and determining the expression of D1gh, a scaffold protein on podocyte in the same model. 29 Importantly, the protective effect of high-dose telmisartan on podocyte injury strongly correlates with the reduction of proteinuria. 29 Apparently, podocyte protection may explain the reason why therapy with valsartan at the maximally effective dosage markedly decreased albuminuria/proteinuria.
An interesting finding in the study presented here is that after a point, increasing the dose did not lead to further reduction in any measure of disease, which was also seen in our previous study. 12 It therefore seems impossible to completely prevent the fibrotic process with ARB alone at any dose. Together with the data that no additivity was seen when enalapril and losartan were combined at maximal anti-fibrotic doses, 12 these results strongly imply that the critical therapeutic factor for maximizing the renoprotection of ARB is dose optimization before adding other agents to ARB at conventionally anti-hypertensive doses.
Finally, it is noteworthy that supramaximal doses of valsartan did not cause any noticeable side effects in the present model, while reducing both proteinuria and renal fibrosis. However, attention should be paid to the potential side effects of high doses of ARB when a longer-term treatment is employed in patients.
In conclusion, the present study demonstrates a clear dose-dependent therapeutic response for valsartan in the anti-Thy 1.1 rat model. Proteinuria was reduced more effectively at higher doses of valsartan. The maximal antiproteinuric dose of valsartan also conferred a maximal reduction in renal fibrosis, associated with an additional protective effect against podocyte dysfunction. Importantly, these studies further illustrate the concept that ARB at any dose may not be sufficient to completely prevent kidney fibrosis. Combination therapy with other agents that act through different pathways may be crucial.
Footnotes
Acknowledgements
We wish to thank Linda Hoge for her excellent technical assistance with the animal studies.
Funding
The work was supported by a grant from Novartis Pharmaceuticals Corporation (Y.H.) and National Institutes of Health grants K01DK077955 (Y.H.) and R21DK081815 (Y.H.). G.Z. is the recipient of a postdoctoral fellowship grant from Shengjing Hospital Foundation affiliated with China Medical University.
Conflict of interest
None declared.