
Abstract
Domestication is often portrayed as a long-past event, at times even in archaeological literature. The term domestication is also now applied to other processes, including human evolution. In such contexts, domestication means selection for friendliness or prosociality and the bodily results of such selective choices. Both such perspectives are misleading. Using dogs and modern humans as entry points, this paper explores why conceiving of domestication as a threshold event consisting of selection for prosociality is both incomplete and inaccurate. Domestication is an ongoing process, not a moment or an achievement. Selection in breeding, including for prosociality, is a part of many domestication histories, but it alone does not sustain this process over multiple generations. Further, much selection in domestication has little to do with human intention. Care, taming, commensalism, material things, and places are critical in carrying domestic relationships forward.
Introduction
The study of animal domestication is a highly productive area of research for the social and biological sciences, and one of the preeminent topics in archaeology (Kintigh et al., 2014). Despite the prominence of the topic, it is argued here that domestication is often portrayed in a misleading manner, leading to overly simplified narratives of how these complex and variable processes unfolded in the past and how they continue to emerge. Here I focus on two areas of theoretical convergence in the biological and social sciences regarding domestication: that it consists of numerous ongoing processes and that it involves more than intentional human selection for friendliness. The notion that domestication is a process is not new to archaeology (Bökönyi, 1969; Clutton-Brock, 1992). Here, the critique is focused on the fact that domestication involves multiple processes (Lien et al., 2018) and that those processes are ongoing, not discrete past events or thresholds (Larson and Burger, 2013). Second, while selection for friendliness may allow for the initial emergence of some bodily changes of domestication, it is argued that this is not the whole of domestication. Many actants figure prominently in the domestication history of a species, not just humans and our intentions.
Dogs and modern humans provide two entry points to explore these perspectives. I choose these two species because scholarship on them directly illustrates the persistence of problematic notions of domestication. Further, both dogs and modern humans have been described in scholarly literature as self-domesticated. Dogs are a comfortable and familiar species to initiate this discussion, as many scholars and the general public consider them domesticated. They were the first mammals to emerge into a close and long-standing relationship with people, an association which began somewhere in Eurasia during the latter portion of the Pleistocene (Bergström et al., 2020; Freedman et al., 2014; Perri et al., 2021). People can recognize most living dogs as being distinct from their closest living relatives, gray wolves. The predominant theory regarding how this relationship first emerged is referred to as the commensal pathway or the self-domestication model (Coppinger and Coppinger, 2001; Germonpré et al., 2021; Morey and Jeger, 2015; Prassack et al., 2020; Zeder, 2012). This theory proposes that a subset of wolves began to feed on human refuse and those that were least fearful or friendliest persisted in this niche over the long term. They entered the relationship on their own terms. Wolves perhaps experienced genetic and phenotypic change by living commensally for many generations in an environment that favored friendliness—human intentionality did not play a role, at least initially. Does this then mean that the domestication of dogs was completed when such bodily changes first emerged? Even some recent archaeological literature on dogs (described below) seems to follow this line of thinking, describing dog domestication as being completed in the Pleistocene—it was (and is) not an ongoing process.
An equally puzzling understanding of domestication can be found when this term is applied to human evolution. Scholars in many disciplines have argued that modern humans are self-domesticated (Boas, 1911; Brüne, 2007; Cieri et al., 2014; Fischer, 1914; Gould, 1977; Hare, 2017; Leach, 2003; Lorenz, 1940; Raghanti, 2019; Thomas and Kirby, 2018; Wrangham, 2021). This proposal also recently gathered significant public attention through the best-selling book Survival of the Friendliest (Hare and Woods, 2021). Such influential lines of thinking are direct evidence that domestication is still often understood in a narrow way—bodily change produced by selection for friendliness. In the most recent versions of this proposal, the emergence of modern humans involved our selection for friendliness (low levels of reactive aggression or prosociality) over aggression. This in turn led to humans having bodily traits shared with other domestic mammals, known as the domestication syndrome. This process of selection also led to the emergence of distinct behaviors or cognitive abilities in our species. However, unlike most other understandings of domestication, human self-domestication is intra-species. Most recent discussions of human self-domestication are directly tied to the famed Russian Farm-Fox Experiment, which has been said to demonstrate that selection for friendliness alone produces the bodily traits that constitute the domestication syndrome.
Domestication is more than a discrete period of selection for prosociality. If domestication is ongoing, as is argued in a suite of archaeological, genetic, and anthropological literature, no animal can be domesticated, but rather domestic, a work in progress. A selective setting that favors prosociality, even when that setting includes human intentionality, is not sufficient to sustain domestication, including the bodily changes that might appear from it. Influencing breeding, including for prosociality, is part of a much more complex assemblage that constitutes these processes and relationships. Further, even intentional human acts in domestication involve selection for factors beyond just prosociality. To begin, a useful starting point is an assessment of how domestication has been defined in the literature. Following this, dog and modern human domestication are examined.
Thinking through domestication definitions
Scholars have not reached a consensus on how domestication should be defined, but many attempts have been made to do so. The term is used to describe the histories of species as variable as rice, silkworms, salmon, turkeys, and horses, particularly their long-term histories with people. Clearly, scholars are asking a single word, domestication, to do much work across highly variable contexts. Multiple social and evolutionary processes are being described, not a single one (Lien et al., 2018). Limiting ourselves to just animal domestication fails to simplify matters, but repeated themes are present (only some of which are addressed here).
First, a common element of domestication definitions has long been human control of (or influence on) animal reproduction over successive generations (Bökönyi, 1969; Clutton-Brock, 1989, 1992; Reitz and Wing, 2008; Russell, 2011; Vigne, 2011; Zeder, 2015). As Clutton-Brock (1992: 79) stated, “It is complete when the new population is permanently isolated from the wild species and its breeding, organization of territory, and food supply is under total human control.” Breeding control is typically said to involve isolation of subsets of populations, pairing select individuals for breeding, or culling animals prior to sexual maturity. Depending upon the animal under consideration, such control might emerge millennia after its domestication is initiated or from its very outset. Whether it is early or late, human intentionality is often invoked. The intentional control element in domestication definitions distinguishes domestication from other intimate human–animal relationships. Commensalism and taming, for example, are common counterexamples to domestication (Bökönyi, 1969; Russell, 2011: 209, 216; Bar-Oz et al., 2014). Animals in a commensal relationship with humans are adapted to living in human-dominated niches, but we make no interventions in their breeding. Taming involves a relationship between a specific animal and human spanning only that animal’s lifetime, where the animal is repeatedly interacted with, but also often fed and provided care. Both can occur and not lead to domestication. These proposed differences between domestication, commensalism, and taming are less discrete when considering dogs and many other domestic animals, as will be shown below.
Most importantly here, biological and social sciences have repeatedly argued against the concept of human control of animal breeding in domestication. For example, geneticists contest notions of continual breeding control (and isolation) because of evidence for repeated introgression in the past between wild and domestic populations (see Frantz et al., 2020). Many dogs, pigs, horses, goats, and reindeer made their own breeding choices, at least at times (Anderson et al., 2017b; Ameen et al., 2019; Daly et al., 2018; Frantz et al., 2015, 2019; Park et al., 2015; Schubert et al., 2014; Verdugo et al., 2019). Anthropologists also critique the narrative of human control and mastery of animals in domestication definitions, noting that it is deceptively unidirectional, rendering animals passive in these processes and relations (Anderson et al., 2017a; Cassidy, 2007; Lien, 2015; Losey et al., 2018a; Mlekuž, 2013; Stépanoff, 2017; Stépanoff and Vigne, 2018; Swanson et al., 2018). Their first-hand observations of domestications in action show that humans are not always in control of such relationships, including breeding decisions. In contrast to ideas of mastery and control, anthropologists often describe domestication (but surprisingly rarely actually define it) using terms such as inter-dependence, mutualism, co-existence, and cohabitation.
Archaeology has also long made arguments against the centrality of human intent in domestication (Zeuner, 1963; Rindos, 1984; O’Connor, 1997; Zeder, 2012; Stépanoff and Vigne, 2018; Bogaard et al., 2021). In a well-known example, Zeder (2012) proposed three potential scenarios through which animals became domestic, including one termed the commensal pathway. This scenario involved a subset of a population recurrently feeding on human refuse or in human niches. Such individuals tended to be less aggressive and to have reduced flight tendencies and high stress tolerances. Through such sustained contacts between this subset of animals and people, “these animals develop closer social or economic bonds with their human hosts, who begin to derive some tangible benefit from their association” (Zeder, 2012: 173). Over time, the group presumably partially drifted away from members of its species living outside this niche, developing somewhat distinct morphologies, genomes, and behavior. In this scenario, the selective actants are human places and the animals themselves, with intentional human influence on breeding emerging far later (with some species, millennia later) in the history of the relationship.
In fact, many recent conceptualizations of domestication argue that human intent is but one of many actants shaping long-term domestication histories. For example, Larson et al. (2014: 6140) have defined animal (and plant) domestication in such a manner, stating, “domestication can be generally considered a selection process for adaptation to human agro-ecological niches and, at some point in the process, human preferences.” This definition reflects the analytical reality in archaeology that human intention and preference in animal breeding are difficult to definitively identify. In many cases, the observed diachronic patterns in genotypes and phenotypes potentially have multiple etiologies. Niche construction theories make related claims, describing how plants, animals, people, and landscapes together act as selective forces in domestication, but also assist in perpetuation of specific domestic relationships (McClure, 2015; Zeder, 2016, 2017). In both examples, domestication is unambiguously considered an ongoing relationship rather than an initial period during which these relationships began or are first materially evidenced. As Larson and Burger (2013: 198) have stated, “Because the evolution of domestic animals is ongoing, the process of domestication has a beginning but not an end.” Human intent is not irrelevant in domestication, but rather is often intermittent, and not the manner in which all domestic relations emerged and were sustained.
Second, domestication is commonly said to involve change in animal genetics, form, behavior, and development (Bökönyi, 1969; Clutton-Brock, 1989, 1992; Ingold, 1980; Larson, 2011; Reitz and Wing, 2008; Russell, 2011; Zeder et al., 2006). Morphologists and geneticists are most interested in documenting such changes and the selective forces and bodily processes that bring them about. They are what need explaining within those disciplinary frameworks. The relations, enskilments, material entanglements, and landscapes of specific domestication histories are of far less interest. Archaeology in some ways also does not begin with a focus on the relations of domestication but rather with an assessment of bodily (skeletal) form. In zooarchaeology, for example, a methodological first step is identification of animal skeletal remains to specific taxa, some of which are domestic, based on their shapes and sizes. This is a starting point for other analyses and explorations. In short, for biology and archaeology, the focus on bodily difference is an operational need. It is a means of getting at the evolutionary histories of long-dead animals or for initially categorizing their remains as an entry point for additional research. Just as critically, while attention to bodily change is where much work on domestication begins, genomic and phenotypic change is not domestication—it is evidence of it.
Studying bodily change as evidence for domestication raises other important issues. Tracing such trends over archaeological time scales often reveals continual and multidirectional change. There is no threshold in either form of data that signals a discrete transition from wild or commensal to domestic, from incipient domesticate to fully domesticated, nor from domestication to some other post-domestication process. Such distinctions are imposed onto evolutionary histories and continuities of practice. These arbitrary cuts are often more about data resolution and reliability of analytical techniques than empirical differences in state. In other cases, the issue seems to be the inferred extent of intentional human selection in animal breeding (e.g., Vigne, 2011; Frantz et al., 2020). Practices described as husbandry, for example, presumably involve extensive and often widespread goal-oriented breeding control, while the other parts of domestication histories consist of carrying out these same processes less often and less strictly. If this interpretation is correct, then differences in degree are being described, not differences in kind.
It is also unclear how the bodily changes of domestication are generated. Many such changes in mammals have been termed the domestication syndrome, but even the existence and characteristics of this set of traits recently have been questioned (Lord et al., 2020, Wheat et al., 2020; but see Trut et al., 2020; Wright et al., 2020; Zeder, 2020). Specifically, syndrome traits such as floppy ears, varied coat colors, and shifts in mating seasonality may not all originate from the same causes. Some may have appeared late in a species’ domestication history, and not all such traits may appear in all domestic mammals, let alone in birds and fish. The most widely cited assessment of potential causative factors underlying the emergence of the domestication syndrome is the decades-long Russian Farm-Fox Experiment (Figure 1) (Belyaev, 1979; Trut, 1999). The core proposal of this experiment was that selection on behavior, particularly tameness, produced the developmental, morphological, and reproductive traits that constitute the syndrome. Such traits are part of an interconnected system that can be rapidly affected by strict selection for prosociality over the course of fewer than 100 generations. To be clear, the Farm-Fox Experiment does not demonstrate that intentional human selection for friendliness alone brings about domestication. At the same time, the experiment itself is a particular domestication history, albeit a very recent one. A fox from the Russian Farm-Fox Experiment. In 2014, the fox was living as a pet outside of Irkutsk in a small garden area (dacha) but was relentlessly pursued by feral dogs. The fox was eventually surrendered to a zoo to prevent it from being killed by the dogs. Photograph by the author.
Regardless of whether the core proposals in the Russian Farm-Fox Experiment are correct, it is important to recognize that this particular domestication itself involves far more entanglements and processes than simply selection for friendliness. Dugatkin and Trut’s (2017) description of the experiment shows that this domestication was brought about by the efforts of caretakers, scientists, and the foxes themselves, an elaborate set of facilities that housed the experiment, an academic system that supported all of this for decades, and even Cold War politics. Further, processes beyond the experiment were significant forces in this particular domestication history. Specifically, when funding for the project dwindled during an economic collapse in the late 1990s, the selective environment of the foxes radically changed. With very few resources left to hire staff or purchase food, many of the foxes starved and others were euthanized for their furs, reducing the population from ∼700 to 130 animals. Without these painful and traumatizing acts (and a number of other remarkable efforts by the foxes’ caretakers), potentially all of these foxes might have perished and their domestication have been terminated. Selection for prosociality was a critical part of this fox domestication, but it was far from the whole of it.
Additionally, characteristics other than tameness were and are selected for in animal domestication histories, including (but not limited to) their appearance, strength, speed, endurance, dietary flexibility, reproductive rate, body size, ability to learn and communicate, song, and fat content. Even aggression surely has been a useful trait in some domestications. Predictable, targeted aggression could be beneficial for dogs that work in hunting, herding, and livestock protection, for example. Further, at least in modern cattle, selection for docility also correlates with decreased brain size (Balcarcel et al., 2021). While caution is warranted in linking brain size to intelligence, it is possible that there are negative cognitive consequences of extreme selection for tameness. These might be inhibitive if domestic animals were being asked to perform particularly complex tasks in their lives with humans.
Domesticated dogs?
The ancestors of modern dogs diverged from gray wolves in Eurasia toward the end of the Pleistocene (Bergström et al., 2020; Freedman et al., 2014; Perri et al., 2021). This process began early enough that at least five major lineages of dogs had emerged by the start of the Holocene (Bergström et al., 2020), and some dogs had moved with humans into the Americas by ∼15,000 years ago (Perri et al., 2021; da Silva Coelho et al., 2021). The leading model for how this human–dog relationship began is a version of Zeder’s commensal pathway scenario (Coppinger and Coppinger, 2001; Larson and Fuller, 2014; Zeder, 2012). Namely, a subset of wolves, particularly individuals that were less fearful of humans, evolved to specialize in feeding on our meaty leftovers. They choose to repeatedly inhabit human places and eat our foods, perhaps for millennia. At some point in this process, these small populations of canids diverged from those living distantly from humans. They were undergoing selection for prosociality through inhabiting human-dominated places—by being commensal. This adaptive process is seen in the model as a prerequisite for eventual human intervention in their breeding, which emerged later in the history of the relationship. When in the relationship such interventions occurred is anyone’s guess. Recent archaeological dog literature refers explicitly to such a model as the self-domestication hypothesis (Germonpré et al., 2021; Morey and Jeger, 2015; Prassack et al., 2020).
Other competing models of dog domestication also persist, including the long-standing wolf pup–adoption model recently reinvigorated by Germonpré et al. (2021; see also Serpell, 2021). This is a human-initiated scenario, with the original intent not being to domesticate animals but rather to keep individuals as companions. In the model, pups younger than 12 days of age (at which point they open their eyes) are taken and raised in human settlements. Lactating human women nurse the pups and provide them with other supplemental food. Children assist the women in socializing and caring for the pups, including keeping them warm. In other words, the pups are tamed and provided care—they become parts of hybrid human–wolf communities. Prolonged contact between people and the pups led to close social bonds between them. Again, there was selection for prosociality, with particularly aggressive individuals being shunned or killed. Females that grew to maturity in such settings had fewer competitors, which resulted in lower concentrations of select hormones. These changes disrupted embryonic development, affecting the behavior and morphologies of subsequent generations. Taming, culling, and selection for prosociality occurred intermittently from this point forward.
It is important to recognize that both of these models attempt to describe and explain the beginnings of dog domestication, not its entire history. The pathway metaphor is potentially misleading in this regard if one considers a pathway a singular route for getting to a discrete end point. The commensal or self-domestication scenario for dog domestication seems to end with human intervention in breeding, suggesting a completed process. As previously described, traditional definitions of domestication advocate for human intervention in breeding, not just commensalism, as the core identifying feature. With this in mind, the model clearly describes a starting point, not the full history of the relationship. In Zeder’s (2012: 171) own words, the scenarios are ways that “most animal domesticates followed into domestication,” not how they became domesticated, completed projects. Perhaps to better employ a pathway metaphor for dog domestication, one would have to imagine such paths as convoluted ones with many trails that diverge and converge, some of which abruptly end or turn back, and others that continue to form.
In the wolf pup–adoption model, Germonpré et al. (2021) state repeatedly that they are attempting to describe the beginning of dog domestication, not its entirety. There is recognition here of a longer-term set of processes. Much of their model involves care and taming. These efforts too are not confined to the early histories of dog domestication. One can very strictly control the breeding of an animal, but without socializing it (taming it), the animal can be difficult to handle, and cooperative tasks such as herding, hunting, or hauling would be nearly impossible. Particularly with working animals, socializing/taming is required with every generation, not just the first. Selection for prosociality makes such processes easier, but it alone does not bring them about. Care too is often a domestication requirement, not something external to the process. Providing food, security, habitat, shelter, health care, and so on are part of what sustains domestication, making it multi-generational. Such efforts and struggles are part of what allows for the persistence of the genotypic and phenotypic changes of domestication (Losey et al., 2021).
Regardless of the models and metaphors one prefers, archaeological and genetic data demonstrate that dog domestication, even defined strictly, is ongoing and involved more than selection for prosociality. For instance, some dogs have developed the capacity to digest starches, which is marked by increased copy numbers of the AMY2B gene (Axelsson et al., 2013). Bergström et al. (2020) show that high copy numbers of this gene were not widespread in Eurasian archaeological dogs until several thousand years after the emergence of starch-rich agriculture—well into the Holocene. Conversely, dogs in many hunter-gatherer communities exhibit low copy numbers of the gene long after this time. This particular adaptation is not merely a byproduct of selection for prosociality but rather seemingly an outcome of long-term habitation in starch-rich places—being commensal in particular food environments. As Bergström and colleagues (2020: 561) state, “These results suggest that selection for increased AMY2B copy number did not take place during the early stages of domestication.” Instead, this adaptation is an outcome of their continuing domestication, one that is ongoing ∼10,000 years following dogs’ first divergence from their wolf kin (Perri et al., 2021). Bergström et al. (2020) also provide evidence of dog introgression into wolves in Eurasia, suggesting their reproduction was not fully controlled by humans, again well into the Holocene. Even by Clutton-Brock’s (1992) strict definition, dogs are not fully domesticated.
Clearly, archaeologists, anthropologists, and geneticists all have argued (in some cases for decades) that domestication is an ongoing and variable set of processes, not a completed, singular one. The Russian Farm-Fox Experiment sought to understand how the domestication syndrome first emerged but was not an attempt to explain the whole of domestication, even for canids. The self-domestication and pup-adoption models for dogs also clearly address origins rather than long-term domestication histories. Despite all of this, it is still common to see domestication understood as a long-past threshold marked by an initial period of bodily change, even in very recent archaeological literature. For instance, Bethke and Burtt (2020) compiled a series of papers entitled Dogs: Archaeology Beyond Domestication. The contributors to the volume address dog sacrifices, diets, habitual activities, and relationships with other domestic animals, with most cases being from the Holocene. Titles are meant to draw one’s attention, but given that the processes described involved adaptions to human niches, intentional and unintentional selection, socialization, provisioning, and care over multiple successive generations, domestication is being described. Dog origins are not described in the volume, and thus, the contributors seem to be addressing post-domestication processes and relations. Ameen et al. (2019) document genetic and morphological patterns in dogs from Russia and the North American Arctic to trace the origins and dispersal of Inuit dogs and dog sledding. The authors identify a previously unknown mitochondrial group termed clade X, which they document in dog remains from Primorye, Chukotka, and Alaska. Bayesian analysis suggests the clade emerged 10,000 to 5,300 years ago. The authors state, “The comparatively recent origin of this clade, compared to some of the more prominent mitochondrial clades, suggest the X clade was not present in dogs at the time of domestication” (Ameen et al., 2019: 7). This tells the reader that dog domestication was finished prior to 10,000 years ago—it was a completed process by the time clade X emerged. Finally, two papers have documented the development of relatively large dogs in parts of western North America during the Holocene, likely for their roles in transport (Welker, 2020; Welker and Byers, 2019). The emergence of these dogs is not described as an outcome of domestication, and Welker and Byers (2019: 88) state directly, “It is generally accepted that dogs were domesticated sometime before 15,000 BP in Europe and 12,500 in East Asia.” The clear inference in all of these cases is that dog domestication was a threshold crossed in the Pleistocene.
Domesticated ourselves?
The proposal that modern humans are domesticated can be traced back centuries (Brüne, 2007), with Boas (1911) perhaps the first scholar to describe humans as self-domesticated. What such notions entail differs somewhat among current scholars, but a common claim is that modern humans share phenotypic traits with other domestic animals, and these emerged through our selection for prosociality. My aim is not to assess the validity of such claims but rather to question if the term domestication is aptly applied here. Does this line of thinking conflate domestication with selection for prosociality, as suggested by Morey and Jeger (2015)?
Helen Leach (2003) reinitiated recent interest in human self-domestication. Her application of domestication to human evolution is broader than what appears in later literature on the topic. At the time, the idea that selection for prosociality generates the domestication syndrome was far less prominent than today. In her model, domestication is clearly used as a heuristic device rather than a literal description (Leach, 2003: 359). Human-built or modified environments helped to bring about bodily changes in humans that parallel those seen in domestic mammals. The model recognizes that domestication as defined at that time involved not just the outcomes of intentional breeding control, but also adaptations to human-dominated niches. Some of the actants affecting our evolution in her model are other species of human-modified environments, including domestic plants and animals. In this sense, there is a multi-species, multidirectional, and niche construction element to the model, although she uses none of these terms. Selection for tameness or prosociality is evident in the paper, but not a major focus.
In subsequent scholarship, self-domestication is selection for prosociality and its bodily outcomes, full stop. Hare (2017), Cieri et al. (2014), and Wrangham (2019, 2021) all propose that humans selected for prosociality (or against forms of aggression) during portions of our evolution, which in turn was closely linked to changes in our behavior, including our ability to cooperatively communicate. Such psychological and behavioral changes are said to be a part of the domestication syndrome. Many of the syndrome’s other traits, such as changes in morphology and development, also emerge in our species as a result of this selection process. Variations of the model are used to explain aspects of our evolutionary history, from the emergence of Homo erectus, to the widespread appearance of symbolic behavior, to the development of collaborative conspiracy. Essentially the same process of self-domestication is also said to account for the evolution of phenotypic differences between bonobos and chimps (Hare et al., 2012).
This line of thinking, now highly influential, stems directly from the findings of the Russian Farm-Fox Experiment. As stated earlier, that experiment did not claim to be describing or explaining domestication but rather how a set of phenotypic traits first arose in some mammals undergoing selection for prosociality—how the syndrome first emerged. Regardless, support for the human self-domestication hypothesis also has been taken from very similar experimental breeding programs with mice, rats, and mink (Gariépy et al., 2001; Kharlamova et al., 2000; Singh et al., 2017; Trut et al., 2000). In other cases, self-domestication is said to be evident through differences in cranial morphology between urban and rural foxes (Parsons et al., 2020) or in free-ranging mice unintentionally selected for tameness due to recurrent exposure to humans (Geiger et al., 2018). Further, even the early domestication of dogs is also invoked in human domestication models, a process also referred to as self-domestication (Hare, 2017). While the term domestication is pervasive in this literature, the terms self-domestication and domestication do not refer to the same processes or relationships, even for some prominent proponents of human self-domestication models. Wrangham (2021: 5), for example, states, “Domestication and self-domestication are thus different process, but the self-domestication hypothesis proposes that they share important underlying commonalities in biological mechanisms.” In this version of self-domestication, domestication is redefined as its “biological consequences,” specifically as a “reduction in a species’ propensity for reactive aggression” (Wrangham, 2021: 5). Wrangham (2021) explicitly recognizes that outside most self-domestication literature, domestication has been defined very differently—as various forms of relationships involving care, ownership, breeding control, and so on.
This use of the term domestication as shorthand for selection for prosociality is misleading in many ways, particularly because multi-species relationships are often eliminated from discussion (Morey and Jeger, 2015: 425). Virtually all other domestication definitions have multi-species relationships as one of their critical elements. An additional issue is that the only selective choices made in this understanding of domestication relate to friendliness and not the suite of other behavioral and bodily traits that have been selected for over time in the domestication histories of dogs and other animals (Morey and Jeger, 2015). Domestication in this line of thinking is the generation of the syndrome. Our attention is drawn to the symptoms, not the disease, to carry this metaphor forward. It is far more direct and accurate to term the human self-domestication proposal the “prosociality hypothesis,” and recognize that the comparisons with domestication are about bodily outcomes rather than the same processes. Domestication has often involved selection for prosociality, but it is far more than this, even in terms of intentional human choice. Dachshunds, draft horses, and farmed carp are not with us simply because friendly individuals bred with one another in the distant past.
Domestication emplaced
Animal domestication has been difficult for academics to pin down because it is not a discrete, singular process. The term encompasses complex and variable sets of processes and relationships that can be best understood by considering the specific entanglements within which they unfold (Losey et al., 2021; Mlekuž, 2013). Those entanglements involve selection for various attributes, a suite of actants that are selective forces, and numerous efforts, things, and places that sustain domestic relations over the short and long term (Figure 2) (Anderson et al., 2017a; Losey et al., 2018b, 2021; Mlekuž, 2013; Swanson et al., 2018). Figure 2 depicts some of the factors that can come into play in animal domestication histories. Not all factors shown are relevant at all times with all species, but also no single factor listed here operates independently. Emplacing domestication entails coming to understand these processes within their contexts—to understand how the relationships emerge and are sustained, and the places, things, efforts, choices, and commitments they entail. Understanding the bodily changes of domestication is critical, but so too are the other elements involved in these processes. Some potential factors playing roles in the domestication of animals. Not all are critical at all times in any given domestication history, and none operate independently.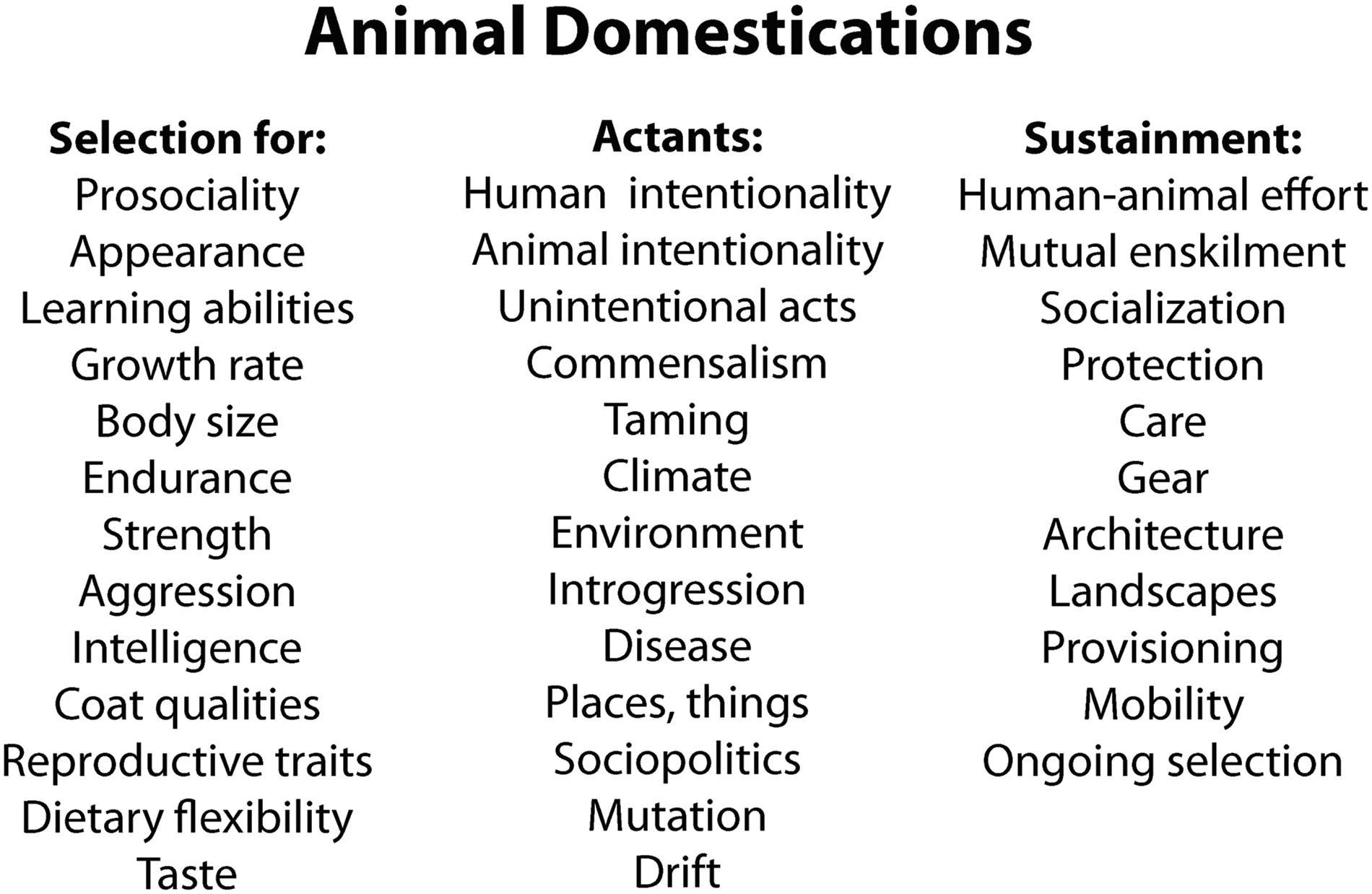
At times, human intentionality can be paramount, but animal domestication by definition must involve nonhuman agents (the animals in question) that can make their own decisions, including in mate choice. Additional actants are always at work, ranging from climate and environment to the built things that enable the social lives, working relations, and mobilities of human–animal communities. These are not merely background or epiphenomenal to the bodily processes of domestication—the latter never occur in isolation, outside of histories, places, and relations. Unintended actions of humans, animals, and the broader worlds within which they are enacted play critical roles. We are often less in control than we might imagine. For example, diseases and droughts reshape animal populations, dramatically reducing both their numbers and diversities. Our histories of colonization and dispersal radically change domestication histories, regardless of their causes. With dogs, such processes have resulted in novel intermixing of dog populations in some areas and the near complete termination of others (Ameen et al., 2019; Bergström et al., 2020; Feuerborn et al., 2021). Humans and domestic animals are just two of many actants at work in the unfolding of domestication, and like all forms of evolution, these processes too involve various entanglements, not just one hand directing a species to a known outcome.
Selection for prosociality is critical in some animal domestication histories, and not just at their beginnings. We continue to make selective choices related to the friendliness of animals. However, prosociality is not the only factor at play, even when considering intentional human breeding interventions. For example, we do not just live with friendly dogs, but also ones that can cooperate in hunting, pulling sleds, and defending our places. Selection for prosociality has in part made such relations feasible, but so too has selection for targeted aggression, persistence, endurance, and particular body sizes. The same might be said for racehorses, dairy cattle, or merino sheep. Their emergence and particular social roles in societies unequivocally have involved intentional selection for factors beyond prosociality. Friendliness can be important, but it is not everything.
Some domestication histories likely began with commensal relationships, but inhabiting human-dominated places continued to affect those histories long after their beginnings. Other domestications probably began through taming, but efforts to care for and interact with animals characterize entire domestication histories, not just their starts. Taming, commensalism, provisioning, and care all are parts of the struggles that help to allow the bodily and behavioral outcomes of selection to be sustained. Moments of selection alone cannot do this. Our gaze, both in archaeology and biological sciences, often focuses on the bodily outcomes of selection because of our operational needs, but those outcomes are not domestication—the syndrome is not the processes or relationships that actually constitute domestication.
Humans and domestic animals must learn to be with one another in various ways and places, and the natures of these mutual enskilments are diverse, again pointing to the multiple nature of domestication. For example, a relationship with a salmon held in a pen in a Norwegian fjord involves a far different way of being together than does working with a draft horse in a field. Saying that both are domestic species actually tells us very little (Losey et al., 2018b; Mlekuž, 2013). Such enskilments are also selective processes, as failing at those relationships can result in escape, injury, or death. Take the example of a foal that was conceived through an intentional pairing of parents that themselves were outcomes of long-term selection for being draft animals. That pairing alone does not make the foal a draft horse, nor does it ensure that its characteristics are passed to future generations. The foal requires care and sustenance to reach reproductive age, and to be a draft horse has to learn that task by repeatedly engaging in it with specific people, gear, landscapes, and so on. How this relationship emerges clearly could have evolutionary outcomes—horses that become skilled at this way of being might well be chosen to sire subsequent generations. This is domestication enacted and practiced, with even choices involving breeding selection emerging through relationships and engagements, not in isolation.
The proposal that the domestication syndrome first arose through selection for prosociality is captivating, and it is unsurprising to see these notions creeping into explanations for evolutionary trends in many species. At the same time, self-domestication is a misleading term for describing such processes, whether they involve humans, dogs, or other animals. It is important to recognize that domestication remains something of a sorting device, at times partnered closely with ideas of nature and culture (Lien, 2015). Thinking domestication as a past threshold event orders societies as creators or borrowers, transformers or the transformed. Such categorical lines of thinking leave little room for actually exploring the emergent processes, relationships, things, places, and bodily changes that constitute specific domestication histories (Mlekuž, 2013).
Finally, archaeological practice can benefit from the realization that domestication is ongoing and more than selection for prosociality. First, when recognizing these two facts we are better enabled to study the processes and relationships of domestication beyond their origin places and periods (Losey et al., 2018a). The topic is not “owned” by those working on the supposed earliest of these entanglements, but rather can be pursued by a diversity of scholars in various contexts. Second, because domestication involves places and things, it can be explored through artifacts, landscape modifications, art, and so on. We are not restricted to the study of bodies. Third, our research on domestication will become more relevant to the present world, as we are no longer describing our work as studying a past phenomenon but rather a continuing set of practices that still affect our lives, evolution, and environments.
Footnotes
Acknowledgements
The inspiration for this paper developed through extensive conversations with Marianne Lien, David Anderson, Tatiana Nomokonova, and several Nenets families over the past several years. I am grateful for their patience, inspiring words, critiques, and friendship. Special thanks are offered to Tatiana Nomokonova for comments on an earlier draft of the manuscript.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Funding for this project was provided by the Wenner-Gren Foundation (SFR1725).