
Abstract
Alzheimer's disease (AD) is a neurodegenerative disorder primarily characterized by cognitive decline. Its pathogenesis is complex, involving multiple pathological processes, including amyloid-β (Aβ) deposition, neuroinflammation, and synaptic dysfunction. In recent years, the role of mesenchymal stem cells (MSCs) in AD therapy has garnered significant attention. MSCs, through their multi-directional differentiation potential and paracrine effects, exhibit remarkable neuroprotective and anti-inflammatory properties, influencing AD progression. This review summarizes the potential mechanisms and effects of MSCs in AD treatment and explores precision therapeutic strategies based on MSC modulation.
Keywords
Background
Pathological mechanisms of Alzheimer's disease
Alzheimer's disease (AD) is a progressive neurodegenerative disorder characterized by memory loss, cognitive deficits, and behavioral changes that culminate in functional disability. 1 The pathogenesis of AD is multifactorial, involving a complex interplay of genetic and environmental factors, with the amyloid-β (Aβ) cascade and tau protein pathology representing two core pathological mechanisms.2–4 The Aβ cascade hypothesis has posited that the imbalanced processing of amyloid-β protein precursor (AβPP), favoring the amyloidogenic pathway, leads to the overproduction and accumulation of Aβ peptides, particularly the aggregation-prone Aβ42.5–7 These Aβ species oligomerize and deposit into senile plaques, primarily in regions critical for cognition like the cortex and hippocampus.8,9 Crucially, both soluble Aβ oligomers and plaques exhibit direct neurotoxicity, disrupting neuronal membrane integrity, ion channel function, and calcium homeostasis, thereby impairing synaptic communication and neuronal survival.10,11 Furthermore, Aβ pathology acts as a potent trigger for chronic neuroinflammation by activating microglia and astrocytes, leading to a persistent release of pro-inflammatory cytokines (e.g., tumor necrosis factor (TNF)-α, interleukin (IL)-1β, IL-6) that exacerbates neuronal damage and dysfunction.12–16 Concurrently, tau pathology unfolds through the hyperphosphorylation of the microtubule-associated protein tau, causing it to detach from microtubules and aggregate into intracellular neurofibrillary tangles.17–19 This process destabilizes the neuronal cytoskeleton, cripples intracellular transport, and ultimately leads to synaptic failure and neuronal death.19–21 The progression of tau pathology, which spreads through the brain in a characteristic pattern, strongly correlates with the clinical decline in AD. 22 The interplay between Aβ, tau, and neuroinflammation creates a vicious cycle that drives disease progression. These core pathologies converge to cause synaptic dysfunction and synaptic loss, a key morphological correlate of cognitive impairment, and culminate in widespread neuronal loss and brain atrophy, particularly in the hippocampus and cortex.23–26
In summary, the pathogenesis of AD is driven by the synergistic toxic effects of Aβ and tau pathologies, which ignite chronic neuroinflammation and directly assault synaptic and neuronal integrity. Despite this advanced understanding, effective treatments remain a challenge, highlighting the need for multi-target therapeutic strategies like mesenchymal stem cell (MSC) therapy.
Role of mesenchymal stem cells in Alzheimer's disease treatment
MSC therapy has brought new hope for the treatment of AD. MSCs exhibit pleiotropic therapeutic effects through multiple pathways, including neuroprotection, neuroregeneration, anti-inflammation, and immunomodulation. Preclinical studies have demonstrated the positive therapeutic effects of MSCs in AD animal models, and preliminary clinical trials have also indicated that MSCs are safe in treating AD patients and may offer certain clinical benefits. Given these promising preclinical and preliminary clinical findings, MSC therapy is expected to become an important component of future comprehensive AD treatment regimens.
Neuroprotective and regenerative effects of MSCs in AD treatment
AD is a neurodegenerative disease that is characterized by neuronal loss, synaptic dysfunction, and cognitive decline.27,28 Its complex pathogenesis involves multiple interconnected factors, namely Aβ deposition, tau hyperphosphorylation, and neuroinflammation.14,29,30 MSCs, owing to their multipotent differentiation potential and paracrine properties, exhibit unique advantages in the treatment of AD.
MSCs differentiation into neural cells
MSCs are a population of adult stem cells capable of self-renewal and multilineage differentiation. 31 They can be isolated from various tissues, including bone marrow, adipose tissue, and umbilical cord, each offering distinct advantages in terms of accessibility and proliferative capacity. 32 While MSCs possess multipotent differentiation capabilities in vitro, their primary therapeutic contribution in AD is not through direct replacement of lost neurons or glial cells. The long-held notion that MSCs can robustly transdifferentiate into functionally integrated neural cells within the complex AD brain environment remains controversial and lacks conclusive evidence.33,34 Although early in vitro studies reported that MSCs could adopt neuron-like morphologies and express neural markers under specific chemical induction, these findings have not been consistently replicated in subsequent in vivo experiments. 35 Crucially, there is a scarcity of evidence demonstrating that transplanted MSCs stably mature into electrophysiologically active neurons that achieve long-term structural and functional integration into host neural circuits.36,37 Rather, it is now widely accepted that the therapeutic benefits of MSCs originate predominantly from their potent paracrine actions.36–38 This “paracrine hypothesis” posits that MSCs act as biological factories, secreting a rich cocktail of bioactive molecules that collectively modulate the disease environment. Upon transplantation and homing to sites of pathology like the hippocampus and cortex, MSCs do not primarily replace dead cells. Rather, they create a supportive microenvironment conducive to repair and regeneration. Key paracrine effectors include: immunomodulatory factors that suppress chronic neuroinflammation, a major driver of neuronal death in AD.39,40 MSC-derived exosomes, which serve as key nanocarriers, deliver functional nucleic acids (e.g., miRNAs) and proteins that directly activate regenerative programs in resident brain cells. 36 A critical manifestation of this paracrine activity is the enhancement of endogenous neurogenesis. MSCs promote the proliferation and survival of neural stem/progenitor cells in the neurogenic niches of the adult brain, such as the subventricular zone and the hippocampal subgranular zone.41,42 By mitigating the inhibitory inflammatory milieu and providing pro-survival signals, MSCs empower the brain's own repair mechanisms to generate new neurons and glial cells, thereby alleviating neuronal damage (Table 1).39,40
Pleiotropic mechanisms: summary of MSC action pathways in Alzheimer's disease treatment.

Aβ: amyloid-β; BDNF: brain-derived neurotrophic factor; CSF-1: colony-stimulating factor-1; GDNF: glial cell line-derived neurotrophic factor; IDE: insulin-degrading enzyme; IDO: indoleamine 2,3-dioxygenase; IL: interleukin; MSC: mesenchymal stem cells; NEP: neprilysin; NGF: nerve growth factor; PGE2: prostaglandin E2; TGF-β: transforming growth factor-β; TNF-α: tumor necrosis factor-alpha; VEGF: vascular endothelial growth factor.
MSC secretion of neurotrophic factors
MSCs exert neuroprotective effects in the treatment of AD by secreting multiple neurotrophic factors, including brain-derived neurotrophic factor (BDNF), nerve growth factor (NGF), glial cell line-derived neurotrophic factor (GDNF), and vascular endothelial growth factor (VEGF).43–46 These factors collectively promote neuronal survival, axonal growth, synaptic plasticity, and angiogenesis.43–45
BDNF, a key neurotrophic factor in the central nervous system, activates the PI3 K/Akt and MAPK/ERK pathways via TrkB receptor binding, thereby supporting neuronal survival, differentiation, and synaptic function (Figure 1).63–66 In AD, BDNF expression levels are significantly reduced, leading to neuronal dysfunction and synaptic loss. 67 MSC transplantation restores BDNF expression in the brain, enhances anti-apoptotic capacity, and improves synaptic protein expression and cognitive function (Figure 1).68,69 Similarly, NGF supports cholinergic neuron survival and function through TrkA receptor activation (Figure 1). 70 Given that cholinergic neuron loss is a major contributor to cognitive decline in AD, MSC-derived NGF helps restore cholinergic neuron number and function, leading to improved memory and learning in AD animal models. GDNF, acting via GFRα and Ret receptors, promotes the survival of multiple neuronal populations.71,72 MSC transplantation elevates GDNF levels in AD brains, supporting neuronal survival and functional recovery. 46 VEGF, in addition to promoting angiogenesis, exerts direct neuroprotective effects.73,74 In AD, cerebrovascular dysfunction and blood-brain barrier disruption exacerbate neuronal damage. 75 MSC-derived VEGF improves cerebral blood flow, promotes vascular repair, and protects neurons from ischemic and toxic insults (Figure 1).73–75 Collectively, these neurotrophic factors secreted by MSCs concurrently enhance neuronal survival, synaptic plasticity, angiogenesis, and microenvironmental homeostasis. This multi-faceted neurotrophic support represents a key mechanism by which MSCs contribute to neural repair and functional recovery in AD (Table 1).
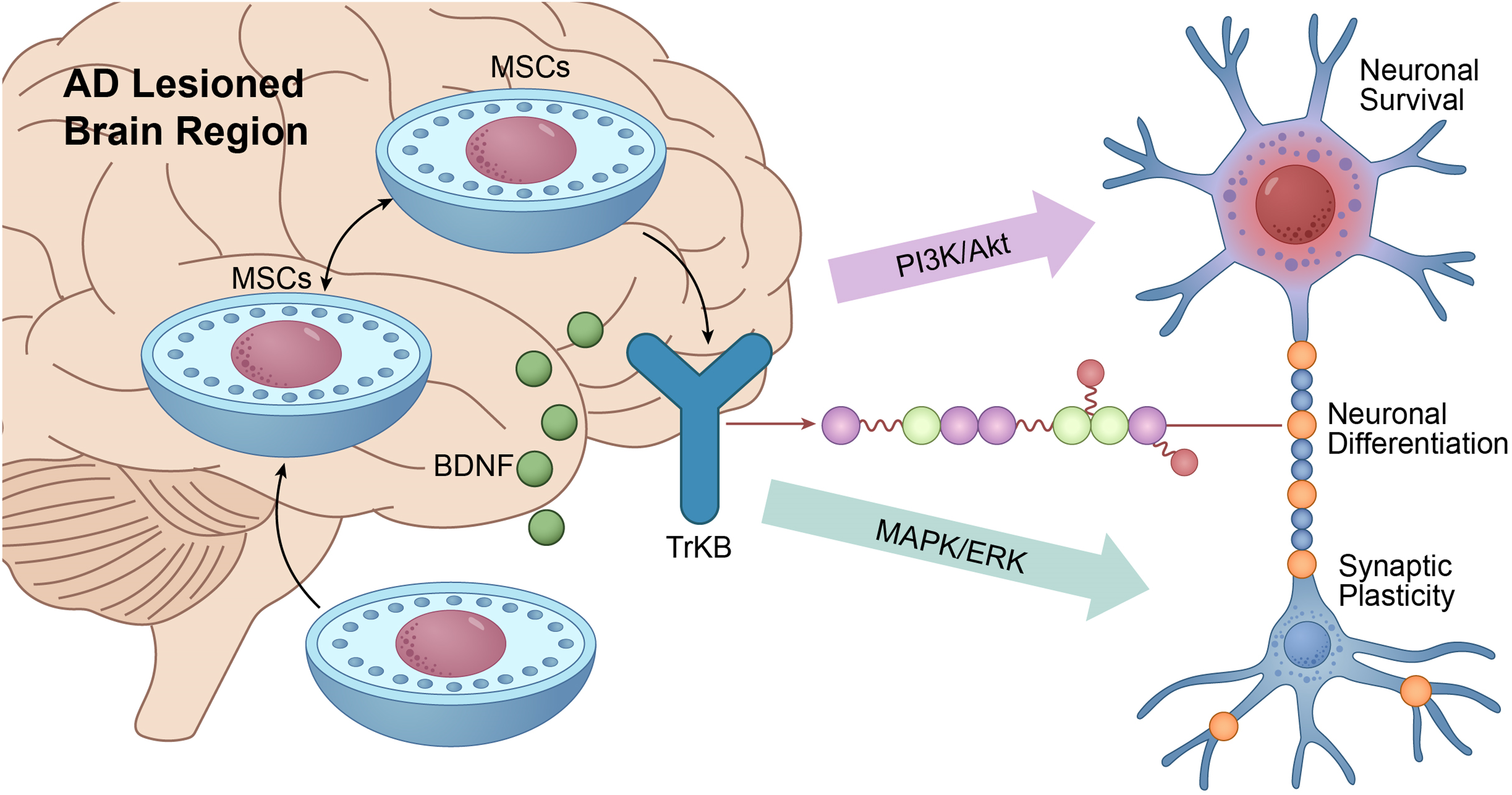
MSCs promote neuroprotection and regeneration via paracrine secretion of neurotrophic factors.
Anti-inflammatory and immunomodulatory effects of MSCs in AD treatment
Chronic neuroinflammation is a pivotal factor in the pathogenesis of AD. MSCs exert potent anti-inflammatory and immunomodulatory effects by suppressing microglial overactivation and regulating adaptive immune responses.
Inhibition of microglial over-activation
Microglia are the primary immune cells in the central nervous system. 22 In the pathological process of AD, Aβ deposition and neuronal damage activate microglia, triggering chronic neuroinflammation.13,76 Over-activated microglia release large amounts of pro-inflammatory factors, such as TNF-α, IL-1β, and IL-6, further exacerbating neuronal damage.77,78 MSCs effectively inhibit neuroinflammation by secreting anti-inflammatory factors and regulating microglial function. 79 MSCs can secrete various anti-inflammatory factors, such as IL-10 and transforming growth factor-β (TGF-β) (Figure 2).47,80 IL-10 is an important anti-inflammatory cytokine that can inhibit the production of pro-inflammatory factors and reduce inflammatory responses.47,81 TGF-β exerts anti-inflammatory effects by inhibiting microglial activation and pro-inflammatory factor release. 82 In AD animal models, after MSC transplantation, IL-10 and TGF-β levels in the brain were significantly increased, microglial activation was inhibited, and neuroinflammation was significantly reduced. 83 Microglia have two main phenotypes: M1 phenotype and M2 phenotype. 50 In AD, microglia mainly exhibit the M1 phenotype, releasing large amounts of pro-inflammatory factors, exacerbating neuroinflammation.48,49 MSCs can promote the transition of microglia from M1 to M2 phenotype, increasing the release of anti-inflammatory factors and reducing the production of pro-inflammatory factors. 47 For example, MSCs regulate microglial phenotype and alleviate neuroinflammation by secreting prostaglandin E2 (PGE2) and indoleamine 2,3-dioxygenase (IDO) (Figure 2).84,85 In summary, MSCs have immunomodulatory functions, can affect the local state of microglia, and have potential clinical application value in treating AD and other neurological conditions (Table 1).
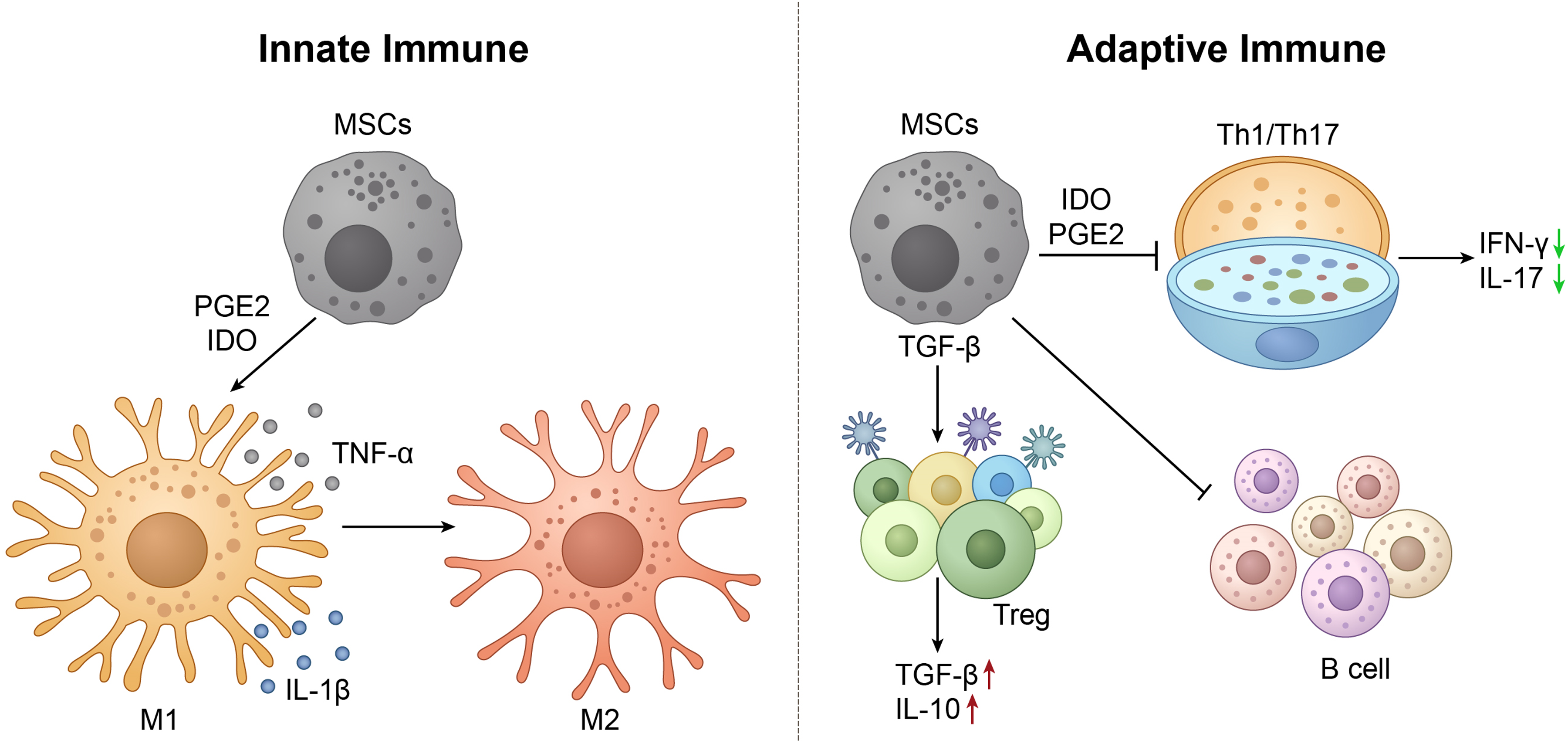
MSCs suppress neuroinflammation in AD by modulating innate and adaptive immune responses.
Modulation of T cell and B cell responses
Peripheral immune cells also play an important role in neuroinflammation in AD. In patients with AD, pro-inflammatory Th1 and Th17 cells responses are enhanced, and the function of regulatory T cells (Treg) is suppressed (Figure 2).51–53 Th1 and Th17 cells are two important pro-inflammatory T cell subsets that exacerbate neuroinflammation by secreting pro-inflammatory factors such as interferon-γ (IFN-γ) and IL-17.86,87 MSCs can inhibit the differentiation and function of Th1 and Th17 cells, reducing the release of pro-inflammatory factors. 88 For example, MSCs inhibit the differentiation of Th1 and Th17 cells and reduce IFN-γ and IL-17 levels by secreting IDO and PGE2, thereby alleviating neuroinflammation (Figure 2).85,88,89 Tregs are an important anti-inflammatory T cell subset that inhibits pro-inflammatory immune responses by secreting anti-inflammatory factors such as IL-10 and TGF-β (Figure 2).90,91 In AD, the number and function of Tregs are suppressed, leading to immune dysregulation. 92 For example, MSCs promote Treg cell differentiation and increase IL-10 and TGF-β release by secreting TGF-β and IL-2, thereby inhibiting neuroinflammation.93,94 In addition, B cells also play an important role in neuroinflammation in AD, exacerbating neuroinflammation by producing autoantibodies and pro-inflammatory factors.95,96 MSCs can inhibit B cell over-activation and autoantibody production, alleviating neuroinflammation (Figure 2). 53
In conclusion, MSCs achieve comprehensive regulation of the neuroimmune environment by simultaneously inhibiting pro-inflammatory cells and promoting anti-inflammatory cells (Table 1).
Role of MSCs in clearing Aβ deposition in AD treatment
The role of MSCs in AD treatment is not only reflected in neuroprotection and regeneration, as well as anti-inflammation and immunomodulation, but also in their ability to clear Aβ deposition. Abnormal Aβ deposition is a core pathological feature of AD. The oligomers and plaques formed are neurotoxic, leading to neuronal dysfunction and death. MSCs promote Aβ clearance directly or indirectly by enhancing Aβ phagocytosis and secreting degrading enzymes.
Enhancement of Aβ phagocytosis
In AD, the phagocytic function of microglia is impaired, leading to decreased Aβ clearance efficiency and increased Aβ deposition. 54 MSCs secrete cytokines such as colony-stimulating factor-1 to activate microglia and enhance their phagocytic function (Figure 3).55–57 In animal models of AD, after MSC transplantation, microglial phagocytic activity was significantly enhanced, and the number and size of Aβ plaques were significantly reduced (Figure 3). 97 In AD, microglia mainly exhibit the M1 phenotype, releasing large amounts of pro-inflammatory factors and exacerbating neuroinflammation, while their phagocytic function is inhibited.48,98 MSCs promote the transition of microglia from M1 to M2 phenotype, increasing their ability to phagocytose Aβ. In summary, MSCs weaken the pathological progression of AD by regulating microglial function, enhancing their ability to phagocytose Aβ, and regulating their phenotype (Table 1).

Dual pathways through which MSCs directly and indirectly promote Aβ clearance.
Secretion of degrading enzymes
In addition to enhancing microglial phagocytosis, MSCs secrete various degrading enzymes, such as insulin-degrading enzyme (IDE) and neprilysin (NEP). These degrading enzymes reduce the formation of Aβ oligomers and plaques by hydrolyzing Aβ, thereby alleviating Aβ-mediated neurotoxicity. IDE is a zinc metalloproteinase that can degrade various substrates, including Aβ, insulin, and glucagon.99,100 NEP is a membrane-bound metalloproteinase that can degrade various neuropeptides, including Aβ and enkephalin.58,59 In AD, the expression levels of IDE and NEP in the brain are reduced, leading to decreased Aβ degradation and increased Aβ deposition (Figure 3). 99 In AD animal models, after MSC transplantation, IDE and NEP levels in the brain were significantly increased, and the number and size of Aβ plaques were significantly reduced (Figure 3).60,61 Furthermore, MSCs upregulate the expression of IDE and NEP in host neurons and glial cells through paracrine effects, further enhancing Aβ degradation (Table 1). 62
Clinical evidence and combined therapeutic strategies of MSCs in AD treatment
In recent years, MSC-based therapies have achieved remarkable progress in the treatment of AD. A growing number of clinical trials have been initiated to systematically evaluate their safety, tolerability, and preliminary efficacy.101,102 To date, completed clinical investigations have primarily focused on Phase I/II trials. These studies often utilize randomized controlled or open-label designs and have mainly enrolled patients with mild to moderate AD.103,104 Notably, this review aligns with recent initiatives advocating for a holistic understanding of AD's pathological mechanisms. 41 It reflects a paradigm shift from single-target interventions toward multi-system regulatory therapies. 105
Regarding cellular sources, clinical trials have utilized MSCs derived from various tissues. These include umbilical cord-derived MSCs (UC-MSCs), bone marrow-derived MSCs (BM-MSCs), which are readily accessible and well-characterized biologically, as well as adipose tissue-derived MSCs (AD-MSCs) known for their abundant supply and strong proliferative capacity.106,107 Administration routes have become increasingly diversified. These include intravenous infusion for its operational simplicity, intrathecal injection to achieve higher central nervous system cell concentrations, and emerging approaches such as intranasal delivery, which bypasses the blood-brain barrier.108–110 Dosing regimens vary considerably across different studies. Common protocols involve single or multiple infusions, with cell doses typically ranging from 1 × 107 to 1 × 108 cells per administration.111–113 Current evidence suggests that intervention during the early stages of AD may be more advantageous for leveraging the neurorestorative and anti-inflammatory effects of MSCs.102,114,115 Regarding safety, MSC transplantation is generally well-tolerated. 116 Most studies have not reported treatment-related serious adverse events. Common adverse events include transient fever, headache, and nausea. These reactions are generally mild and typically resolve spontaneously within a short period.113,116 Preliminary efficacy results indicate improved cognitive function scores in some patients. Additional findings include reduced Aβ burden and attenuated neuroinflammation. These observations provide initial support for the potential benefits of MSC-based therapy.103,104,115,117
The therapeutic value of MSCs extends beyond cell replacement. It lies primarily in their potent paracrine and immunomodulatory functions. For example, MSCs significantly alleviate neuroinflammation by modulating microglial polarization and promoting regulatory T-cell proliferation. 104 When combined with anti-Aβ antibodies, this approach simultaneously addresses both the inflammatory microenvironment and amyloid plaque deposition.118–120 Furthermore, MSCs secrete neurotrophic factors, such as BDNF and GDNF. 121 In combination with anti-tau therapeutics, these factors may further enhance neuronal survival and synaptic function. Beyond directly targeting Aβ and tau, immunomodulatory therapies have emerged as a promising new direction in AD treatment. The NLRP3 inflammasome has been identified as a critical driver of neuroinflammation in AD. Specifically, the inhibitor MCC950 has been shown to improve cognitive function in mouse models by attenuating NLRP3-mediated inflammation.120,122 Furthermore, emerging evidence suggests that MCC950's benefits may extend beyond simple inhibition, potentially involving the modulation of autophagic pathways to clear pathological proteins. 123 This dual mechanism of action presents a promising novel therapeutic perspective for AD.
Notwithstanding these promising developments, the field continues to face several challenges. These include inconsistencies across study outcomes, incomplete elucidation of mechanistic actions, uncertainties regarding long-term efficacy sustainability, and hurdles in standardized manufacturing protocols. Consequently, to address these challenges, future progress requires the implementation of larger-scale randomized controlled trials that integrate multi-omics technologies and mechanistic investigations, to further advance the clinical translation of MSC-based therapeutics (Table 2).
From bench to bedside: overview of translational research on MSC therapy for AD.

Conclusion
MSCs have demonstrated considerable potential in the treatment of AD through their multifaceted mechanisms of action, which include neuroprotection and regeneration, anti-inflammation and immunomodulation, and Aβ clearance. Preclinical studies in various AD animal models have confirmed their efficacy in ameliorating pathological features and cognitive behavior. Preliminary clinical trials have also verified their safety and potential therapeutic benefits. However, the clinical application of MSCs still faces challenges, such as the need to resolve issues related to in vivo survival rates, functional integration, standardization of cell sources, and dosage protocols. In the future, by optimizing delivery methods, improving cell survival rates, and combining MSCs with other treatment strategies, such as gene editing and drug combination therapy, MSCs are expected to become a key component of a comprehensive AD treatment framework, offering patients more effective therapeutic options. This multi-target approach aligns closely with the emerging holistic perspective on AD management, which advocates for integrated therapeutic strategies addressing the complex, multifactorial nature of the disease. 124
Footnotes
Acknowledgements
The authors have no acknowledgments to report.
Author contribution(s)
Funding
The author received no financial support for the research, authorship, and/or publication of this article.
Declaration of conflicting interests
The author declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.