
Abstract
The role of endothelium-derived hyperpolarizing factor (EDHF) in either the healthy circulation or in those with hypercholesterolemia is unknown. In healthy and hypercholesterolemic subjects, we measured forearm blood flow (FBF) using strain-gauge plethysmography at rest, during graded handgrip exercise, and after sodium nitroprusside infusion. Measurements were repeated after
Keywords
Introduction
The endothelium plays a dynamic role in regulating vasomotor tone and blood flow in response to physiologic and chemical signals by releasing paracrine vasoactive factors that include nitric oxide (NO), prostacyclin, and endo-thelium-derived hyperpolarizing factors (EDHF). Even after inhibition of NO and cyclooxygenase, endothelium-dependent vasodilation persists, revealing the existence of a substantial NO- and prostaglandin-independent component that has been attributed to EDHF release.1–4 The hallmark of EDHF-mediated responses is their abolition by the combination of apamin (a specific inhibitor of K+Ca channels of small conductance vessels) plus charybdotoxin (a non-selective inhibitor of large-conductance and intermediate-conductance channels, as well as some voltage-dependent K+ channels), as noted in pre-clinical studies.5,6 Although potential EDHFs may differ by species and vascular bed,6–10 they are thought to promote vasodilation via a common pathway of activation of calcium-dependent potassium (K+Ca) channels.11,12 Recently, we demonstrated using tetraethylammonium chloride (TEA) – a specific K+Ca channel antagonist that EDHF contributes to both resting vasodilator tone and to bradykinin-stimulated vasodilation.13,14
Microvascular forearm vasodilation during exercise is a multi-factorial phenomenon that includes contribution from local muscle metabolites such as lactate, adenine nucleotides, acidosis, decreased tissue oxygen tension, increased carbon dioxide tension, and osmolality. 15 The magnitude of the contribution of NO and prostacyclin to exercise-induced vasodilation appears to be modest; inhibition of NO and prostacyclin causes an 11% and 13% reduction in exercise-induced vasodilation, respectively.2,16–22 The contribution of EDHF to exercise-induced forearm microvascular vasodilation in humans remains unknown, with a variable contribution observed in previous studies in leg muscles.16,17 Importantly, the impact of cardiovascular risk factors that are known to impair NO activity on the contribution of EDHF to vasodilation during exercise remains un-known.23–27 We investigated the contribution of NO and EDHF to exercise-induced forearm vasodilation in healthy humans and in those with hypercholesterolemia. We hypothesized that endothelium-dependent hyperpolarization contributes to exercise-mediated forearm microvascular vasodilation via activation of K+Ca channels, and that the contribution of NO and EDHF varies between the healthy and hypercholesterolemic vasculature.
Methods
Subjects
Twenty-six healthy individuals and 19 subjects with hypercholesterolemia aged between 21 and 60 years (47% male) were enrolled into two separate protocols (Figure 1). All subjects were non-smokers and free from hypertension, cardiovascular disease, systemic disorders, and any oral medications. Pregnant females were excluded. Hyper-cholesterolemia was defined as a low-density lipoprotein (LDL)-cholesterol level of >140 mg/dL. The study was approved by the Emory University Institutional Review Board and all subjects provided informed consent.
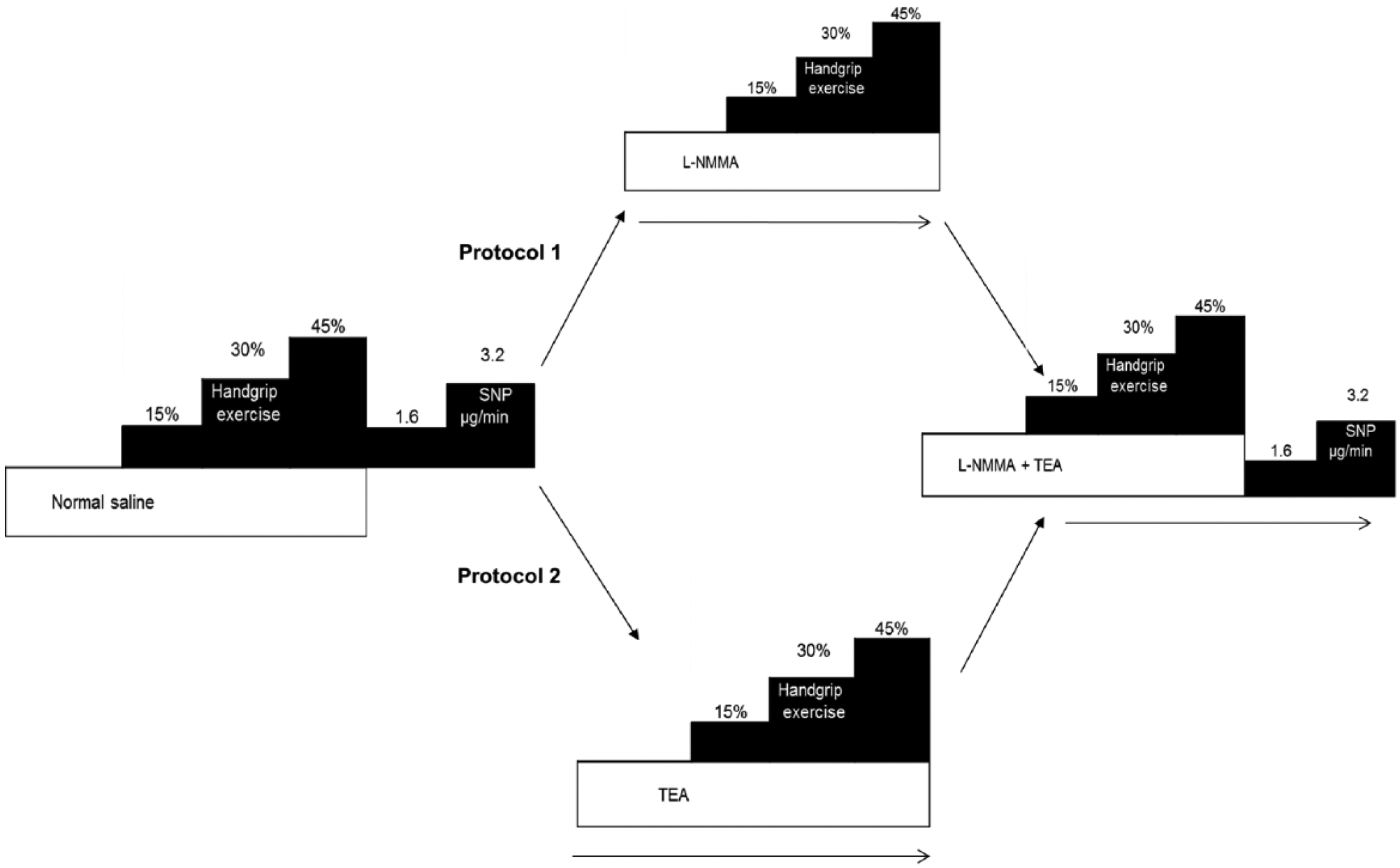
Study design. Aspirin (975 mg) was administered 1 hour prior to commencement of the study for cyclooxygenase inhibition. Forearm blood flow measurements were performed after handgrip exercise (15%, 30% and 45% of maximum grip strength) and after endothelium-independent vasodilation with intra-arterial sodium nitroprusside (SNP; 1.6, 3.2 µg/min). Measurements were repeated after
Measurement of forearm blood flow at rest and during exercise
Measurements were performed after an overnight fast in a quiet temperature-controlled (22°C to 24°C) room. Subjects refrained from exercise, alcohol and caffeine for at least 24 hours previously. After insertion of an intra-brachial arterial cannula for arterial pressure monitoring and delivery of drug infusions, subjects received oral aspirin (975 mg) to inhibit prostacyclin synthesis at least 1 hour prior to the study. 28 Simultaneous forearm blood flow (FBF) measurements were obtained in both arms using a dual-channel venous occlusion strain-gauge plethysmograph (model EC6; DE Hokanson, Bellevue, WA, USA) as described previously. 19 Flow measurements were recorded for approximately 7 seconds, every 15 seconds up to eight times, and a mean FBF value in mL·min−1·100 mL−1 was computed. Forearm vascular resistance (FVR) was calculated as the mean arterial pressure ÷ FBF and expressed as mmHg per mL·min−1·100 mL−1.
The instrumented forearm was exercised with intermittent handgrip by squeezing an inflated pneumatic bag, as previously described.2,29 Exercise was performed at 15%, 30%, and 45% of maximum voluntary grip strength, which was determined in each subject. Each contraction lasted for 5 seconds followed by relaxation for 10 seconds. The contraction–relaxation sequence was repeated for a total of 5 minutes at each workload and FBF measurements were made in the final 2 minutes at each workload. Preliminary studies and previous studies in our laboratory demonstrated that FBF remained constant after 2 minutes of exercise. 2
Contribution of K+Ca channel activation and NO to exercise-induced vasodilator tone
Protocol 1
All agents were administered intra-arterially after resting measurements were made during saline infusion (2.5 mL·min−1). To investigate the effect of inhibition of K+Ca channels without prior inhibition of NO on exercise-induced vasodilation, we studied 10 healthy and nine hypercholesterolemic subjects. FBF was measured during infusion of saline (1 mL·min−1), after exercise, and after intra-arterial infusions of the endothelium-independent vasodilator sodium nitroprusside at 1.6 and 3.2 mg/min for 8 minutes each (Figure 1). After repeating resting flow measurements, FBF was measured during an 8-minute infusion of intra-arterial tetraethylammonium chloride (TEA; Clinalfa, Switzerland) at 1 mg/min. While continuing the infusion of TEA, exercise was repeated as before. After a 30-minute rest period, FBF measurements were repeated during an 8-minute infusion of
Arterial blood pressure and FBF measurements were repeated in the last 2 minutes of each intervention (Figure 1). This protocol enabled measurements of the contribution of NO and combined NO and EDHF blockade on exercise-induced vasodilation.
Protocol 2
To investigate the effect of inhibition of K+Ca channels after NO inhibition on exercise-induced vasodilation, protocol 1 was repeated in separate experiments in 16 healthy and 10 hypercholesterolemic subjects. During saline infusions, FBF was measured at rest, after graded handgrip exercise, and after infusions of sodium nitroprusside. Measurements were repeated after an 8-minute infusion of
Statistical analysis
FBF and FVR were log-transformed into a normal distribution. The effects of
Results
Baseline characteristics
Hypercholesterolemic subjects had higher total and LDL-cholesterol and triglyceride levels. They were also older, had a higher diastolic blood pressure and a greater BMI compared to healthy subjects (Table 1).
Characteristics of study subjects.
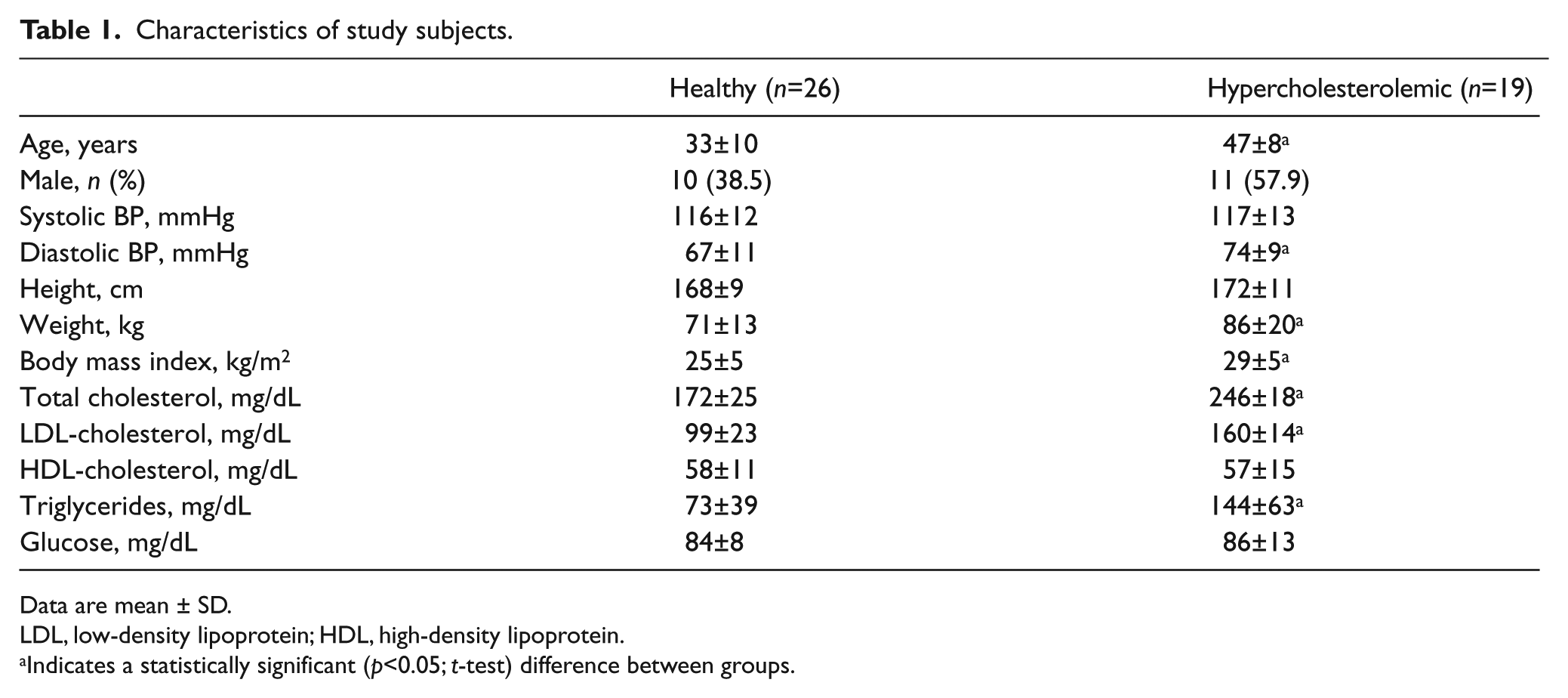
Data are mean ± SD.
LDL, low-density lipoprotein; HDL, high-density lipoprotein.
Indicates a statistically significant (p<0.05; t-test) difference between groups.
Effects of l -NMMA and TEA on resting vasodilator tone
There were no significant differences in baseline FBF and FVR between healthy and hypercholesterolemic subjects. FBF decreased and FVR increased significantly with
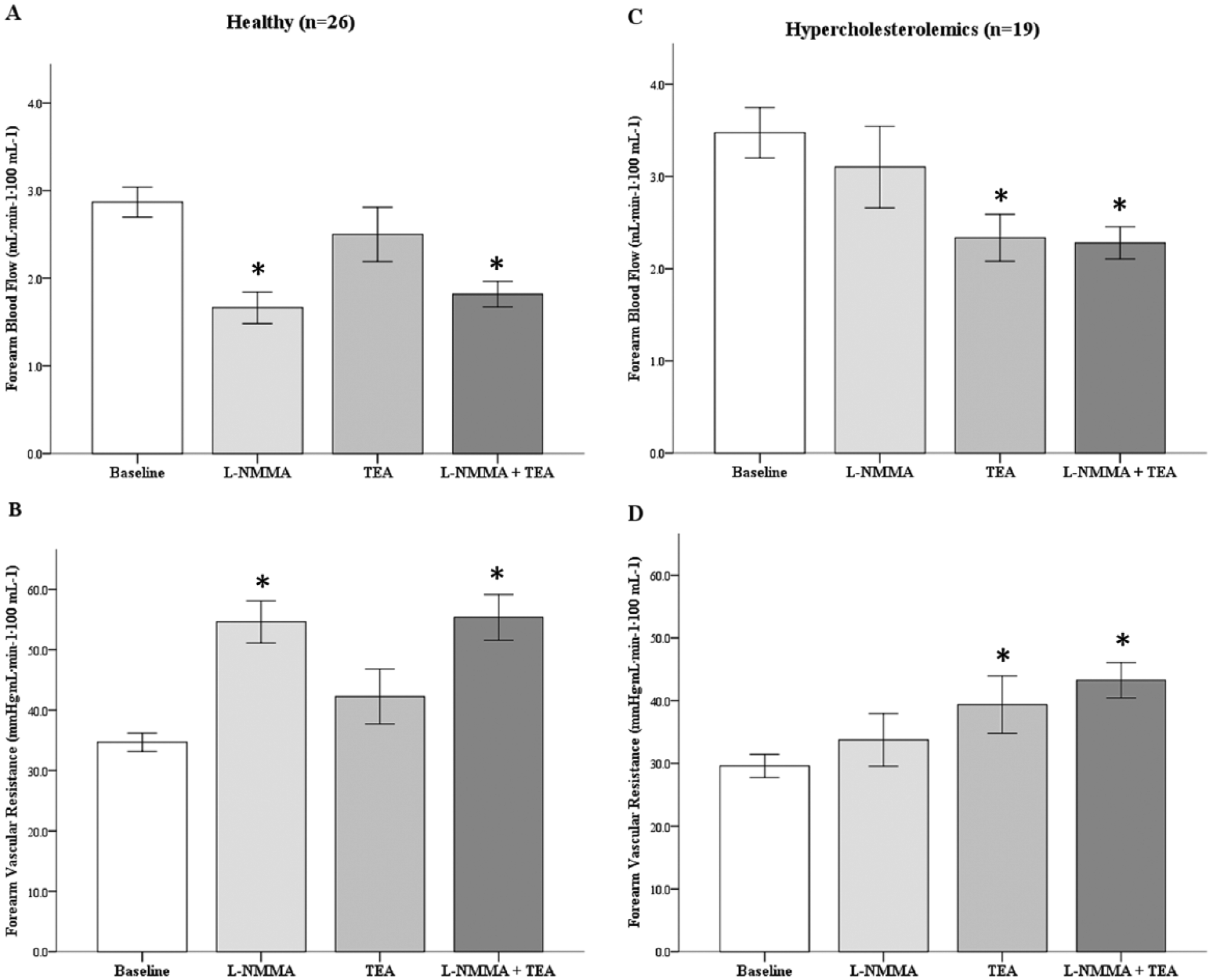
Contribution of NO and K+Ca channel activation to resting vascular tone in healthy and hypercholesterolemic subjects. Resting forearm blood flow (FBF) (A and C) and forearm vascular resistance (FVR) (B and D) in response to
Exercise-induced vasodilation in healthy and hypercholesterolemic subjects
Handgrip exercise produced increasing vasodilation that was similar at the lower two levels of exercise (15% and 30% of maximal handgrip) in both groups, but was lower at peak exercise (45% of maximal handgrip) in hypercholesterolemic compared to healthy subjects (change in FBF: +274% vs +438%, p=0.017; FVR: –69% vs –78%, p=0.004, respectively) (Figure 3). Throughout the experiments, the handgrip exercise did not cause significant change in mean arterial blood pressure, heart rate, or blood flow in the contralateral control arm.
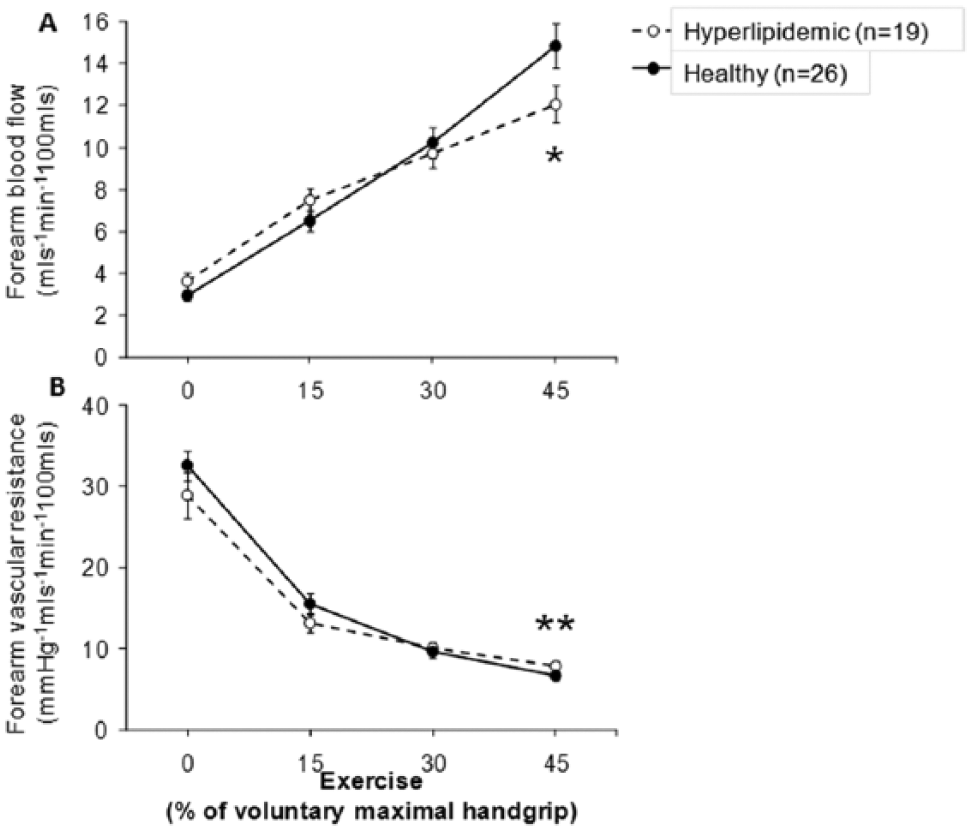
Forearm vasodilator response to exercise-induced vasodilation in healthy and hypercholesterolemic subjects. (A) Forearm blood flow (FBF) and (B) forearm vascular resistance (FVR) responses to handgrip exercise in 26 healthy and 19 hyperlipidemic subjects. Data shown as mean ± SEM. *p<0.05, **p<0.005.
Effect of TEA alone and in combination with l -NMMA on forearm vascular tone during exercise
Healthy subjects (n=10)
Exercise-induced vasodilation was attenuated by TEA in the absence of
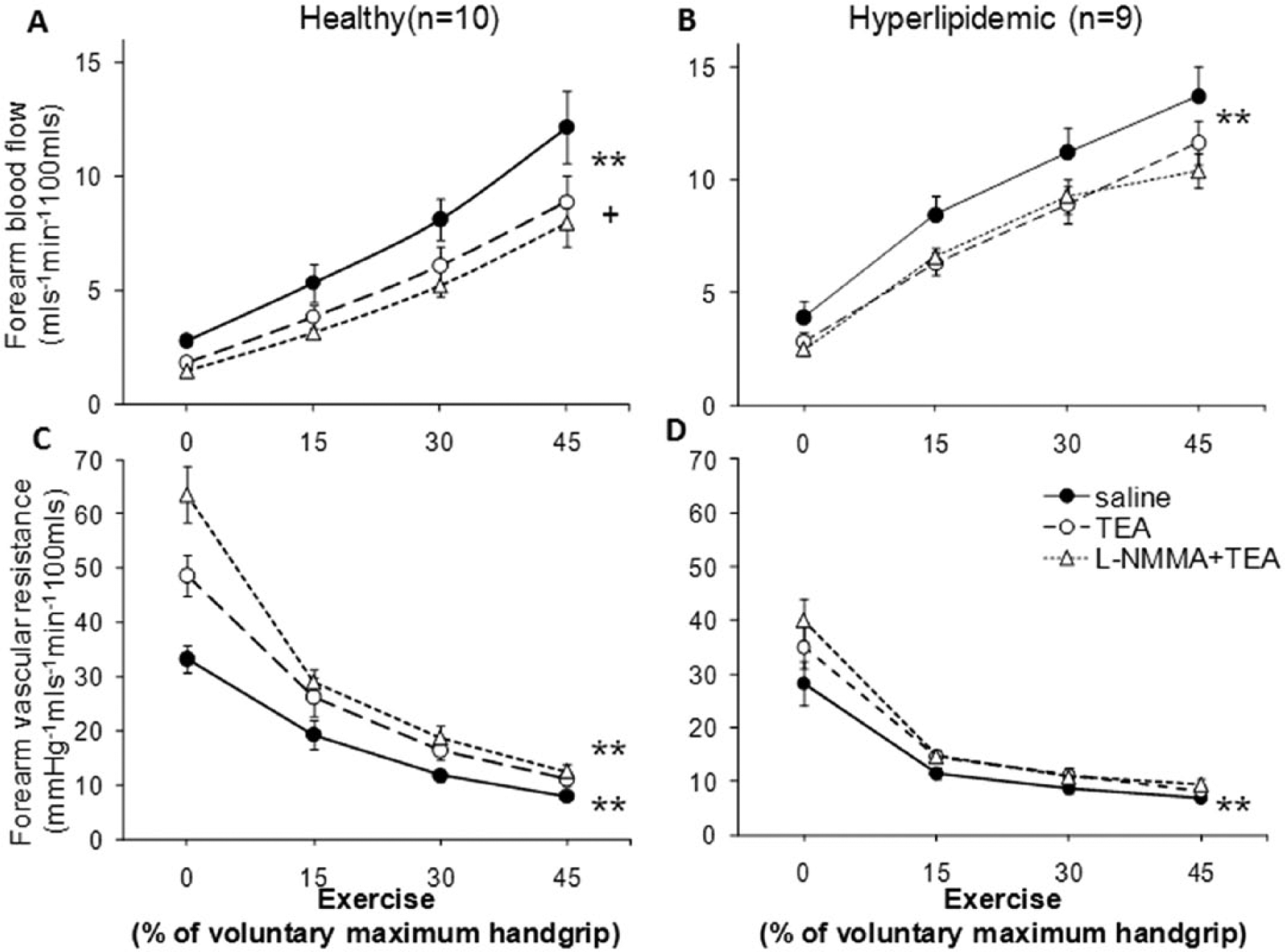
Contribution of K+Ca channel activation, in the presence and absence of nitric oxide, to exercise-induced forearm vasodilation in healthy and hypercholesterolemic subjects. Forearm blood flow (FBF) and forearm vascular resistance (FVR) changes in response to increasing handgrip exercise before and after TEA (1 µmol/min) and combined administration of TEA and
Hypercholesterolemic subjects (n=9)
Infusion of TEA alone attenuated exercise-induced vasodilation in hypercholesterolemic subjects; mean FBF was reduced by 20.3% (p<0.0001) and mean FVR was increased by 24.9% (p<0.0001) (Figure 4B, 4D). However, co-infusion of
Effect of l -NMMA alone and in combination with TEA on forearm vasodilation during exercise
Healthy subjects (n=16)
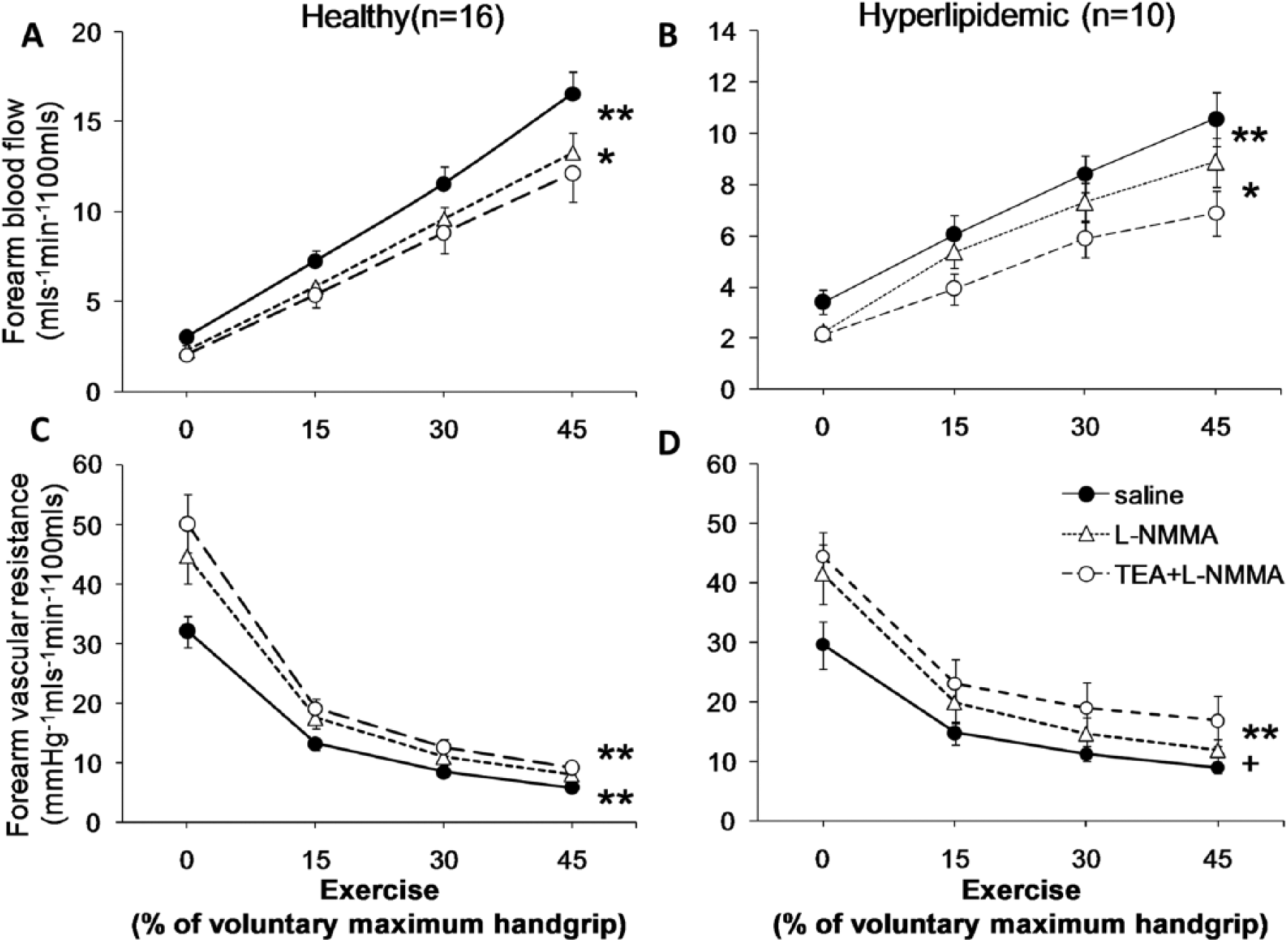
Contribution of nitric oxide, in the presence and absence of K+Ca channel activation, to exercise-induced forearm vasodilation in healthy and hypercholesterolemic subjects. Forearm blood flow (FBF) and forearm vascular resistance (FVR) changes in response to handgrip exercise before and after
Hypercholesterolemic subjects (n=10)
Vasodilator responses to sodium nitroprusside
Infusion of sodium nitroprusside produced a stepwise vasodilator response that was similar (FBF, p=0.37; FVR, p=0.70) in healthy and hypercholesterolemic subjects. Combined infusions of
Discussion
Herein we demonstrate that both NO and K+Ca channel activation (EDHF) contribute equally and in concert to exercise-induced microvascular vasodilation in the healthy human forearm circulation. Dual inhibition of these endothelium-derived factors caused further inhibition of microcirculatory vasodilation during exercise, demonstrating the independent and additive contribution of NO and EDHF to physiologic vasodilation. In contrast, in hypercholesterolemia, K+Ca channel activation appeared to be the predominant mechanism for exercise-induced vasodilation; after K+Ca channel blockade, inhibition of NO had no further effect on the vasodilator response to exercise in these subjects. Finally, the lack of inhibition of sodium nitroprusside-mediated endothelium-independent vasodilation by
NO and exercise-induced vasodilation
The mechanisms underlying exercise-induced vasodilation are complex and the understanding of the potential mediators remains incomplete. Skeletal muscle blood flow increases in proportion to the metabolic demand.33–37 The precise contribution of the various vasoactive compounds to exercise-induced vasorelaxation may also differ by muscle beds studied and involve the interaction of the autonomic nervous system.37–40 Prostacyclins contribute modestly and transiently to exercise-induced vasodilation
20
and, for the purposes of this study, cyclooxygenase was inhibited prior to the study. Previous studies have either shown modest inhibition of exercise hyperemia with NO blockade2,17,20 or have failed to demonstrate any contribution of NO to exercise,21,41,42 raising the concept of redundancy, where compensatory release of other vasodilators may explain the lack of effect of NO inhibition.
16
Differences in experimental methods, sample size, and muscle beds examined may also account for the differences.21,41,42 Herein, we have demonstrated a 19.3% reduction in mean vasodilation in response to exercise in healthy individuals after inhibition of NO, consistent with, or more pronounced than the 11% reduction observed previously.
2
This difference may be due to the comparatively younger age and consequently greater NO activity in the current healthy cohort. There was a contribution of NO to exercise-induced vasodilation even after K+Ca channel blockade, with a further 13.6% reduction in flow with
EDHF and exercise-induced vasodilation in healthy subjects
In a previous study of seven healthy subjects, inhibition of cytochrome P450 2C9 with sulphaphenazole did not alter exercise-induced vasodilation in thigh muscles, suggesting lack of contribution of epoxyeicosatrienoic acids to exercise hyperemia. 16 Our results provide the first evidence for the contribution of EDHF, and in particular of K+Ca channel activation, to physiological vasodilation of the forearm microvasculature during exercise. We have demonstrated a 27.3% reduction in exercise-induced vasodilation with TEA, even in the absence of inhibition of NO, and an additional reduction in FBF with TEA after NO blockade. Taken together with the findings of the previous study, it would appear that EDHFs stimulated by exercise that lead to K+Ca channel stimulation may not be cytochrome P450-derived peptides, and potentially involve hydrogen peroxide and other EDHF mechanisms. In contrast to our findings, Mortensen et al. failed to demonstrate an effect of K+Ca channel blockade with TEA when measuring blood flow changes in response to leg exercise. 17 Variations in muscle beds studied and techniques (thermodilution versus plethysmography) may have accounted for the observed differences. The intravascular concentration of TEA (0.5 mmol/L) used was similar in this study, and has previously been demonstrated to specifically inhibit K+Ca channels.30,43–46
The contribution of both NO and K+Ca channel activation to exercise hyperemia was similar in magnitude in the healthy subjects. This was unexpected based on experimental studies, demonstrating that the contribution of EDHF is only evidenced after NO blockade.16,46–48 Even after inhibition of NO with
EDHF and exercise-induced vasodilation in hypercholesterolemia
In hypercholesterolemic subjects, the attenuation of exercise-induced vasodilation after NO blockade, when EDHF was not inhibited, was similar to the responses observed in healthy subjects, suggesting that NO contributes to physiologic vasodilation. This is consistent with our previous observations of preserved endothelium-dependent vasodilation with bradykinin in hypercholesterolemia. 13 However, in contrast to healthy subjects and the effects on bradykinin responses, after EDHF blockade with TEA, no further attenuation of exercise-induced vasodilation was observed with NO blockade in subjects with hypercholesterolemia. This suggests that in vasculature exposed to hypercholesterolemia, K+Ca channel activity may be the predominant mechanism for exercise-induced vasodilation. It also suggests that NO and K+Ca channels may interact such that inhibition of K+Ca channels may inhibit the NO pathway in hypercholesterolemia, an observation that needs further investigation.
We have further demonstrated that hypercholesterolemic individuals have an attenuated forearm vasodilator response to exercise at the highest handgrip effort when compared to healthy individuals. Also, in contrast to the healthy microvasculature, there is a lack of contribution of NO to exercise-induced vasodilation after K+Ca channel blockade, and this NO deficiency may contribute to the lower vasodilator response in the hypercholesterolemic subset. This reduction of NO activity was as a result of decreased NO production or availability and not from vascular smooth muscle dysfunction, as the sodium nitroprusside responses were similar in both groups.
It may be argued that the reduced vasodilation observed during exercise with
Limitations
Our findings are limited to the forearm microcirculation and thus other vascular beds, including conductance arteries, warrant further investigation. Nevertheless, it is known that the contribution of EDHF is less in con-ductance vessels than in the microvessels.8,12,49 Second,
Conclusions
We have demonstrated an important contribution of EDHF, via the activation of K+Ca channels, to forearm microvascular vasodilation during exercise in healthy subjects with a greater contribution compared to NO in hypercholesterolemic subjects. Whether enhancing EDHF in conditions with impaired NO activity would be of therapeutic value, and whether agents that improve endothelial dysfunction, such as statins and angiotensin antagonists, also enhance EDHF bioactivity remains to be studied.
Footnotes
Declaration of conflicting interest
The authors declare that there is no conflict of interest.
Funding
National Institutes of Health Research Grant RO1 HL79115, PHS Grant UL1 RR025008 from the Clinical and Translational Science Award Program, and PHS Grant M01 RR00039 from the General Clinical Research Center Program, National Institutes of Health, National Center for Research Resources, British Cardiovascular Society Research Fellowship, and the National Blood Foundation.