
Abstract
Atherosclerotic cardiovascular disease has been acknowledged as a chronic inflammatory condition. Monocytes and macrophages lead the inflammatory pathology of atherosclerosis whereas changes in atheromatous plaque thickness and matrix composition are attributed to vascular smooth muscle cells. Because these cell types are key players in atherosclerosis progression, it is crucial to utilize a reliable system to investigate their interaction. In vitro co-culture systems are useful platforms to study specific molecular mechanisms between cells. This review aims to summarize the various co-culture models that have been developed to investigate vascular smooth muscle cell and monocyte/macrophage interactions, focusing on the monocyte/macrophage effects on vascular smooth muscle cell function.
Introduction
Vascular smooth muscle cells (VSMCs) and monocytes/macrophages (Mo/MΦ) have distinct roles yet intimate interaction in the inflammatory pathology of atherosclerotic cardiovascular disease. It is thus essential to examine the cross-talk and molecular pathways involved in their exchanges. Several models have incorporated murine cells to ascertain a better understanding of the role of MΦ and VSMC in vascular disease formation; 1 however, there is only about a 50% overlap between human and mice MΦ at the transcriptional level, 2 creating a limitation in the utility of murine models to study human macrophage impact. While there are several benefits to using in vivo systems, in vitro co-culture systems are also useful platforms to study specific interaction mechanisms without the inherent variables associated with in vivo models. This review will focus on the impact of Mo/MΦ on VSMC function with the following objectives: (1) summarize the related atherosclerotic biology of these cells; (2) evaluate the various co-culture models that have been developed; (3) address the advantages, limitations, and key findings of each model; and (4) describe the implications of co-culture for clinical applications and potential future directions.
Pathophysiology of atherosclerosis
Atherosclerotic cardiovascular disease is the leading cause of death in developed countries. Its onset is believed to be precipitated by endothelial cell (EC) dysfunction and the recruitment of inflammatory compounds. Dysfunction of the endothelium leads to the subsequent migration of circulating immune cells to the sub-endothelial space. A variety of immune cells (monocytes, macrophages, platelets and T-cells) have been identified as participants in the disease progression; however, Mo/MΦ are considered the primary players regulating atherosclerotic inflammation. Although the majority of MΦ are believed to originate from the circulating Mo that have transmigrated to the sub-endothelial space,3,4 several mouse-based studies have demonstrated MΦ proliferative capacity in plaques, suggesting the role of tissue-resident macrophages.5,6 A recent investigation using apolipoprotein E (ApoE)-knockout mice, an atherosclerotic animal model, revealed that the majority of plaque MΦ originated from tissue-resident MΦ. 7 However, a correlation to human atherosclerotic plaque composition remains to be elucidated.
Atherosclerotic plaque progression also involves the differentiation, migration and proliferation of VSMCs. Stimulated by a dysfunctional endothelium and inflammatory triggers, VSMCs are believed to differentiate and migrate from the tunica media to the intima where they proliferate and induce the formation of a fibrous cap, a structure that has been linked to plaque stability. VSMC apoptosis, on the other hand, can contribute to plaque vulnerability and rupture,8,9 which elicits luminal thrombosis, distal embolization, and terminal clinical events, such as myocardial infarction and stroke. 10
Role of macrophages in atherosclerosis
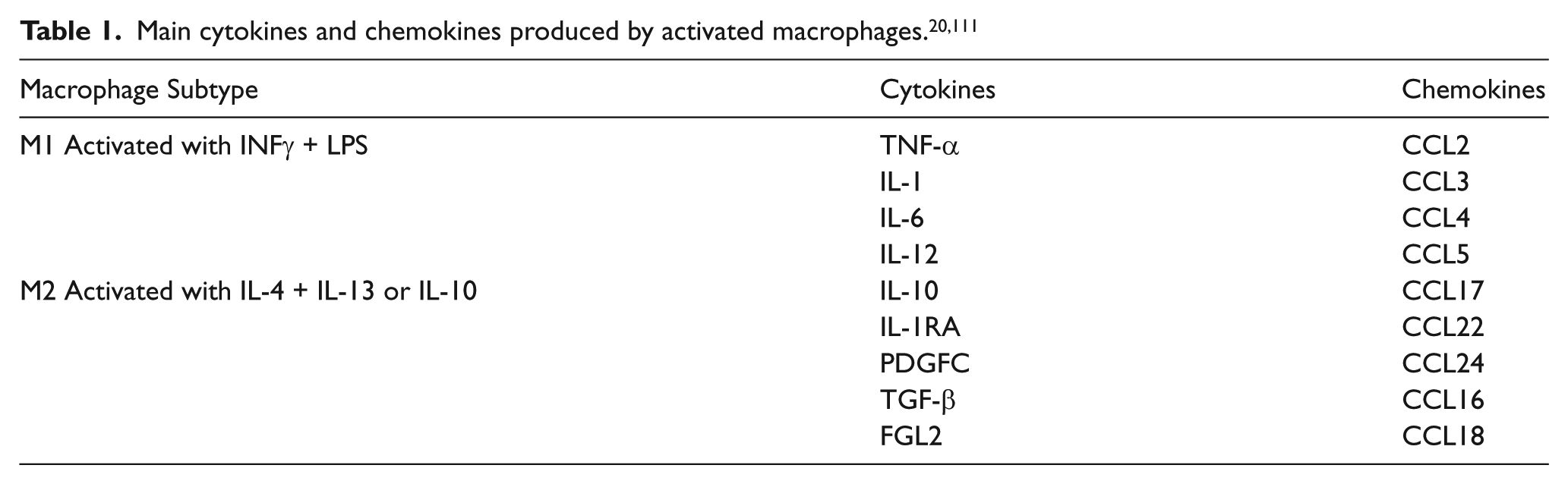
In 1958, Poole and Florey were the first to describe the presence of MΦ beneath the endothelial layer of vessel walls using experimental rabbit atheroma. 11 Thereafter, several other research groups also documented a number of MΦ within unstable, human atherosclerotic walls.12–17 Within the vessel wall, the local microenvironment drives MΦ differentiation and polarization. Macrophage colony-stimulating factor (M-CSF) and granulocyte-macrophage colony-stimulating factor are two growth factors necessary for Mo differentiation towards MΦ;18,19 however, the atherosclerotic microenvironment is complex as it contains a multitude of factors, cytokines, and local byproducts. In the last decade, researchers have noted the ability of MΦ to differentiate into sub-phenotypes when exposed to different external stimuli, demonstrating the heterogeneity and plasticity of MΦ. The two main MΦ subtypes widely accepted are: pro-inflammatory macrophages (M1) and anti-inflammatory macrophages (M2). A microenvironment containing the cytokines interleukin 4 (IL-4) and interleukin 13 (IL-13) or interleukin 10 (IL-10) promotes M2 polarization, whereas interferon-γ (INF-γ) and lipopolysaccharides (LPS) promote the M1 phenotype. 20 Based on their cytokine/chemokine secretions upon polarization (Table 1), M1 are believed to be involved in inflammation whereas M2 have been linked to tissue repair.
The plasticity of MΦ phenotype can also be affected by the presence of oxidized low-density lipoproteins (ox-LDL), which can accumulate in atherosclerotic vessels due to an increased LDL influx from the blood stream and oxidative stress in the vessel wall. Upon entrance to the intima, some Mo may undergo MΦ foam cell transformation via MΦ uptake and metabolism of ox-LDL through membrane scavenger receptors, which deliver LDL to lysosomes for breakdown into cholesterol and fatty acids. 21 MΦ that retain cholesterol in their cytoplasm develop a foam-like appearance and are lipid-laden—hence, ‘foam cells’—and are commonly found in early atherosclerotic lesions. 22 Like M1 and M2, these foam cells can also impact the functionality of VSMCs, and set a particular path of progression for atherosclerosis. MΦ exist in a range of both pro- and anti-inflammatory states depending on the area within the plaque as well as the vessel type. 23 Generally, M1 macrophages have been found to be the most populous subtype within inflamed plaque areas.
Role of VSMCs in atherosclerosis
The tunica media of healthy vessel walls is composed of VSMCs responsible for modulating vessel diameter in response to external stimuli. Displaying phenotypic plasticity, VSMCs are often noted for their ability to switch between a ‘contractile’ and ‘synthetic’ state in response to different environmental conditions. 24 In a normal vessel environment, VSMCs are contractile and responsive to stimuli generated by changes in blood flow and blood pressure; however, in an atherosclerotic environment, VSMCs undergo a transformation to a proliferative and migratory phenotype (synthetic).25,26 VSMCs can then migrate from the media to the intima via cytoskeletal and extracellular matrix (ECM) remodeling, as previously noted by several reviews on the molecular mechanisms involved in VSMC migration.27,28
An increased expression of matrix metalloproteinases (MMPs) MMP-1, MMP-2, MMP-3, and MMP-9 has been observed in atherosclerotic plaques, 29 which have been shown to play roles in ECM remodeling for VSMC migration.30–34 As atherosclerosis progresses, VSMCs continue to proliferate, altering the formation of atheroma, which is largely composed of VSMCs and their synthetic products (collagens, proteoglycans, and elastin), as well as inflammatory cells.35,36 As a result, VSMCs become a major cellular component of an atheroma through their migrative, proliferative, and matrix-producing activity.
Physical interaction between vascular and immune cells
VSMCs and ECs can both express intercellular adhesion molecules that stimulate Mo recruitment to the vessel wall. Specifically, intercellular adhesion molecule-1 (ICAM-1), vascular cell adhesion molecule-1 (VCAM-1), macrophage-chemoattractant protein 1 (MCP-1), and fractalkine (CX3CL1) have been shown to stimulate Mo attachment. 37 Furthermore, Mo binding to VSMCs through VCAM-1 and ICAM-1 can prevent Mo death, 38 whereas the attachment of Mo to VSMCs via CX3CL1 can increase the expression of proatherogenic molecules. 39 In addition, the interaction of VSMCs with Mo/MΦ has also been shown to enhance MMPs production, which can play pivotal roles in VSMC migration,39–41 alter VSMC phenotype and proliferative capacity,42–45 and mediate VSMC apoptosis via Fas receptor-ligand binding to MΦ.46,47 At the advanced atherosclerotic disease stages, functional impairment of both cell types is reflected by the development of a necrotic core in plaques; the necrotic core is composed of accumulated cholesterol, inflammatory cells, cytokines, and apoptotic VSMCs.9,14,48 In turn, this may result in an overwhelming plaque destabilization leading to thrombi formation and potential vessel rupture.
In vitro co-culture models to study VSMCs and Mo/MΦ in atherosclerosis
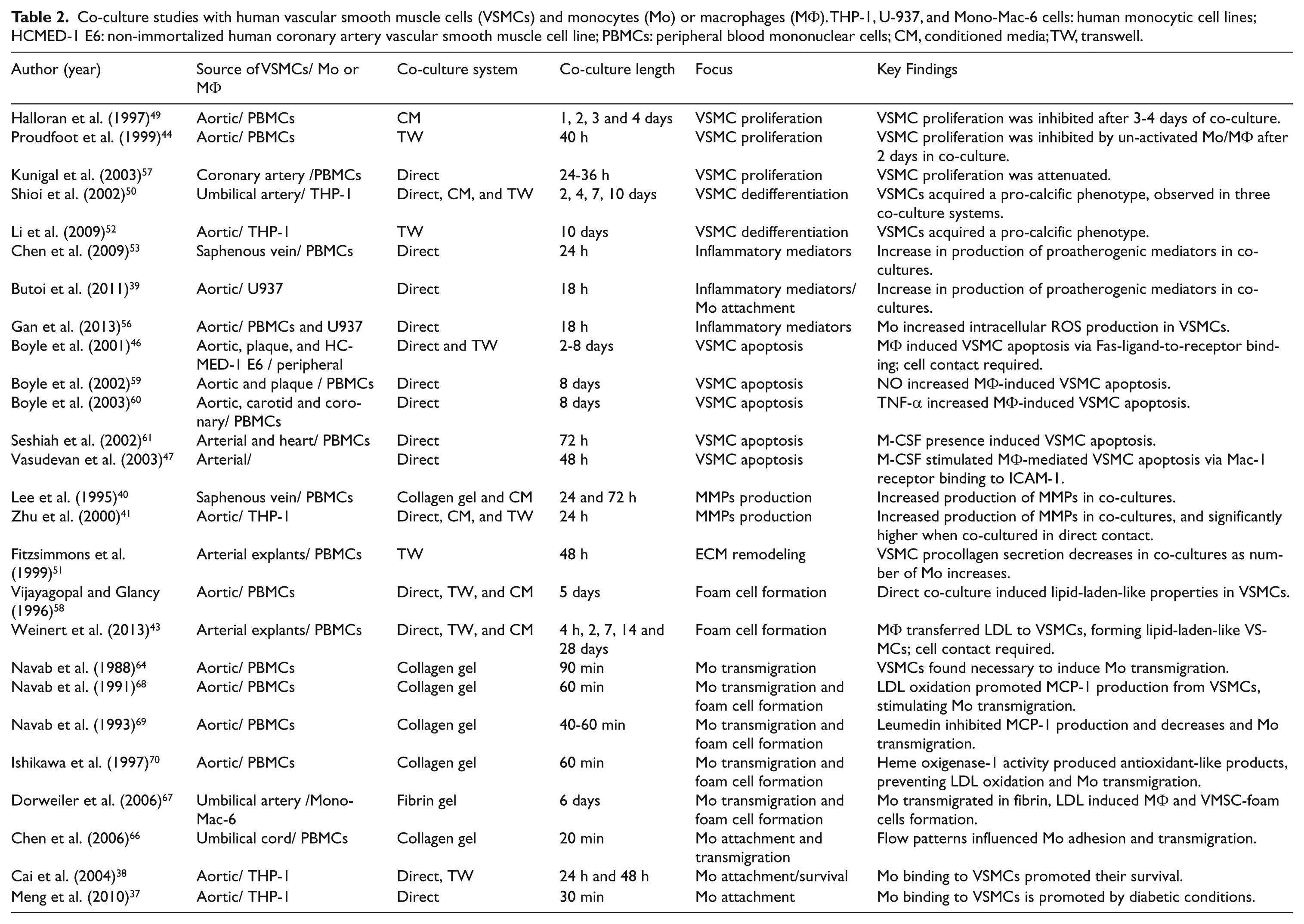
To address the complex interactions between VSMCs and Mo/MΦ, we have classified three main types of in vitro co-culture models used to study various atherosclerotic-related events: (1) indirect cell contact-conditioned media or transwell (Figure 1), (2) direct cell contact (Figure 2), and (3) gel scaffold co-cultures (Figure 3). All these models have been widely utilized. This review will primarily focus on studies performed with human VSMCs and Mo/MΦ (Table 2).
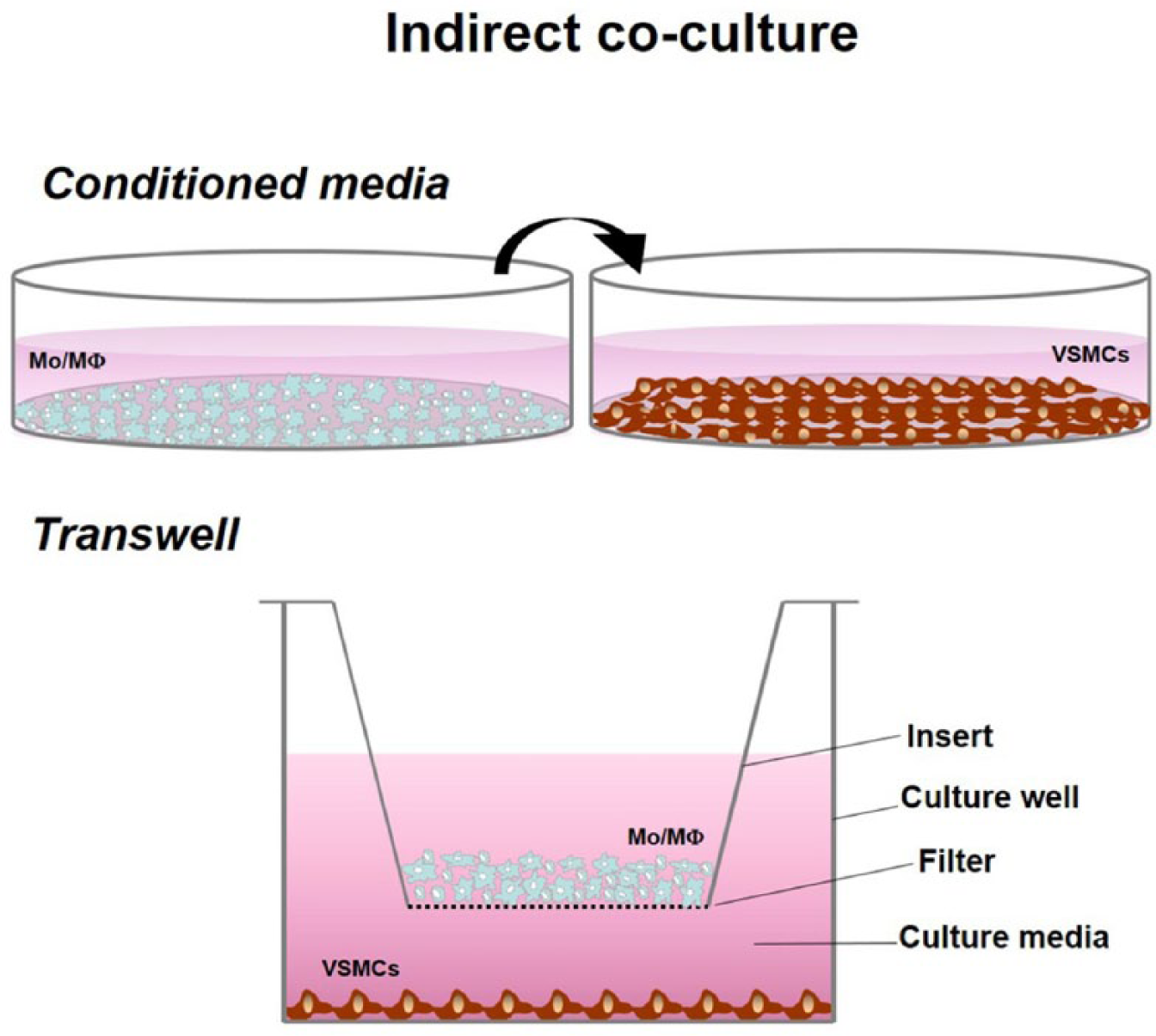
Indirect co-culture model. Transwell – VSMCs are seeded in the culture well and Mo/ MΦ on top of the insert. Conditioned Media – each cell type is cultured separately and the supernatant of one culture is used to treat the other. Cell-to-cell exchanges relay in soluble secretions.
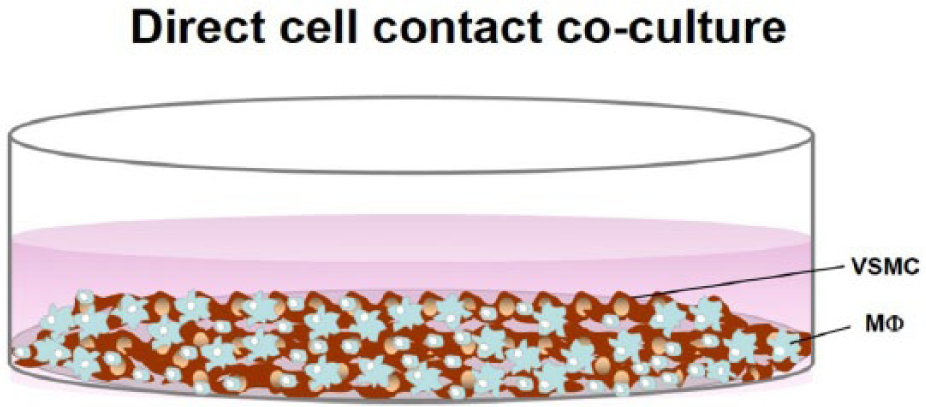
Direct co-culture model. VSMCs and Mo/ MΦ are cultured together for cell-to-cell interaction.
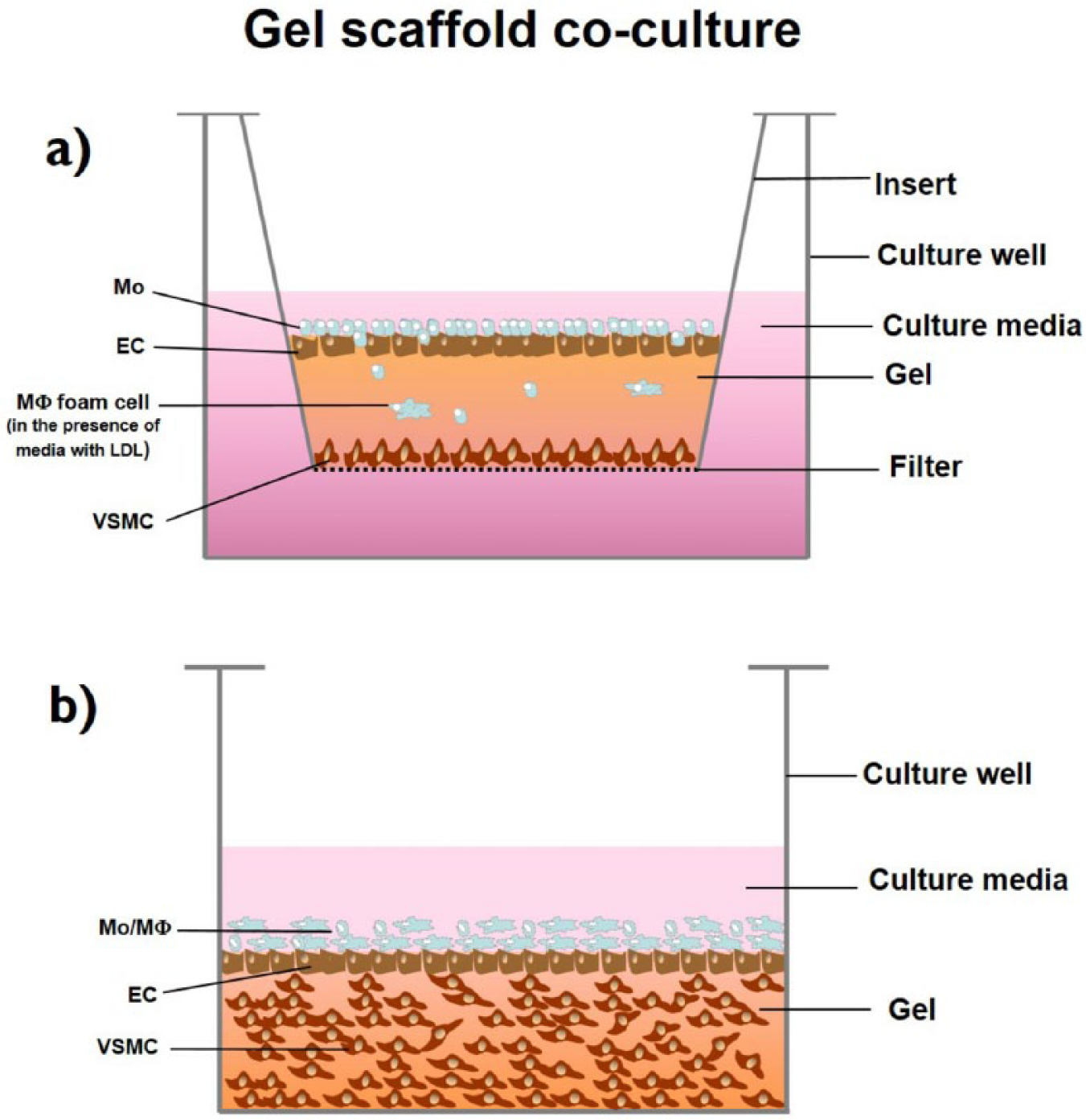
Gel scaffold co-culture model. A gel made of collagen or fibrin is used as an artificial intima. VSMCs are cultured below the gel (gel-separation model) (a) or they are anchored in the gel and (gel-anchoring model) (b); Mo/ MΦ rest on top of gel, optionally separated from VSMCs with a layer of endothelial cells [Illustration partially based on co-culture descriptions and depictions from the reviewed articles: Navab et al., 64 Chen et al., 66 and Dorweiler et al. 67 ].
Co-culture studies with human vascular smooth muscle cells (VSMCs) and monocytes (Mo) or macrophages (MΦ). THP-1, U-937, and Mono-Mac-6 cells: human monocytic cell lines; HCMED-1 E6: non-immortalized human coronary artery vascular smooth muscle cell line; PBMCs: peripheral blood mononuclear cells; CM, conditioned media; TW, transwell.
Indirect co-culture
Conditioned media (CM) and transwell co-cultures are often jointly applied to investigate VSMC and Mo/MΦ interactions. These co-culture methods utilize the secretory properties of Mo/MΦ to study their cross-talk with VSMCs (Figure 1). Researchers have employed similar methodology in transwell and CM co-cultures to address phenotype change, proliferation, and cytokine production.
Conditioned media
CM from Mo/MΦ may be collected pre- or post-activation and used to test its effects on VSMCs. This method allows for studying the secretory cytokines from Mo/MΦ, as well as experimental flexibility since CM can be frozen and stored for later use.45,49 Since media-suspended cells or residual culture-debris may affect the outcomes of some assays, CM can also be centrifuged or filtered prior to usage.42,50
The CM method has been used to study the effects of Mo/MΦ on VSMC proliferation, dedifferentiation and MMPs production. Halloran et al. generated CM from MΦ to test its effects on VSMC proliferation. 49 Mo were cultured in suspension to produce MΦ, and their supernatants (CM) were collected post-activation with LPS. The MΦ-CM was diluted with normal growth media and evaluated for its effects on VSMC proliferation. The diluted MΦ-CM was found to inhibit VMSC proliferation after 48 hours in co-culture. Tintut et al. investigated the roles of MΦ-CM, derived from activated THP-1 cells (a monocytic cell line) or monocytes, respectively, on vascular calcification.42,50 CM co-cultures maintained for three or four days induced VSMC dedifferentiation towards a pro-calcific phenotype, which was determined by measuring the activity of the isoenzyme alkaline phosphatase (ALP), an osteoblast phenotypic marker. Focusing on the factors responsible for VSMC dedifferentiation, Tintut et al. identified that an increased immunoreactivity for tumor necrosis factor alpha (TNF-α) in CM from LPS-activated MΦ was associated with increased ALP activity in VSMCs. Other studies have focused on the impact of Mo-CM on MMPs production from VSMCs. Using a 24-hour CM treatment based on a 1:1 cell ratio of VSMCs to Mo, Lee et al. found untraceable amounts of MMP-1 and MMP-3 in individual VSMC or Mo cultures, but both MMPs were markedly present in cultures of VSMCs treated with Mo-CM. 40 Zhu et al. utilized a similar 1:1 VSMC to Mo co-culture approach with THP-1 cells to generate CM and evaluate MMP-1 for 48 hours. 41 The study corroborated the lack of MMP-1 production by Mo and only small amounts by VSMCs. However, after co-culture the production of MMP-1 by VSMCs increased 50 fold, as detected by the enzyme-linked immunosorbent assay (ELISA), suggesting that Mo secretions present in the CM can induce VSMC production of MMPs.
Transwell
Unlike CM, both cell types are present in transwell co-cultures, but a membrane filter and distance prevents direct cell-to-cell contact. One cell type is cultured on the bottom of culture plate and the other on the insert filter; each cell type can be maintained separately until ready for co-culture. Co-culture is achieved by placing the insert into the culture plate, which rests at a fixed distance from the bottom of the culture well. The variability of membrane filter types and pore size facilitate a range of applications such as drug transport, permeability studies, or cell-cell/cell-substrate interactions within a co-culture system. In addition, this model allows microscopy examination before and after co-culture, as well as cell collection.
In transwell co-cultures, VSMCs are typically seeded in the culture plate and MΦ on top of the filter, allowing VSMCs to grow a desired density while Mo are maintained and differentiated to MΦ. Using this arrangement, the effects of MΦ on VSMC proliferation, matrix degradation and dedifferentiation have been studied. Proudfoot et al. observed an inhibition of VSMC proliferation in a transwell co-culture system that was maintained for approximately 40 hours with a 1:50 VSMC:Mo/MΦ ratio. 44 Proliferation was determined via radioactive labeling of VSMCs in the bottom chamber of the transwell with 3[H]-Thymidine. A similar approach has also been used to investigate VSMC matrix degradation. Fitzsimmons et al. co-cultured monolayers of 3[H]-proline-labeled VSMCs with 0.5–3 × 106 Mo on inserts to evaluate Mo effects on VSMC procollagen secretion. 51 After a 48-hour co-culture, inhibition of procollagen secretion from VSMCs was observed by evaluating 3[H]-proline incorporation in supernatants. Procollagen is the precursor of collagen, a key VSMC synthetic product highly present in atherosclerotic fibrous caps; thus, these results suggest potential suppressive effects of Mo on VSMC synthetic activity. Lastly, to assess Mo/ MΦ induction on VSMC dedifferentiation towards a pro-calcific phenotype, Shioi et al. cultured a monolayer of VSMCs with 2 × 106 MΦ-differentiated THP-1 cells on transwell inserts, and found that 2–4 days of co-culture induced dedifferentiation, whereas Li et al. tested dedifferentiation of VSMCs in a 10-day co-culture of THP-1/VSMCs.50,52
CM and transwell co-cultures have frequently been used in combination or interchangeably to investigate the effects of soluble secretions from Mo/MΦ on VSMCs. Applying these indirect co-cultures techniques, researchers have demonstrated that Mo/MΦ can affect VSMC proliferation, phenotypic changes, and migrative capacity. However, a key limitation of the current literature is the variation of Mo/MΦ ratios used to produce the documented effects on VSMCs. In some cases, researchers have failed to address the Mo/MΦ to VSMC ratio used, which can influence the levels of cytokine production, as demonstrated by Chen et al. who observed that the production of IL-6 and monocyte chemoattractant protein-1 (MCP-1) in VSMC/Mo co-cultures was proportional to the number of Mo. 53 Yet, these investigations also display the flexibility offered by indirect co-cultures: standard molecular techniques can be used to evaluate protein and gene expression in each cell type, secretory cytokines in supernatants can be examined, and there is potential for microscopy-based analysis in each cell type. The major disadvantage of indirect co-cultures rests on their restriction to soluble secretions; consequently, their utility in mechanisms requiring direct cell-to-cell contact is limited. Multiple studies have documented that some atherosclerosis-related events require cell-to-cell interactions;46,54,55 therefore, alternative approaches may be necessary for a thorough evaluation.
Direct contact co-culture
Direct cell contact co-cultures provide crucial information concerning cell-to-cell exchanges that require cellular junctions to communicate. Usually, VSMCs are allowed to attach at a particular seeding density or confluence on a plate, followed by the addition of Mo/MΦ. Depending on the seeding densities, cells may construct a bilayer or be spread throughout the plate (Figure 2); however, the co-culture ratios and duration may vary depending on the research focus. This co-culture approach has been used to assess a diverse selection of atherosclerosis-related events, such as cytokine secretion, VSMC dedifferentiation, and apoptosis.
Direct co-cultures have been particularly important to investigate how Mo/VSMC interaction affects the production of proatherogenic mediators. Chen et al. studied the production MCP-1 and interleukin-6 (IL-6), two pro-inflammatory mediators with roles on monocyte transmigration and diverse cellular functions. 53 VSMCs were allowed to attach for 24 hours, with subsequent co-cultures evaluated at VSMC:Mo ratios ranging from 1:0.2–30, with or without LPS. After 24 hours in co-culture and analysis of supernatants with ELISA, the authors found a synergistic and significant increase in the production of MCP-1 and IL-6 in co-cultures with LPS-stimulated Mo as compared with monocultures. The effect was detectable in co-culture supernatants at the lowest ratio of 1:0.2 VSMC:Mo, increasing with the number of Mo. Similarly, Butoi et al. and Gan et al. used the human monocyte-like cell line U937 to study the effects of Mo/VSMC cross-talk on the production and expression of different proatherogenic mediators.39,56 Both studies cultured a monolayer of VSMCs and added 1 × 106 of LPS-activated Mo for 18 hours of co-culture. Using ELISA, Butoi et al. first identified increased amounts of TNF-α and IL-6 in co-culture supernatants. Further protein and gene expression analysis in cells after co-culture revealed up-regulation of TNF-α, IL-6, IL-1β, MMP-2, and MMP-9 in both cell types, as compared with monocultures. 39 On the other hand, Gan et al. evaluated the production of resistin and reactive oxygen species (ROS), and determined that Mo/VSMC interaction induced both resistin production from Mo and ROS production from VSMCs. 56
Direct cell contact co-cultures have also demonstrated that Mo-derived MΦ can alter VSMC proliferation and phenotype. Kunigal et al. investigated Mo effects on VSMC proliferation, using immunofluorescence, and the molecular regulatory mechanisms at a 1:3 VSMC:Mo ratio and observed that Mo repressed VSMC proliferation. 57 Vijayagopal et al. co-cultured VSMCs with 2–3 × 106 lipoprotein-loaded MΦ for 5 days and observed that VSMCs were transformed into lipid-laden cells via cell-to-cell lipid transfer from MΦ to VSMCs. 58 Weinert et al. co-cultured fluorescently labeled low-density lipoprotein (LDL) MΦ at a 1:3 VSMC to MΦ ratio for 4 hours up to 28 days, 43 and observed LDL transfer from MΦ to VSMCs and induction of phagocytotic properties in VSMCs. Production of intracellular cholesterol ester (a product of intracellular lipoprotein oxidation) in VSMCs was also documented. These observations by the aforementioned investigations suggest that the plastic nature of VSMCs potentially allows them to become an additional source of atherosclerotic foam cells.
Finally, research groups have utilized direct co-cultures to study the effects of Mo/MΦ on VSMC apoptosis. The Bennett research group demonstrated the capability of Mo and un-activated MΦ to induce apoptosis in VSMCs. 46 First, Mo effects were evaluated by seeding VSMCs and adding Mo for a total of 8 days at VSMC:Mo ratios ranging from 1:0.5 to 1:64. VSMC death was observed at a ratio as low as 1:1, increasing with the number of Mo present. 46 Mo were cultured for 6 days to differentiate them to MΦ; MΦ were then directly co-cultured with VSMCs at 1:4 and 1:8 (VSMC/MΦ cell ratio) for an additional 2 or 8 days in co-culture. The authors noted that while cell ratio affected MΦ-mediated apoptosis, the length of co-culture did not impact the results. In the 1:4 co-cultures, 50% of VSMC death was observed, whereas the 1:8 co-cultures induced over 90% of VSMC death. 46 Based on these results, the group then investigated the regulatory mechanisms for MΦ-mediated VSMC apoptosis, and determined that cell-to-cell contact was required to induce apoptosis via Fas-ligand binding and that TNF-α and nitric oxide, two proatherogenic mediators, augmented MΦ-mediated VSMC apoptosis.46,59,60 Using similar techniques, the Goldschmidt-Clermont group evaluated Mo-mediated VSMC apoptosis in 1:3 VSMC to Mo co-cultures in the presence or absence of M-CSF;47,61 however, Mo-mediated apoptosis was only observed when M-CSF was present, and in a concentration-dependent manner. These outcomes accentuate the complexity of atherosclerotic cardiovascular disease associated with multiple atherogenic mediators and growth factors, which can affect molecular exchanges and cellular behavior.
Direct cell contact co-cultures have provided insightful evidence for VSMC and Mo/MΦ exchanges, addressing the impact on VSMC proliferation, apoptosis, lipid-uptake, and production of pro-atherogenic mediators. The major advantage of direct cell-contact co-cultures appears to be a more accurate representation of in vivo exchanges between VSMCs and MΦ/Mo, including both juxtacrine and paracrine signaling, which cannot be accomplished with indirect co-cultures. Yet, the key limitation of this model lies in the lack of techniques to separate cells for further analysis without modifying their properties. Similar to indirect co-culture models, a major constraint of the existing literature is the lack of consistency in cell species (not addressed in this review), cell ratio, and co-culture duration for this model. Nonetheless, it appears that ratios of 1:0.2 to 1:8 VSMCs to Mo/MΦ are commonly used, and these ratios are adequate to produce the targeted effects.
Gel scaffold co-cultures
Gel scaffold platforms originated from tissue engineering research groups as a potential means to study cells in a three-dimensional (3D) environment. Because cellular exchanges in living organisms occur in 3D, gel scaffold platforms were created to more closely mimic an in vivo setting. Collagen and fibrin are the two main substrates utilized for the creation of scaffolds that simulate an extracellular matrix environment. 62 Type I collagen is the most common type used for collagen scaffolds while fibrin scaffolds are constructed from fibrinogen and thrombin. 63 These substrates create a fiber network for natural cell and protein anchorage. Collagen and fibrin polymerize at 37°C and neutral pH, generating a loose network of fibers upon polymerization.64–67 These properties allow successful cell culture and anchorage, and free passage of molecules. For atherosclerosis-related studies, gel-based co-cultures have been arranged in two different ways: gel-separation and gel-anchoring models (Figure 3). Both set-ups are primarily employed to investigate Mo infiltration and MΦ transformation in the presence of VSMCs.
In the gel-separation model (Figure 3(a)), a polycarbonate filter is placed inside a culture well and VSMCs are cultured above the filter to confluence. Then, a layer of collagen gel is coated over the VSMCs and Mo/MΦ are added on top of the polymerized gel; a layer of ECs may also be grown over the gel prior to Mo/MΦ incorporation. The filter serves as support for the multilayered structure and it also permits free media passage. Using this design, the Fogelman research group created the first successful in vitro, gel-based co-culture model for atherosclerosis applications. 64 VSMCs were cultured to confluence, followed by a layer of ECs, and then a Mo layer of 2.5–5 × 105 cells/cm2 added in the presence of a chemotactic agent. Maintaining the co-cultures for 40–90 minutes, the Fogelman research group studied the mechanisms behind LDL-mediated Mo transmigration, transmigration inhibition, and MΦ lipid-laden formation.64,68–70 First, the Fogelman research group determined that the three components in the co-culture (collagen, VSMCs, and ECs) were necessary to induce Mo transmigration. Collagen by itself and VSMCs or ECs combined with collagen had little effect on Mo transmigration; conversely, when the three components were present, over 11% of Mo were induced to migrate. 64 The research group then focused their subsequent investigations on the roles of LDL for Mo transmigration. Navab et al. observed that Mo transmigration increased over seven folds in co-cultures with LDL as compared those without LDL, inducing foam cell formation. Searching for the mechanisms involved, the group determined that LDL oxidation (ox-LDL), promoted the production of MCP-1 by VSMCs and ECs in co-cultures, enhancing Mo attachment and their subsequent transmigration. 68 Further investigations also identified mechanisms to inhibit LDL-mediated Mo transmigration through the prevention ox-LDL, and thereby inhibition of MCP-1 production,68,69 and via induction of antioxidant-like products in co-cultures. 70
In the gel-anchoring model (Figure 3(b)), VSMCs are suspended in the collagen or fibrin solution, plated in culture wells, and the substrate is polymerized at 37°C to induce anchorage of VSMCs. The VSMCs are grown for several days/weeks within the gel, and eventually Mo/MΦ are added on top of the gel for co-culture. ECs may also be included in the system by seeding them on top of the gel-embedded VSMCs prior to the inclusion of Mo/MΦ. Chen et al. assembled a collagen gel co-culture by suspending 1 × 106 VSMCs/ml in a collagen solution. 66 A layer of ECs was cultured on top of the gel, and a flow chamber was incorporated to deliver Mo at 2 × 105 cells/ml in order to study the effects of different flow patterns on Mo attachment and transmigration. The co-cultures were maintained for 20 minutes and observed with video-microscopy, which showed increased adhesion and transmigration when Mo were delivered in disturbed flow patterns. Because atherosclerotic plaques tend to form in vessel areas with disturbed flow, this co-culture model serves as a promising approach to investigate the roles of flow disturbance in atherosclerosis. Similarly, Dorweiler et al. used 6 × 105 VSMCs/ml to create the first fibrin-gel model. 67 The embedded cells were grown for 14 days to produce a multilayered structure of VSMCs. A layer of ECs was cultured on top of the fibrin-embedded cells for an additional 14 days before Mo were added at 1 × 106 cell/ml, and the co-culture was maintained for 6 more days. The co-culture was fixed and Mo infiltration/foam cell formation was studied with immunostaining and electron microscopy. The authors observed trans-endothelial migration of Mo and Mo differentiation to MΦ within 4 days. Incubation with LDL resulted in foam cell formation not only for MΦ, but also VSMCs, identifying an additional source of foam cell relevant to atherosclerotic progression.
The primary advantage of the gel-based co-cultures is the ability to integrate and study multiple cell types in 3D. These models can be fixed after co-culture and processed for visualization histologically and microscopically.66,67 While 3D cultures are relatively new in the cardiovascular field, they may play important roles in atherosclerotic research in the future. Several studies have suggested significant differences between two-dimensional (2D) and 3D cell culture. Li et al. performed genomic analysis comparing 2D and 3D collagen-based cultures of VSMCs, and determined that VSMCs cultured in 3D exhibited less proliferation and about one-third less α-actin expression as compared with 2D. 71 Stegemann and Nerem reported a similar alteration of VSMC function in collagen-gel-embedded culture using flow cytometry, 72 and confirmed that the matrix substrate of 3D gel-based cultures may interfere with assay outcomes.
Studies with non-human cells
Thus far, we have focused on VSMC and Mo/ MΦ co-culture using human cells, but in vitro co-cultures with cells from other animal species, including mouse, rat, and rabbit, have been extensively utilized to investigate Mo/ MΦ and VSMCs interaction. These studies have shown an alteration of VSMC proliferative activity when co-cultured with MΦ,45,72–74 an up-regulation of proatherogenic mediators, 75 and MΦ-mediated vascular dedifferentiation towards a pro-calcific phenotype.52,76,77 However, experimental findings from these investigations, particularly those addressing VSMC proliferation, have not always correlated with studies involving human cells. For instance, Greenburg et al. showed that MΦ induced an increase in VSMC proliferation using rabbit and rat VSMC/MΦ co-culture systems, respectively,73,74 while studies with human cells (Table 2) have found opposing results, suggesting that an extended co-culture with un-activated and pro-inflammatory activated Mo/MΦ decreases VSMC proliferation.
In addition, numerous murine models have been developed to investigate MΦ involvement in vascular disease formation by studying MΦ distribution, phenotype, and plasticity in atherosclerosis.45,78 However, as previously mentioned, only about 50% of MΦ polarization markers overlap between humans and mice at the transcriptional level, 2 and no comparisons between human, rat, and rabbit MΦ have been noted. Moreover, a recent investigation comparing gene expression and signaling pathways between human subjects with different inflammatory stresses and their counterpart murine models, found a poor correlation between the two species’ response to inflammation. 79 Seok et al. also highlighted the wide differences in immune system adaptations between humans and mice. Such discrepancies emphasize the limitations in utilizing in vivo animal models to study human macrophages and highlight the importance of using test models that mimic the human condition at the molecular level. Therefore, employing in vitro co-culture models with human cells for atherosclerotic cardiovascular disease investigations can help to ensure relevance and reliability, not only for mechanistic studies, but also for drug discovery and therapeutic applications.
Relationship between human atherosclerosis and in vitro co-culture
Inflammation plays a key role in VSMC dysfunction and atherosclerotic plaque formation, 80 and an examination of clinical plaques has revealed close relationships between VSMCs and MΦ that can account for atherosclerotic inflammation.81,82 Increased MΦ content has been linked to acute coronary syndromes and unstable atherosclerotic plaques. 16 In addition, dispersal of MΦ subtypes within plaques has been correlated to the distribution of several inflammatory mediators within associated plaque regions.83–86 Stöger et al. observed that both M1 and M2 macrophages can both increase with atherosclerotic plaque progression. 87 The fibrous cap regions of atherosclerotic plaques appear to have equal amounts of the two general MΦ subtypes, but more distinct patterns emerge when comparing vulnerable versus stable areas. Rupture-prone shoulder areas contain more M1 macrophages, whereas M2 are more common in the adventitia and cell-rich areas.87,88 Finally, MΦ are known to induce VSMC apoptosis, 9 which has been associated with an increased macrophage accumulation and plaque vulnerability.14,89
In addition to cell types, an examination of human atherosclerotic plaques has provided general insights about VSMC:Mo/MΦ ratios. In a comparison of intact versus thrombotic atherosclerotic plaques, Davies et al. measured the plaque area occupied by VSMCs and Mo/MΦ and observed that only 10–25% of the area was composed of these two cell types, while the majority of the plaque composition was the ECM. 14 Intact plaque caps were composed of 11.8% VSMCs and 2.9% MΦ whereas thrombotic plaque regions contained 2.8% VSMCs and 13.8% MΦ. This represents a significant shift in cell composition with a VSMC:MΦ ratio of approximately 4:1 for intact and 1:5 for thrombotic plaque areas. Similarly, van der Wal et al. examined thrombosed atherosclerotic plaques, and reported that the ruptured areas are almost completely absent of VMSCs and primarily populated by MΦ. 15 Evaluating patients affected with angina, Moreno et al. reported that the plaque area occupied by MΦ from patients affected with unstable angina and myocardial infarction is larger (~ 8–20%) than those with stable angina (~2–4%). 16 Bauriedel et al. showed that patients with stable angina contained a VSMC:MΦ ratio of approximately 9:1, while those from patients with unstable angina had a 3:1 ratio. 90 Based on these studies, VSMC:MΦ ratios of 1:5 or more may represent the most vulnerable/thrombotic stages of atherosclerotic plaque development, while ratios of 4:1–3:1 could account for advanced, but not necessarily thrombotic unstable plaques.
Although not all studies discussed in this review specify ratios, those that do suggest that decreased ratios of VSMCs to Mo/MΦ and higher Mo/MΦ content (1:1, 1:4, and 1:8) lead to functional changes in VSMC physiology, such as in proliferation, phenotype, and apoptosis. A prime example of such would be the 1:4 ratio used by Boyle et al. to evaluate VSMC apoptosis (inducing 50% of VSMC apoptosis over a 2–8 day co-culture), which is close to the 1:5 ratio found in thrombosed plaques.14,46 The overall effect of Mo/MΦ on VSMC phenotype and function in co-culture models correlate with the physiological changes observed in atherosclerotic VSMCs, suggesting the relevancy of these investigative models in evaluating therapeutic targets in human atherosclerosis.
Limitations of in vitro co-culture
Different co-culture systems and their associated advantages have been discussed; however, the proper selection of the most appropriate system requires critical evaluation of the phenomena under investigation. Table 2 summarizes the studies discussed in this review and a trend of mechanism-based selection can be observed. Overall, indirect co-culture systems can address questions regarding soluble mediators, and direct co-culture systems are more appropriate for molecular mechanism(s) that requires direct cell–cell interactions. A deficit in the current literature is the inconsistent use of polarized/stimulated MΦ/Mo; whereas some investigations report polarization towards a particular MΦ phenotype, others use unstimulated MΦ or fail to address the phenotypic state of the cells. Nonetheless, outcomes generally agree within each system (indirect or direct), but results may vary between indirect co-culture and direct co-culture systems (Table 2; Key findings). When similar results were observed in both systems individually, a more amplified effect was typically seen from direct co-culture.41,42 On the other hand, gel scaffold systems allow the opportunity to engineer a ‘plaque structure’ using particular cell ratios in an ECM, but this system is largely used to examine the mechanisms of Mo/MΦ, such as Mo/MΦ attachment, infiltration and transformation. The effectiveness of this system in evaluating the effects of MΦ on VSMCs remains uncertain, and requires an improvement in technique development for molecular-specific studies.
Several other factors may also limit the widespread use of human cell co-culture systems. Management of primary cells can be challenging. In particular, identifying a suitable patient cohort, and developing a process for cell acquisition, Mo isolation and differentiation are costly and time-consuming tasks that require meticulous planning.91,92 Establishment and replication of complex co-culture systems can be an additional obstacle. For example, the gel scaffold systems require an appropriate substrate selection and preparation in addition to obtaining and arranging the multiple cellular components. The selection of proper co-culture conditions to ensure optimal growth conditions for both cell types under study may further bias investigators towards a simpler monoculture option. This review systemically provides a comprehensive overview of cell-cell ratio, conditions, and experimental design in various co-culture systems.
Another import aspect to consider is substrate material for cell growth. Collagen is the most abundant component of the basement membrane and ECM in healthy tissues, 93 and fibrinogen and thrombin are two essential proteins for coagulation. 63 Whereas culture plates and transwell inserts offer easy culture management, these surfaces poorly resemble the natural ECM conditions. Aiming for better resemblance, some researchers opt for pre-coating of plates or filter inserts with collagen. Transwell inserts are most commonly made of either polycarbonate or polystyrene, and the most common substrate for tissue culture plates is polystyrene. Although polycarbonate and polystyrene are similar in biocompatibility properties for tissue culture goals, 94 transwell inserts and culture plates differ in stiffness and topography, which can influence cell behavior.95–98 Transwell inserts are porous, thin membranes with some tensile properties, while polystyrene plates are flat and rigid. In the context of atherosclerosis, findings suggest that VSMC proliferation and migration increase with surface stiffness,99–101 while the surface topography can influence MΦ spreading, size, and activation.102,103 Gel stiffness in gel scaffold-based systems also impacts cellular behavior and VSMC physiology,104,105 but a recent study suggested that stiffness-mediated response in gels vanishes when VSMCs form cell-to-cell contact instead of being sparsely scattered. 106 Thus, substrate controls may be necessary to discern experimental outcomes.
Compositional variations of collagen types and fibrin in the ECM and basement membrane can also impact the experimental outcomes. Stable atherosclerotic plaques are characterized by their high collagen content, while vulnerable plaques have thin fibrous caps due to limited fibrillar interstitial collagen. 107 In addition, fibrin has been reported to be absent in healthy vascular walls, but it is present in fibrous and advanced atherosclerotic plaques, particularly in thrombotic areas and those with loose connective tissue. 108 Consequently, while fibrin and collagen both represent good scaffold sources, the composition of collagen and fibrin may need to be altered based on the health state of the ECM, and a purpose-guided selection is encouraged.
Clinical implications and future directions
Although examination of clinical samples has provided important information on cell–cell interaction, the uncertainty in the underlying molecular mechanisms restrains clinical therapeutic strategies for disease treatment and prevention. Most of the knowledge on the effects of macrophages in VSMCs is based on in vitro studies overlooking potential cellular cross-talk and the multiple cytokines involved in the pathology of atherosclerosis. With the discovery of complex MΦ subtypes and the plasticity of MΦ and VSMCs, the exchanges between both cell types need to be considered to better understand atherosclerotic plaque formation and progression. The VSMC-MΦ co-culture systems offer a unique opportunity for more in-depth investigations.
A recent review highlighted the prospective of atherosclerosis prevention from the anti-inflammation standpoint. 109 Because of their plasticity properties and the blood-borne origin of some MΦ, these represent a possible therapeutic target to reduce atherosclerotic inflammation in cardiovascular disease. Antagonists against MΦ-produced inflammatory cytokines/factors can be used to suppress some MΦ function. Therefore, understanding the interactions between Mo/MΦ and the surrounding vascular cells becomes increasingly important. In vitro co-culture systems can be a valuable tool to further explore the signal transduction pathways and cytokines involved in Mo/MΦ and VSMC exchanges at the early stages of human atherosclerotic disease for disease prevention. Future studies are needed to elucidate how interactions between VSMCs and Mo/MΦ affect atherosclerotic plaque formation in the presence of appropriate substrates. In addition, there is limited data on how VSMCs and proatherogenic mediators affect MΦ physiology, proliferation, and death. The mechanisms and biomarkers of MΦ polarization in atherosclerosis and mechanisms of VSMC phenotype regulation also need to be clarified. Advanced technologies in vessel wall mimicking, such as microfabrication and microfluidic systems, have been created for the study of angiogenesis. 110 These technologies may offer potential applications for atherosclerosis investigations in the future after significant improvements in material constructs and technique standardization.
Conclusion
Indirect and direct cell contact co-culture systems have been used to investigate the regulatory effects of Mo/MΦ on VSMC functionality, whereas gel scaffold co-cultures have been mainly utilized to evaluate Mo infiltration and MΦ transformation. These are key aspects involved in the pathogenesis of atherosclerosis. The co-culture systems described in this review highlight the impact of both cell types in disease progression, and the necessity of considering cell-to-cell interactions. In this review, we examined cell types and ratios, culture conditions, and targeted effects in various human Mo/MΦ and VSMC co-culture models. We also described the plastic nature of both cell types and summarized and correlated clinical observations with cellular processes. While this review focused on the regulatory effects of Mo/MΦ on VSMC function, it is important to recognize that modulation occurs both ways. As the scientific community ventures to fully understand the pathogenesis of atherosclerosis and to identify novel therapeutic targets for atherosclerotic disease treatment and prevention, the importance and utilization of an in vitro co-culture model with human cells should be recognized for the contributed value added to research studies.
Footnotes
Declaration of conflicting interest
The author declares that there is no conflict of interest.
Funding
This work was supported by the U.S. Department of Veterans Affairs [grant number I01BX001398] and the National Institutes of Health [grant number NHLBI 1T32HL098049].